Проблемы интродукции и внутривидовой гибридизации Apis mellifera

Автор: Удалов М.Б., Козьминов С.Г., Беньковская Г.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 1-3 т.12, 2010 года.
Бесплатный доступ
В работе приведены экспериментальные данные по изучению проблемы внутривидовой гибридизации медоносной пчелы Apis mellifera на Южном Урале. Для оценки состояния аборигенных, интродуцированных и гибридных семей медоносных пчёл использовались методы полимеразной цепной реакции, а также биохимические параметры стресс-реакции рабочих пчёл.
Медоносная пчела, интродукция, внутривидовая гибридизация
Короткий адрес: https://sciup.org/148199052
IDR: 148199052 | УДК: 595.7:575.174.015.3
Problems of introduction and intraspecific hybridization of Apis mellifera
In work experimental data on studying the problem of intraspecific hybridization of melliferous bee Apis mellifera in Sonth Ural are resulted. For an estimation of condition native, introductive and hybrid families of melliferous bees methods of polymerase chain reaction, and also biochemical parameters of working bees stress-reaction were used.
Текст научной статьи Проблемы интродукции и внутривидовой гибридизации Apis mellifera
Ареал башкирской популяции среднерусской расы медоносной пчелы Apis mellifera mellifera сокращается, и основную часть ареала вида в республике составляет расширяющаяся гибридная зона [3]. Этим процессам активно способствуют изменения экологических условий, связанные с антропогенными воздействиями, а также бесконтрольный завоз на пасеки Республики семей, принадлежащих к южным расам ( A.m. caucasica, A.m. carpatica и другим), отличающимся большей продуктивностью и длительным периодом сбора нектара и пыльцы. Гибридные особи и семьи в большинстве своем хуже переносят длительную зимовку, подвержены заболеваниям. В республике резко снижается продуктивность семей и отмечается большая гибель, как следствие сплошной гибридизации [4].
Полиморфизм длин рестрикционных фрагментов изучался и для ядерной [6], и для митохондриальной ДНК (мтДНК) пчелы [7]. Затем использование метода полимеразной цепной реакции (ПЦР) позволило выявить различия в структуре межгенного участка мтДНК COI-COII медоносной пчелы, что было использовано нами для оценки генетического полиморфизма в семьях Apis mellifera на территории Башкортостана [2]. Локус COI-COII у расы A.m. mellifera с частотой встречаемости более 0,99 представлен аллелем PQQ. У южных рас A.m. caucasica, A.m. carpatica и др. (основной источник генетического загрязнения башкирской популяции A.m. mellifera ) аллель Q, возможно, фиксирован. Ожидаемый размер ам-плифицируемых с использованием специфических праймеров фрагментов мтДНК составлял 350 пн в случае комбинации Q и 600 пн при комбинации PQQ [2]. Как и ожидалось, нами были выявлены различия в размерах амплифицированных
фрагментов мтДНК у A.m. mellifera и A.m. caucasica . Они составили 600 и 350 пн, соответственно (рис. 1).
Использование метода ПЦР дало возможность более детально исследовать вариабельный участок мтДНК пчелы [5]. Даже при сравнительно небольшой выборке, описанный выше метод позволяет охарактеризовать каждую семью по вариабельности локуса COI-COII и может быть предложен для анализа расового происхождения по материнской линии A. mellifera . В Иглинском районе на племенных пасеках были выявлены семьи, заявленные как чистопородные с принадлежностью к A.m. mellifera , и, тем не менее, содержащие мтДНК A.m. caucasica . В дальнейшем проверка расовой принадлежности семей медоносной пчелы по мтДНК использовалась как обязательный компонент во всех экспериментах.
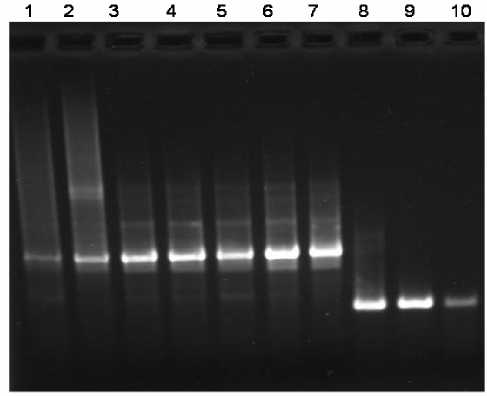
Рис. 1. Результаты электрофоретического разделения продуктов, полученных при амплификации локуса COI- COII мтДНК Apis mellifera L.: Образцы 1-7 – A.m. mellifera (600 п.н.), образцы 8-10 – A.m. caucasica (350 п.н.), образцы 1-5 получены из семей в заповеднике Шульган-Таш (среднерусская раса); 6-8 – с пасек в Иглинском районе; 9 и 10 – с пасеки учхоза БГАУ (кавказская раса)
Интродукция видов и подвидовых группировок насекомых в новых регионах влечет за собой первоначальный период адаптации к условиям соответствующих эколого-географических зон. Семьи пчел A.m. caucasica, интродуцируемые на территории Башкортостана в течение относительно короткого периода, менее адаптированы к условиям Южного Урала, чем аборигенные семьи A.m. mellifera, что показали результаты наших экспериментов.
Нами были исследованы тирозиназная и ДОФА-оксидазная активности у пчел среднерусской ( A. m. mellifera L.) и кавказской ( A. m. caucasica Gorb.) рас в норме и их распределение в организме имаго рабочих особей. Гибридизация в тех случаях, когда биоценотические условия остаются стабильными, может проявиться и в положительных эффектах (гетерозис), но нельзя забывать о том, что при этом высока вероятность разрушения сложившихся комбинаций и, соответственно, снижения адаптивного потенциала, по крайней мере, на начальном этапе процесса гибридизации.
Для сравнения состояния рабочих особей в чистопородных и метизированных семьях по каждому из биохимических показателей (ферменты фенолоксидазной и гидролазной систем) были проведены измерения в выборках семей средне- русской, кавказской (по 50 семей) и карпатской (10 семей) рас, а также в выборке из 50 семей гибридизированных (в ходе пасечного содержания) пчел. Измерения проводились в одних и тех же семьях весной и осенью 2001 и 2002 гг. (табл. 1). В таблице приведены не средние для группы, а максимальные значения параметров, поскольку именно с их помощью удалось выявить существенные различия между группами семей. В зависимости от года и сезона разброс показателей оказался довольно значительным, однако удалось установить существование различий между особями из чистых и гибридных семей практически по всем показателям Таким образом, эти величины могут служить для предварительной оценки чистопородно-сти семей. Установлено воспроизводимое расхождение в значениях по основным группам ферментов, признанных интегральными показателями состояния у насекомых (лизосомальные гидролазы), либо выполняющих конкретные защитные функции (фенолоксидазный комплекс) в семьях пчел среднерусской расы, аборигенных на территории Башкирии и Южного Урала в целом, и в семьях пчел интродуцируемых южных рас (кавказской и карпатской).
Таблица 1. Максимальные значения биохимических параметров (удельная активность в нативных условиях) в семьях пчел из различных групп
|
Ферменты |
Среднерусская раса |
Кавказская раса |
Карпатская раса |
Гибридные семьи |
|
ДОФА-оксидаза |
0,01±0,0008 |
0,01±0,0005 |
0,012±0,001 |
0,011±0,001 |
|
тирозиназа |
0,048±0,003 |
0,013±0,001* |
0,033±0,002* |
0,031±0,003* |
|
щелочная фосфатаза |
0,81±0,077 |
1,21±0,1* |
1,01±0,08* |
1,39±0,09* |
|
кислая фосфатаза |
0,89±0,07 |
1,24±0,012* |
2,38±0,17* |
0,86±0,066 |
|
эстеразы |
7,32±0,56 |
9,42±0,8* |
19,7±1,2* |
6,9±0,45 |
Примечание : * – достоверное (Р≥0.95) отличие от особей среднерусской расы
Существенное отличие, позволяющее дифференцировать семьи кавказской и карпатской рас, отмечено для уровней активности кислой фосфатазы и неспецифических эстераз. У особей карпатской расы эти показатели выше, чем у пчел среднерусской расы почти в 3 раза, тогда как у особей кавказской расы они выше всего в 1,5 раза. Гибридные семьи отличаются от чистопородных по уровню активности щелочной фосфатазы (значения уровня активности выше, чем у пчел среднерусской расы, в 1,7 раз). Следует отметить несовпадение изменений по сравнению с чистопородными семьями. Так, у гибридных особей по сравнению с особями из семей среднерусской расы снижена активность кислой фосфатазы и неспецифических эстераз и повышена активность ДОФА-оксидазы, однако эти отличия статистически мало достоверны. Данный набор биохимических маркеров позволяет дифференцировать не только семьи аборигенной среднерусской расы и южных рас, но и интродуцируемые расы между собой. Возможно также выделение гибридных семей.
О перспективности селекции гибридных семей и семей интродуцентов свидетельствуют достаточно редкие, но наблюдающиеся проявления гетерозиса, что можно продемонстрировать на примере одного из наших экспериментов с бактериальным препаратом (БТБ). Чаще в аналогичных экспериментах наблюдается отрицательное влияние гибридизации, выражающееся в снижении жизнеспособности взрослых особей в гибридных семьях (табл. 2). Реакция на токсикант (БТБ, 0,5%) у пчел разных рас сопровождается значительно различающимися изменениями активности тирозиназы. Максимальные значения для каждой временной точки стресс-реакции отмечены для среднерусских пчел, при этом профиль динамики активности характерен именно для стрессовой реакции. Изменения активности тирозиназы у пчел кавказской и карпатской рас имеют противоположную направленность и приводят к 3х-кратному снижению ее в конечной точке регистрации. Примечательно, что динамика активности тирозиназы у гибридных особей абсолютно несходна ни с одним из описанных паттернов. Судя по всему это также является следствием дисбаланса генетических комплексов у гибридных особей пчел.
Модель стресс-реакции может служить основой для оценки уровня адаптированности к региональным условиям у аборигенных и интродуцируемых семей разных рас. У особей интродуцируемой расы A.m. caucasica изменения локомоторной активности, динамика активности ферментов фенолоксидазного и гидролазного комплексов достоверно отличаются от таковых для особей аборигенных семей A.m. mellifera. Выявленные особенности стресс- реакции свидетельствуют о том, что семьи интродуцируемых южных рас медоносной пчелы переносят стресс, связанный с акклиматизацией в новых эколого-климатических условиях.
Таблица 2. Отрицательные эффекты гибридизации в семьях Apis mellifera (2004 г.).
|
Раса |
мтДНК, COI-COII |
Длительность жизни без воздействия, сутки |
Длительность жизни после воздействия БТБ, сутки |
Коэффициент вариации, % |
|
A.m. mellifera |
PQQ |
16,5 ± 0,5 |
10,0 ± 1,0* |
10,00 |
|
A.m. caucasica |
Q |
16,5 ± 0,5 |
7,5 ± 0,5* |
6,67 |
|
Гибридные семьи |
PQQ/Q |
16,3 ± 0,82 |
7,83 ± 2,14* |
27,33 |
Примечание: * – достоверное (Р≥0,.95) отличие от контрольных вариантов. Приведены средние значения с ошибкой
Классический путь развития защитной реакции у насекомых на воздействие неблагоприятных факторов, к которым относятся и резкие изменения температуры, в гипотетической форме был описан достаточно давно [1], однако с проблемами адаптированности насекомых это не связывалось. Нами было высказано предположение о существовании различий на подвидовом уровне в механизмах реализации защитных реакций у вида Apis mellifera L. Различия между расами медоносной пчелы в контролируемых условиях не слишком велики; но модель стресса, вызванного каким-либо фактором, позволяет сделать эти различия значимыми. Адаптированность пчел башкирской популяции среднерусской расы к региональным условиям и отсутствие такой адаптированности у пчел кавказской расы, относительно недавно интродуцируемой на территории Башкортостана, нашли отражение в последовательности и интенсивности активации компонентов фенолоксидазной ферментной системы в условиях теплового стресса, что особенно наглядно удалось продемонстрировать на фоне влияния аскорбиновой кислоты. Как установлено, рабочие взрослые особи из метизи-рованных семей отличаются в большинстве случаев более низкой устойчивостью к неблагоприятным воздействиям, и в результате – пониженной жизнеспособностью. Параметры стресс-реакции при экспериментальных температурных и токсических стрессах свидетельствуют о снижении согласованности компонентов защитной неспецифической реакции у метизированных особей по сравнению с особями из родительских семей, относящихся к различным расам. Особенности стресс- реакции в гибридных семьях свидетельствуют о вероятном разрушении коадаптированных генетических комплексов родительских рас. Обнаруженные особенности параметров стресс-реакции могут быть использованы в тестах для оценки состояния и породной принадлежности семей медоносной пчелы на пасеках.
Список литературы Проблемы интродукции и внутривидовой гибридизации Apis mellifera
- Гилмур, Д. Метаболизм насекомых. -М.: Мир, 1968. -230 с.
- Никоноров, Ю.М. Использование метода ПЦР для контроля чистопородности пчелосемей Apis mellifera mellifera L. в условиях Южного Урала/Ю.М. Никоноров, Г.В. Беньковская, А.В. Поскряков и др.//Генетика. -1998. -Т.34, №11. -С. 1574-1577.
- Сатаров, В.Н. Популяционно-генетический полиморфизм башкирской популяции медоносной пчелы Apis mellifera L. Дис… канд.биол.наук. -СПб-Пушкин, 2000. -136 с.
- Шакиров, Т.Д. Нужны не слова, а дела//Пчеловодство. -1987. -№12. -С. 9.
- Cornuet, J.M. Pupative origin and function intergenic region between COI-COII of Apis mellifera L. mitochondrial DNA/J.M. Cornuet, L. Garnery, M. Solignac//Genetics. -1991. -V. 128. -P. 393-403.
- Hall, G.H. PCR amplification of a locus with RFLP alleles specific to African honey bees//Biochem. Genet. -1998. -V. 36, No. 9/10. -P. 351-361.
- Smith, D.R. Polymorphisms in mitochondrial DNA of European and Africanized honeybees (Apis mellifera)/D.R. Smith, W.M. Brown//Experientia. -1988. -V. 44. -P. 257-260.
