Проблемы экофизиологической интерпретации климатического сигнала в дендрохронологических и дендроизотопных рядах

Автор: Чуракова О.В., Румянцев Д.Е.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесоведение и лесоводство
Статья в выпуске: 2, 2024 года.
Бесплатный доступ
Важной проблемой современной дендроклиматологии является интерпретация выявленных статистически достоверных связей между изменчивостью метеорологических переменных (среднемесячная и дневная температура воздуха, ежемесячные суммы осадков текущего и предшествовавшего формированию годичного кольца календарного года) и вариацией дендрохронологических параметров (ширина годичного кольца, максимальная плотность поздней древесины, анатомические характеристики годичного кольца и композиционный состав стабильных изотопов в кольце). Адекватная интерпретация необходима, чтобы понимать, какие экофизиологические механизмы определяют выявленные статистические связи, какую смысловую нагрузку, прогностическую и познавательную ценность имеет эта информация в экологическом, экофизиологическом и экогидрологическом аспектах. Наиболее простым путем надежной интерпретации обнаруженных зависимостей является проведение прямых наблюдений за внутрисезонной динамикой формирования годичного кольца и динамикой окружающей экологической обстановки, что, однако, в большинстве случаев невозможно. В качестве одного из вариантов решения этой научной задачи предлагается применять экофизиологические модели в сочетании с прямыми измерениями δ13C, δ18O и δ2H в древесине и целлюлозе годичных колец.
Дендрохронология, дендроклиматология, дендроизотопология, экофизиология растений, лесоведение
Короткий адрес: https://sciup.org/143182677
IDR: 143182677 | УДК: 630.561.24 | DOI: 10.24419/LHI.2304-3083.2024.2.02
Challenges of eco-physiological interpretation of the climatic signal in dendrochronological and dendroisotope time series
Interpretation of statistically significant relationships between meteorological parameters like average or daily monthly air temperature, the sum of precipitation of the previous and current year of the tree-ring formation versus variability of dendrochronological parameters (e.g., tree-ring width, maximum latewood density, anatomical parameters of tree rings, and variability of stable isotopes in tree rings) is crucial for the modern dendroclimatology. After the identification of statistically significant correlations, it is necessary to interpret them in a biologically competent manner to adequately understand what eco-physiological mechanisms are behind them, and what meaningful load, prognostic, and cognitive value this information has in ecological, eco-physiological, and eco-hydrological aspects. Like any separately taken biological object, trees of temperate latitudes have their specificity, which differs in connection with hereditary properties of species and physiographic tree growth conditions. The simplest way to reliably interpret the discovered dependencies is to conduct direct observations of the intra-seasonal dynamics of annual tree-ring formation and the dynamics of the surrounding environmental conditions, which is impossible in most cases. Eco-physiological models combined with direct measurements of δ13C, δ18O, and δ2H 18 in wood and tree-ring cellulose are proposed as one of the options for solving this scientific problem.
Текст научной статьи Проблемы экофизиологической интерпретации климатического сигнала в дендрохронологических и дендроизотопных рядах
Дендроклиматология как наука ориентирована, прежде всего, на реконструкцию климата прошлых столетий и тысячелетий по ширине годичных колец деревьев (ШГК) и в меньшей степени учитывает иные параметры их изменчивости. Для этого на основе имеющихся метеорологических наблюдений и временных рядов дендрохронологических показателей (ширины годичного кольца, параметров анатомической структуры толщины клеточной стенки, максимальной плотности зоны поздней и ранней древесины, вариации стабильных изотопов в целлюлозе для калибровочного периода) составляется регрессионное уравнение и выявляется связь между изменчивостью параметров годичных колец и вариацией метеопараметров за период калибровки, а далее те метеопараметры, которые оказывают наиболее значимое воздействие (p < 0,0001), включают в уравнение. Полученная модель верифицируется с имеющимися данными метеорологических наблюдений. Если регрессионное уравнение достоверно, как для калибрационного, так и для верификационного периодов, то строится реконструкция в глубь веков для метеорологического параметра, оказывающего наибольшее влияние на радиальный прирост деревьев [1, 2]. Ценность климатических реконструкций не отменяет самоценность дендроклиматического анализа как такового. В случае полного достижения своих целей он позволяет понять экофизиологические процессы и механизмы, обусловливающие влияние на рост и структуру годичных колец деревьев метеорологических факторов, а также изменчивости изотопной композиции в древесине или целлюлозе, вечной мерзлоты и влаги, продолжительности солнечного сияния, накопления углерода и процессов газообмена на уровне листа. Понимание данных процессов важно для интерпретации полученных статистических регрессионных зависимостей и климатических реконструкций.
Некоторые статистически значимые связи между биометрическими и анатомическими характеристиками годичных колец, стабильными изотопами и климатическими параметрами вне вегетационного периода (сезона роста) не всегда объяснимы по одному параметру. Так, например, ширина годичных колец деревьев из Субарктики Евразии отражает изменения температуры воздуха июня–июля [1]. В то время как анатомические параметры структуры годичного кольца (например, толщина клеточной стенки) регистрируют сигнал температуры воздуха июня–августа [2]. Стабильные изотопы в годичных кольцах деревьев позволяют, помимо летней температуры воздуха, выявить влияние летних осадков [3–5], продолжительности солнечного сияния [6, 7], дефицита упругости водяного пара [7, 8], относительной влажности воздуха [7], арктического колебания (AO).
Арктическое колебание через атмосферную циркуляцию воздушных масс оказывает воздействие на рост хвойных видов в центральной части Сибири [9], в то время как на западную европейскую скандинавскую часть влияет североатлантическая циркуляция [10]. Например, арктическое колебание определяет зимние температурные режимы и количество осадков в Евразии и Северной Америке. Положительные фазы арктического колебания (AO+) в мае обусловливают теплые и часто сухие погодные условия в Субарктике Сибири. Исследования выявили, что AO+ демонстрируют более высокую амплитуду колебаний в средневековом периоде по сравнению с современным периодом [9–12]. Однако в течение современного периода частота положительных колебаний более продолжительна. В то время как отрицательные фазы арктического колебания приводили к похолоданиям, вызванным стратосферными вулканическими извержениями в 536, 541–542, 1257, 1812, 1814–1815, 1822, 1835, 1883 гг. н. э., которые отчетливо зафиксированы в реконструкции. В течение XX–XXI вв. колебания АО не превышают амплитуды второй половины первого тысячелетия и средневекового потепления (IX–X вв. н.э.) [9].
Сравнение региональной реконструкции АО, полученной для Субарктики Сибири, с реконструкцией Североатлантического колебания (NAO) выявило основной источник изменчивости зимней атмосферной циркуляции в Северном полушарии. Хронологии значительно согласуются в основном в течение Малого ледникового периода (LIA, 1350–1800 гг. н.э.). Как положительная аномалия во время позднеантичного Малого ледникового периода (LALIA, 516–600 гг.), так и раннесредневековая климатическая аномалия (MCA, 700–1000 гг. н.э.) значимо регистрируются в сезонном реконструированном индексе AO [9].
Междисциплинарный подход и использование различных косвенных параметров позволяют получить более расширенную информацию об изменении климата и более качественные и статистически достоверные климатические реконструкции. Однако экофизиологическая интерпретация сигнала остается ограниченной в силу отсутствия прямых измерений для удаленных и труднодоступных участков, где их ежедневное проведение в течение вегетационного периода крайне затруднительно. Кроме того, в отдельных случаях инструменты и приборы с таких участков исследования забирают местные жители либо их повреждают дикие животные. Для районов исследования с более благоприятными условиями проводят прямые измерения, хотя экофизиологическая интерпретация результатов остается недостаточной и базируется в основном на статистическом анализе, в лучшем случае с привлечением экологических или процессных моделей [2, 13–15].
Основной проблемой при экофизиологиче-ской интерпретации результатов статистического анализа влияния климатических факторов на формирование дендрохронологических параметров, на наш взгляд, является отсутствие формализованных алгоритмов. Как правило, объяснения экофизиологического механизма связей ведутся произвольно и субъективно. В основе трактовок может лежать сопоставление результатов данного конкретного исследования с результатами иных дендроклиматических исследований (для других видов и регионов), а также построение концептуальных экофизио-логических моделей (либо трактовка на основе существующих экофизиологических моделей).
Хорошим методическим подспорьем в этой ситуации может быть включение в анализ результатов исследований по стабильным изотопам, уточняющих наше понимание конкретных физиологических процессов в организме дерева.
Концептуальная классическая модель связей в системе «ширина годичного кольца» (ШГК) и «климатический параметр» была предложена Фритцем [16] и сформулирована в форме трех основных подсистем: (1) влияние малого количества осадков и высоких температур на формирование узких годичных колец; (2) влияние малого количества осадков и высоких температур предшествующего вегетационного периода на формирование узкого годичного кольца данного года и (3) влияние большого количества осадков и низких температур на формирование узких годичных колец.
Экофизиологическая модель изменчивости прироста Е.А. Ваганова с соавт. [2] прежде всего была предназначена для работы с деревьями из зоны лесотундры, где на прирост влияет ограниченное число лимитирующих климатических факторов, в основном температура воздуха июня и июля. В настоящее время модель апробирована и нашла широкое применение для многих районов исследования по всему земному шару. Однако полученные результаты для европейской части Евразии и Африки все еще нуждаются в более тщательной экофизиологической интерпретации. В принципе, наибольшая часть работ в области дендроклиматологии посвящена исследованиям деревьев, произрастающих в экстремальных климатических условиях в зоне вечной мерзлоты. На протяжении многих лет это был традиционный объект дендроклима-тических исследований [1, 2]. Достоинствами биома лесотундры с точки зрения организации дендроклиматических исследований являются относительно медленный рост деревьев, высокая продолжительность их жизни, хорошая сохранность отмершей древесины в условиях климата данного биома, что позволяет получать сверхдолгосрочные хронологии, а также наличие ярко выраженного в своем влиянии экологического лимитирующего фактора (температуры воздуха в течение вегетационного периода), который определяет все процессы жизнедеятельности в организме дерева. Ярко выраженный температурный сигнал легко выявляется методами статистического анализа дендрохронологических параметров и без затруднения экофизиологически интерпретируется.
Иная ситуация складывается при дендрокли-матическом анализе роста деревьев в неэкстремальных условиях произрастания. Он затруднен в силу того, что лимитирующие радиальный прирост деревьев факторы меняются от года к году. Поэтому в лесной зоне, по определению представляющей наибольший интерес для специалистов в области лесоведения, как правило, наблюдается ограниченное количество корреляций между дендрохронологическими параметрами и метеопараметрами, статистические связи слабые и значения коэффициентов корреляции в основном ненамного превышают пороговые значения, при которых их можно считать статистически достоверными. Однако метод дендроклиматиче-ского анализа на основе климатограмм показывает, что минимумы и максимумы радиального прироста в этих условиях имеют ярко выраженную климатическую обусловленность [17]. При этом формирование узкого годичного кольца, например у ели в условиях Центрально-Лесного заповедника, может быть вызвано как избытком осадков в течение вегетационного периода, так и их недостатком [17–20], что хорошо соотносится с результатами, предсказанными концептуальной экофизиологической моделью Фритца [16].
С точки зрения классических представлений экологии, в той части ареала вида, в которой ежегодные значения климатических факторов близки к составляющим зону оптимума, статистически значимые отклонения от величины средних значений воздействия экологических факторов, как в большую, так и в меньшую сторону, отрицательно сказываются на продуктивности вида, индикатором которой является ширина годичного кольца. Наличие одинаковой реакции прироста деревьев на разнонаправленные экстремальные отклонения факторов от среднего значения в совокупности со сменой лимитирующего фактора от года к году обусловливает слабые по тесноте корреляционные связи между динамикой радиального прироста и изменениями метеопараметров.
Следует отметить, что в настоящее время дендроклиматические исследования не ограничиваются использованием такого показателя, как ширина годичного кольца и производных от него показателей, полученных путем математических преобразований исходных временных рядов радиального прироста. Использование современного оборудования ведет к детализации исследований и, соответственно, выявляются новые зависимости в системе «радиальный прирост – климат», требующие экофизиологического обоснования. Исследование стабильных изотопов в годичных кольцах деревьев и применение концептуальных моделей [21] позволяет лучше понять, как деревья реагируют на окружающую среду. Изучение соотношения изотопов углерода (13С/12С), кислорода (18О/16O) и водорода (2H/1H) в годичных кольцах стало мощным инструментом для выявления функциональных реакций деревьев на изменение окружающей среды [23–27]. Этот подход опирается на установленные взаимосвязи между газообменом листьев и изотопным фракционированием для получения серии модельных сценариев, которые могут быть использованы для выводов об изменениях фотосинтетической ассимиляции и устьичной проводимости, вызванных динамикой параметров окружающей среды (CO2, доступность воды, влажность воздуха, температура, питательные вещества) [28–31]. В тех случаях когда результаты исследований вариаций стабильных изотопов отклоняются от физиологически правдоподобных выводов, такого рода несоответствие между газообменом и изотопным откликом обращает на себя внимание и может позволить получить новые сведения о механизмах, лежащих в основе протекания физиологических процессов в организме дерева.
Цель исследования – рассмотреть проблему экофизиологической интерпретации климатического сигнала во временны´х рядах дендрохронологических параметров на примере использования для этих целей анализа стабильных изотопов и их вариации в древесине годичных колец. На наш взгляд, только совместный подход дендрохронологов и экофизиологов поможет привлечь внимание к проблеме поиска путей формализации процессов экофизиологической интерпретации дендроклиматического сигнала в разнообразных дендрохронологических рядах: начиная от временных рядов анатомических структур ширины годичных колец до временных рядов изотопной композиции стабильных изотопов в годичных кольцах.
Методы исследования
Методы сбора дендрохронологических образцов, их предварительной обработки, измерения ширины годичных колец, построения датированных древесно-кольцевых хронологий хорошо изучены и подробно описаны, разработаны общепринятые методические подходы для решения этих задач [1, 2, 17–20, 32].
Для анализа стабильных изотопов в годичных кольцах первоначально перекрестно-датированные образцы нарезают при помощи скальпеля под бинокуляром для каждого года и каждого дерева отдельно и затем упаковывают в термостойкие пакеты для экстракции целлюлозы. Затем выполняют комплекс физико-химических исследований, направленных на анализ стабильных изотопов углерода, кислорода и водорода.
Каждый образец древесины помещают в пакетик для фильтрации (F57, Ankom Technology, NY, USA), который дважды промывают в течение 2 ч в 5 %-м NaOH для удаления липидов, смол и гемицеллюлозы. Далее в течение 36 ч проводится обработка 7 %-м NaClO2 для удаления лигнина [33]. Образцы промывают и высушивают в течение 24 ч при температуре 50 °C, затем гомогенизируют с помощью ультразвука. Каждый образец целлюлозы из древесных колец взвешивают (около 1,0 мг) и упаковывают в серебряные или оловянные капсулы для анализа стабильных изотопов.
Для изотопного анализа водорода необходимо исключить влияние обменного водорода. Существует два подхода: а) нитрование целлюлозы или б) уравновешивание целлюлозы и воды известного изотопного состава с последующей процедурой коррекции для получения (S 2 H) водорода, связанного с углеродом. В данной работе мы придерживались второго подхода, основанного на новом протоколе, подробно описанном в работе [34]. Если объяснить в нескольких словах – образцы уравновешиваются горячим водяным паром (130 °C) в закрытой камере в течение 2 ч, а затем их высушивают с помощью N2 в той же камере при той же температуре. Далее образцы переносят в автодозатор пиролизного устройства, который непрерывно промывается аргоном, и преобразуют путем высокотемпературной конверсии при 1 420 °C в H2 с последующим анализом на масс-спектрометре (IRMS) (MAT 253, Thermo, Германия) [6].
Образцы целлюлозы (0,2–0,3 мг) взвешивают в оловянных капсулах для анализа 13C/12C с помощью масс-спектрометра изотопного соотношения delta-S (Finnigan MAT, Бремен, Германия), соединенного с двумя элементными анализаторами (EA-1110 Carlo Erba, Италия) через интерфейс с переменным открытым разделением (CONFLO-II, Finnigan MAT, Бремен, Германия). Содержание 13C/12C определяют путем сжигания в избытке кислорода при температуре реактора 1 020 °C, работающего в режиме непрерывного потока.
Кроме того, их можно анализировать в серебряных капсулах (около 1 мг) с помощью vario PYRO (Elementar, Ханау, Германия) путем термического разложения при 1 450 °C и преобразования в CO при исключении присутствия O2 в среде из гелия [6]. Эта система связана с IRMS (Delta plus XP, Thermo Finnigan, Бремен, Германия). Во время пиролиза целлюлозы к образовавшемуся СО добавляется небольшое, но постоянное количество углерода, который используется для изотопного анализа как углерода, так и кислорода. Были проведены многочисленные тесты для определения функции коррекции этого эффекта путем анализа более 100 образцов (513C скорректированный = 1,1142 - S13C исходный + 1,45)
[35], а также анализа внутренних стандартов, известного изотопного состава с каждой последовательностью для проверки этой коррекции. Межлабораторное сравнение между 9 европейскими исследовательскими лабораториями [36] показало хорошую согласованность между результатами в пределах точности используемого метода масс-спектрометрии изотопного соотношения (IRMS): (0,2 ‰ для углерода и 0,3 ‰ для кислорода).
Системы EA-IRMS и PYRO показывают очень высокую точность (± 0,2 ‰), и значения, полученные с помощью двух приборов, являются статистически достоверными [6]. Точность (±0,1 ‰ для δ13C, ±0,3 ‰ для δ18O и ±2 ‰ для δ2H) основана на большом количестве измерений стандартного материала международных эталонов (n = 99).
Изотопные соотношения выражаются в обычной дельта-нотации (δ) относительно международных стандартов, где: Rsample – отношение 13C/12C, или 18O/16O, или 2H/1H в образце, а Rstandard – отношение либо для углерода (13C/12C) в Венском белемните Пи-Ди (VPDB), либо для кислорода (18O/16O) и водорода 2H/1H в Венской стандартной средней океанической воде (VSMOW).
Поправка на изменение δ13C атмосферного CO2 [37] применяется ко всем измерениям изотопного соотношения углерода в целлюлозе годичных колец. В то время как к δ18O и δ2H никаких специфических поправок не применяется.
Детальное описание методик дендроизотоп-ного анализа содержится в работе О.В. Чураковой с соавт. [6, 23, 24].
Результаты и обсуждение
Рассмотрим экофизиологические процессы, которые значимы для формирования древесины годичных колец и формирования в ней разных концентраций стабильных изотопов углерода, кислорода и водорода.
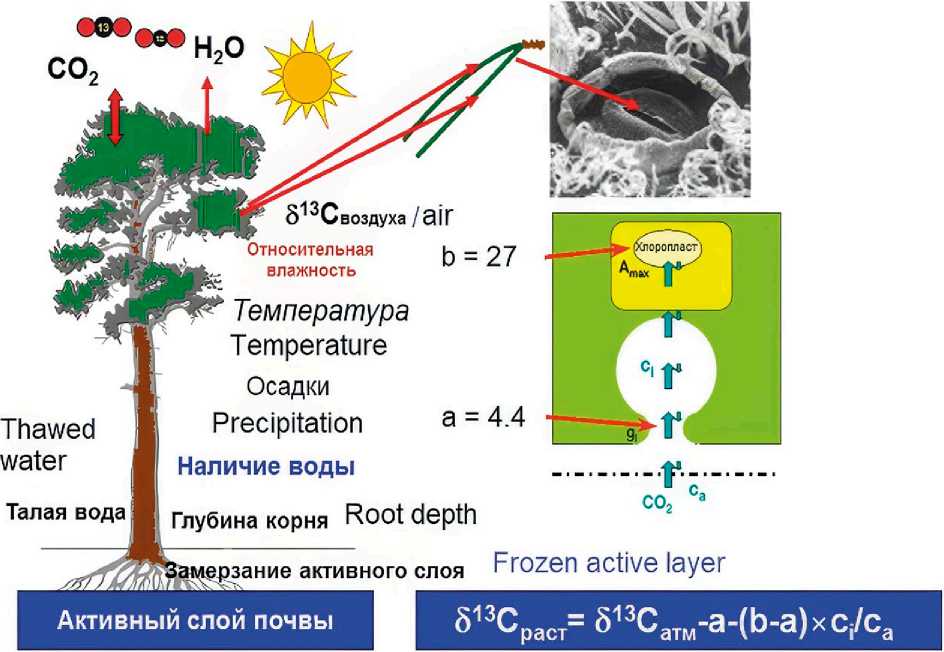
Дискриминация углерода (13C/12C). Известно, что на рост дерева влияет множество факторов, включая осадки, температуру, источник воды, относительную влажность, дефицит упругости водяного пара и солнечную радиацию. Изотопное соотношение углерода (13C/12C, или δ13C) в кольцах деревьев отражает сигналы о наличии воды и влажности воздуха, как результат воздействия климата через фотосинтез. В процессе фотосинтеза происходит несколько этапов фракционирования, первый – когда CO2 поступает из атмосферы (ca) и диффундирует в межклеточные пространства листа (хвои) (ci), второй – в процессе фиксации CO2 RuBisCo [23–26]. Открытие и закрытие устьиц (gs) определяет водный контроль. В тёплых и сухих условиях деревья реагируют на ограниченные водные ресурсы снижением устьичной проводимости (gs), что приводит к уменьшению поглощения CO2 и производства биомассы, а также к снижению концентрации межклеточного CO2 (ci), и наоборот. Таким образом, изменение скорости ассимиляции будет влиять на межклеточную концентрацию CO2 (ci) через изменение скорости утилизации CO2 для образования сахаров, а увеличение или уменьшение устьичной проводимости будет воздействовать на скорость восполнения этого внутреннего CO2 (ci). Деревья дискриминируют 13С в условиях высокого содержания СO2 в межклеточном пространстве в листе/хвое (ci), когда устьица относительно широко открыты или фотосинтез находится на низком уровне. При повышении концентрации CO2 в атмосфере уменьшается обмен водяного пара между хвоей и окружающим воздухом и снижается устьичная проводимость [25].
На рис. 1 представлено влияние на дискриминацию углерода таких факторов, как: температура воздуха, атмосферные осадки, относительная влажность, наличие талой воды, глубина корней и промерзание почвы (ее активного слоя), а также схематическая модель фотосинтеза и потока СО2 во внутриклеточное пространство листа/ хвои через устьица.
Фракционирование кислорода (18O/ 16O). Фракционирование изотопов кислорода – это более сложный процесс по сравнению с дискриминацией углерода. Вариация 18O/16O (δ18O) в годичных кольцах деревьев фиксирует изотопный
Active soil layer
Рис. 1. Модифицированная версия схематической диаграммы дискриминации углерода:
а – дискриминация относительно 13СО2 в результате диффузии в межклеточное пространство листа через устьице (~ -4.4 %o), b - дискриминация в результате карбоксилирования (~ -27 %о ), С . - ВНУТРЕННЯЯ И С а - ВНЕШНЯЯ концентрация С02, д . - УСТЬИЧНАЯ ПРОВОДИМОСТЬ [25].
состав относительно температуры воздуха и осадков [30, 38, 39], которые представлены в виде источника воды для деревьев (рис. 2).
Кислород проникает в клетку и вовлекается в цикл Кальвина через СО2. Однако до того как CO2 будет доступен для клетки, он диссоциирует в воде, покрывающей межклеточные мембраны клеток. В этой диссоциированной форме происходит обмен атомами кислорода между молекулами CO2 и H2O, и таким образом в CO2 переходит атом кислорода из H2O, и наоборот (с коэффициентом фракционирования около 41 %о) [26, 38]. Поскольку количество и потоки воды, направленные в организм дерева, превышают потоки CO2 в 100–500 раз, то в химическом составе древесины годичного кольца проявляется преимущественно изотопное влияние воды на CO2, в то время как О-сигнал, источником которого является CO2, едва заметен в воде. Это очевидный механизм, функционирующий в силу того, что наблюдается разница в массах поступающих веществ (как упоминалось выше, масса воды в 100–500 раз превышает массу CO2 (100-500 * H2O / 1 х CO2). Поэтому молекула CO2 включает в себя изотопы кислорода воды до того момента, как она вступает в цикл Кальвина и превращается в глюкозу и затем в другие органические вещества. Другими словами, после того как CO2 диффундирует в межклеточные пространства листа, он впоследствии диссоциирует в водной пленке, покрывающей клеточные мембраны. Поэтому изотопный сигнал кислорода воды из тканей листа присутствует в ассимилятах, чего в принципе быть не должно, так как, согласно реакции Хилла, кислород воды выделяется в атмосферу в качестве «отхода» фотосинтеза. Соответственно, это находит отражение
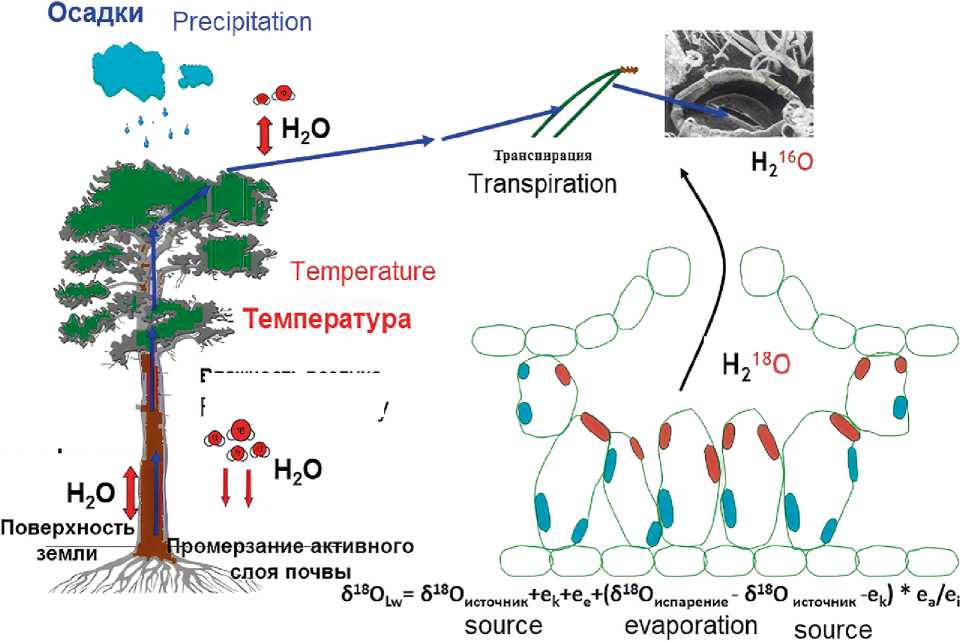
Рис. 2. Модифицированная версия схематической диаграммы изотопного фракционирования между осадками и хвоей [24, 29–31, 40, 42]:
e k – кинетическое фракционирование; ea и ei – атмосферное и внутриклеточное давление пара
Evaporation-
Испарение
Влажность воздуха
Relative humidity
в наблюдаемом химическом составе целлюлозы [29–31, 40].
Вода поступает из почвы (источник воды, талая мерзлотная вода либо грунтовая вода) через ксилему к хвое, где легкие изотопы кислорода (16O) испаряются быстрее по сравнению с более тяжелыми (18O), в результате чего происходит насыщение воды, находящейся в листе, изотопом 18O. Обогащение 18O может осуществляться особенно интенсивно в условиях засухи [24, 40].
Изотопная комбинация δ18O – источника воды, используемая корнями деревьев, дополнительно определяется эвапотранспирацией в листе/хвое δ18OLW и отражает информацию о наличии влаги. Атмосферная вода, содержащая сигнал об изменении δ18O, напрямую зависит от влажности/температуры воздуха [41], так же как эвапотранспирация и процессы конденсации в глобальных гидрологических циклах [42, 43].
Вода, как входной параметр, модифицирована степенью насыщения 18O в хвое (H218O) вследствие транспирации (легкие изотопы кислорода 16O испаряются быстрее), которая включена в фотосинтез и формирование целлюлозы через биохимическое фракционирование и процессы обмена.
Фракционирование водорода (2H/1H). Изотопный состав водорода в годичных кольцах деревьев все еще мало изучен по сравнению с исследованиями 2H в воде [23, 24, 29]. Вариация 2H /1H (δ2H) в годичных кольцах деревьев зависит от происхождения источника воды, например: осадки, водяной пар, грунтовая вода или талая мерзлотная вода. Чем выше широта (эффект широты) и высота над уровнем моря
(эффект высоты) (Rayleigh-эффект), а также чем более континентальной является область формирования облаков, тем более обеднёнными 2H будут осадки [41]. Поверхностная вода становится 2H-обогащённой из-за более активного испарения изотопно более лёгкой воды. Вода, просачивающаяся в более глубокие слои почвы, перемешивается с водой предыдущих осадков, которая часто бывает обеднена 2H по сравнению с поверхностной водой, так как не испаряется. Изотопный состав грунтовых вод может сильно отличаться от дождевых вод из-за того, что они могут иметь другую площадь водосбора и другое временное происхождение. В зависимости от глубины залегания корней и скорости транспирации вода в ксилеме дерева может иметь смешанный
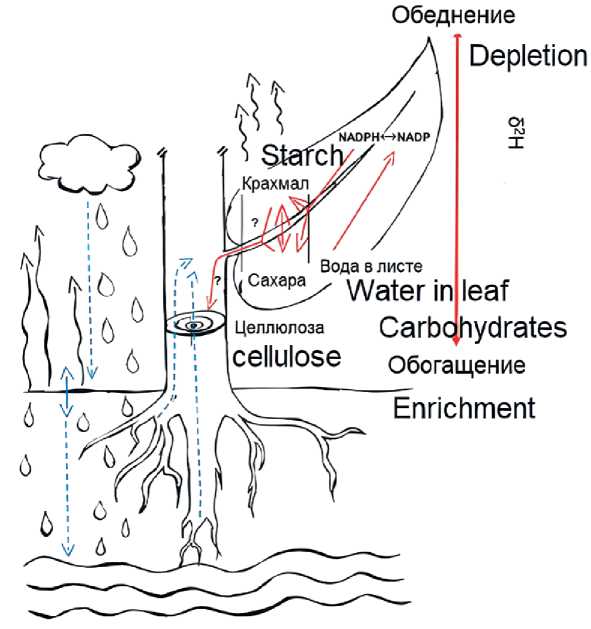
Рис. 3. Схема изотопного состава водорода в кольцах деревьев (модифицирована после Lehmann et al. [27]): Синие линии относятся к воде, пунктирными линиями обозначены процессы, в которых 2H-фракционирование незначительно или отсутствует. Красные линии и ось относятся только к 2H-фракционированию в органических молекулах, включая NADPH и углеводы (т.е. сахара, крахмал и целлюлозу).
изотопный состав почвы и грунтовых вод, при этом дальнейшее 2H-фракционирование в процессе поглощения обычно не происходит [29]. Вода в листе или хвое обогащается 2H за счёт транспирации изотопно более лёгкой воды. На этот эффект могут влиять изменения устьичной проводимости, температуры и влажности воздуха (рис. 3).
На рис. 3 показано, как обогащенная 2H вода в листьях/хвое и обедненный 2H никотинамида-дениндинуклеотидфосфат (NADPH), полученный при фотолизе H2O, являются основными изотопными источниками первичных ассимилятов. Неразрешенные процессы 2H-фракционирова-ния, связанные с фиксацией углерода, синтезом крахмала, ремобилизацией и последующим транспортом сахаров, приводят к 2H-обогащению целлюлозы древесных колец по сравнению с асси-милятами листьев.
Большинство исследований, проведенных в Субарктике Евразии, выявили отрицательно значимые корреляции между стабильными изотопами углерода (13C) в годичных кольцах и летними осадками, что свидетельствует о засушливых условиях в данных регионах [3–6, 8, 15, 23]. Во время засухи устьица закрываются, чтобы предотвратить потерю воды. Медленное увеличение cа и ci может привести к дальнейшему снижению устьичной проводимости при сохранении интенсивности фотосинтеза и радиального прироста деревьев на прежнем уровне. Снижение устьичной проводимости отражается на увеличении значений δ18O, поскольку обогащение воды в листьях H218O снижается в меньшей степени за счет конвективного потока обедненной исходной воды (через ксилему), чем при более высоких скоростях транспирации (эффект Пекле) [38].
Увеличение температуры воздуха и продолжительности солнечного сияния оказывает значимое воздействие на почву в Субарктике. Понижение суммы выпадающих осадков ведет к более глубокому оттаиванию верхнего активного слоя почвы. Тающая мерзлотная вода сильно обеднена 18O, поэтому может приводить к снижению значений кислорода в годичных кольцах деревьев [6, 23]. Однако талая мерзлотная вода не может сохраняться длительное время и теряется за счет испарения и стока в ручьи и реки, что приводит к засухе в активном слое почвы и, как следствие, к снижению прироста деревьев, как это наблюдается в зоне средней тайги [6, 23]. В этом случае дождевая вода, которая изотопно тяжелее талой мерзлотной воды, может играть ключевую роль в качестве источника воды для деревьев [44]. Точное положение мерзлоты имеет решающее значение для роста деревьев и может быть губительным, если оно слишком низкое или слишком высокое.
Положительные корреляции между хронологиями изотопов углерода и летними температурами воздуха (как правило июня и июля) для северной части Евразии вполне закономерны, поскольку июнь и июль – самые теплые месяцы, и такие связи указывают на увеличение фотосинтетической активности и дефицита упругости водяного пара [8]. Теплые и сухие условия вызывают закрытие устьиц и указывают на снижение изотопного фракционирования, что приводит к повышению значений δ13C. Увеличение дефицита упругости водяного пара связано с уменьшением устьичной проводимости и с дальнейшим увеличением обогащения δ18O воды в листьях. Даже в самый теплый месяц (июль) почвенная вода на глубине 20–30 см находится в замерзшем состоянии [15]. Талая вода остается холодной и не может быть использована корнями при низкой температуре почвы [15]. Таким образом, доступность воды для деревьев ограниченна, что может привести к засухе. Очевидно, что возникновение засухи сильно зависит от местных почвенных условий, поэтому обобщение результатов, полученных на одном участке, применительно к большим территориям затруднительно.
Приходящая солнечная радиация и ее сезонное распределение играют важную роль для деревьев с точки зрения фотосинтеза, производства углеводов и, впоследствии, соотношения изотопов углерода в органическом веществе [6, 7, 21]. Увеличение фотосинтетически активной радиации в целом приводит к увеличению значений δ13C, особенно если при этом в системе «дерево–среда» вода не является лимитирующим жизнедеятельность фактором [45]. Результаты, полученные по бореальной зоне лесов (50–70° c.ш. – 12–149° в.д.; 133–148° з.д.), показывают значимую статистическую связь между стабильными изотопами углерода, а также кислорода и водорода с ШГК и с продолжительностью летнего солнечного сияния в течение вегетационного периода [6, 7, 23]. Помимо хорошо известных δ13C и δ18O, менее изученный δ2H в целлюлозе годичного кольца показывает отрицательную корреляцию с продолжительностью солнечного сияния в течение осенне-зимних месяцев [6, 7, 23]. Такая связь может быть обусловлена недостатком света во время полярных ночей для регионов, расположенных в высоких широтах (> 60° с.ш.), а для лиственницы также и с отсутствием хвои для производства продуктов фотосинтеза в холодной среде. Могут возникать и косвенные эффекты, так как большое количество зимнего солнечного света означает наличие устойчивых систем высокого давления с малым количеством осадков, что негативно сказывается на снегонакоплении, которое является важным источником воды после таяния снега. Снижение поступающей солнечной радиации наблюдается также в осенние и зимние месяцы в западной части бореальной зоны, где произрастают вечнозелёные виды хвойных деревьев. Эти деревья потенциально могут производить фотоассимиляты и превращать их в органическое вещество, когда почва ещё не промёрзла, а температура воздуха составляет около +5 °C (температурный порог) [45], и поэтому их дендрохронологические ряды показывают значимые корреляционные связи с динамикой стабильных изотопов.
Гидрологический режим бореальных лесов Северного полушария довольно сложен и представляет собой синусоиду сезонного хода изотопов воды, зафиксированных в осадках (δ18O и δ2H), которые отражают температуру конденсации облаков [41, 42, 46]. Поглощение зимних осадков возможно в весенние и летние месяцы после таяния снега и оттаивания активного слоя почвы. Это подтверждается значимыми корреляциями между δ18O в годичных кольцах и зимневесенними температурами воздуха. Изотопы кислорода в органическом веществе изменяются под влиянием вариаций изотопного состава исходного источника воды, который тесно связан с изотопным составом осадков в почвенной воде (хотя и модифицируется испарением на поверхности почвы) [29, 30, 43].
Зимние изменения температуры воздуха, например, хорошо улавливаются сосной обыкновенной в Скандинавии, а зимние изменения влажности – лиственницами в Сибири, что может быть связано с закономерностями арктического колебания через зимне-весеннюю относительную влажность и изменчивость осадков. Более раннее исследование для сибирского участка (полуостров Таймыр) показало влияние арктического колебания в весенний период на изменчивость стабильного изотопа кислорода за последние тысячелетия [6, 9], что может быть обусловлено засушливым летом в последние десятилетия. Напротив, летние штормы Северной Атлантики и Северо-Атлантической осциляции могут влиять на хвойные деревья в Скандинавии, повышая относительную влажность воздуха [10].
Стабильные изотопы углерода, кислорода и водорода могут обеспечить исчерпывающей информацией о климатических факторах не только в течение летних месяцев, но и в зимние периоды. Данный сигнал вряд ли можно выявить по таким классическим древесно-кольцевым хронологиям бореальных хвойных пород (ширина кольца, плотность поздней древесины). Однако интерпретация результатов для этих регионов с их сложной гидрологией не является прямолинейной из-за часто неоднозначной совокупности сигналов, регистрируемых годичными кольцами деревьев. Экофизиологическое моделирование и выявление механизмов, оказывающих влияние на вариацию стабильных изотопов в годичных кольцах, позволит более четко определять значимость влияния различных климатических факторов [47].
Результаты наших исследований показывают значимое влияние летних температур воздуха, влажности и продолжительности солнечного сияния на рост деревьев по всей бореальной зоне лесов.
Динамика засушливых условий в течение вегетационного периода обычно тесно связана с сочетанием таких метеорологических параметров, как: температура воздуха, осадки, дефицит упругости водяного пара и эвапотранспирация, – которые являются наиболее важными для регионов с отсутствием вечной мерзлоты. В то время как вечномерзлотные почвы служат резервуаром воды и ее дополнительным источником поступления в засушливые годы для хвойных пород в Сибири [6, 23, 44]. Естественно, подобного рода экофизиологические эффекты не наблюдаются и не фиксируются в дендрохронологических параметрах в регионах, где вечномерзлотные почвы отсутствуют, например, в Скандинавской Субарктике.
Однако наблюдаемая неоднородность реакции на климат может быть объяснена специфическими для конкретного места и вида различиями, например, между вечнозелёными ( Pinus в Скандинавии и Picea в Западной Канаде) и листопадными видами (например, Larix в Сибири) [6, 23].
С нашей точки зрения, для современной дендроклиматологии и дендроизотопологии наиболее актуальна проблема поиска путей формализации процессов экофизиологической интерпретации климатического сигнала в параметрах годичных колец деревьев и изотопном составе в целлюлозе годичных колец деревьев. Применение экофизиологических моделей в сочетании с прямыми измерениями δ13C, δ18O и δ2H в древесине и целлюлозе годичных колец поможет выявить и разделить влияние на прирост деревьев таких гидроклиматических факторов, как атмосферные осадки или почвенная влага.
Заключение
Использование в дендрохронологических исследованиях данных о вариациях трёх важнейших изотопов (δ13C, δ18O и δ2H), несомненно, дополнит методологическую базу для моделирования значений метеопараметров в целях их реконструкции и прогноза на основе дендрохронологической информации и поможет улучшить качество экогидрологических реконструкций за последние тысячелетия. В общем и целом дендро-изотопный анализ расширяет возможности для экофизиологической интерпретации результатов дендроклиматического анализа, так как позволяет непосредственно исследовать физиологические процессы, протекающие в дереве. Важно, что эти исследования могут быть выполнены для старовозрастных деревьев, произрастающих в конкретных условиях определенной экосистемы и испытывающих все многообразие воздействия типичных абиотических и биотических факторов с их аддитивными, кумулятивными и компенсационными эффектами при взаимодействии друг с другом в ходе влияния на процессы формирования годичных колец древесины.
Список литературы Проблемы экофизиологической интерпретации климатического сигнала в дендрохронологических и дендроизотопных рядах
- Ваганов, E.A. Дендроклиматические исследования в Урало-Сибирской Субарктике / Е.А. Ваганов, С.Г. Шиятов, В.С. Мазепа. - Новосибирск: Наука, 1996. - 246 с.
- Vaganov, E.A. Growth dynamics of conifer tree rings. Image of past and future environments / E.A. Vaganov, M.K. Hughes, A.V. Shashkin. - Berlin: Springer Verlag, 2006. - Р. 358. doi:10.1007/3-540-31298-6.
- Isotopic composition (513C, 518O) in Siberian tree-ring chronology. - Текст: электронный / O.V. Sidorova [et al.] // Geophys. Res. Biogeosci. - 2008. - Vol. 113. doi:10.1029/2007JG000473.
- Do centennial tree-ring and stable isotope trends of Larix gmelinii (Rupr.) indicate increasing water shortage in the Siberian north. - Текст: электронный / O.V. Sidorova [et al.] // Oecologia. - 2009. - Vol. 161. - № 4. - Р. 825835. - Режим доступа: https://link.springer.com/article/10.1007/s00442-009-1411-0.
- Spatial patterns of climatic changes in the Eurasian north reflected in Siberian larch tree-ring parameters and stable iso-topes. - Текст: электронный / O.V. Sidorova [et al.] // Global Change Biology. -2010. - № 16. - Р. 10031018. doi:10.1111/j.1365-2486.2009.02008.x
- Climate impacts on tree-ring stable isotopes across the Northern Hemispheric boreal zone. - Текст: электронный / O.V. Churakova (Sidorova) [et al.] // Sci. Total Env. - 2023. - Vol. 870. doi:10.1016/j.scitotenv.2023.161644.
- Dual carbon and oxygen isotopes in Siberian tree rings as indicator of millennia sunshine duration changes. -Текст: электронный / O.V. Churakova (Sidorova), R.T.W. Siegwolf, MS Zharkov, M. Saurer // Science of the Total Environment. - 2024. - Vol. 927. - Режим доступа: https://doi.org/10.1016/j.scitotenv.2024.172042.
- Recent atmospheric drying in Siberia is not unprecedented over the last 1500 years. - Текст: электронный / O.V. Churakova (Sidorova) [et al.] // Sci. Rep. - 2020. - Vol. 10. - Р. 15024.DOI: 10.1038/s41598-020-71656-w.
- Spring Arctic Oscillation as a trigger of summer drought in Siberian subarctic over the past 1494 years. - Текст: электронный / O.V. Churakova (Sidorova) [et al.] // Scientific Reports. - 2021. - Vol. 11. - P. 19010. - Режим доступа: https://doi.org/10.1038/s41598-021-97911-2.
- North Atlantic summer storm tracks over Europe dominated by internal variability over the past millennium. -Текст: электронный / M. Gagen [et al.] // Nat. Geosci. - 2016. - Vol. 9. - P. 630-635. doi:10.1038/ngeo2752.
- Impacts of the Siberian High and Arctic Oscillation on the East Asia winter monsoon: Driving down welling in the western Bering Sea Aquatic Ecosystem. - Текст: электронный / Wang, J. [et al.] // Health Manag. - 2012. - 15(1). -P. 20-30. - Режим доступа: https://doi.org/10.1080/14634988.2012.648860.
- Analysis of the positive Arctic Oscillation index event and its influence in the winter and spring of 2019/2020. -Текст: электронный / J. Zhang [et al.] // Front. Earth Sci. - 2021. - Vol. 8. - P. 1-17. - Режим доступа: https://doi. org/10.3389/feart.2020.580601.
- 20th century changes in carbon isotopes and water-use efficiency: tree-ring-based evaluation of the CLM4.5 and LPX-Bern models / K.M. Keller [et al.] // Biogeosciences. - 2017. - Vol. 14. - P. 2641-2673. doi:10.5194/ bg-14-2641-2017.
- Transient simulations of the carbon and nitrogen dynamics in northern peatlands: from the Last Glacial Maximum to the 21st century. - Текст: электронный/ R. Spahni [et al.] // Clim. Past. - 2013. - Vol. 9. - P. 1287-1308. doi:10.5194/cp-9-1287-2013.
- Application of eco-physiological models to the climatic interpretation of 513C and 518O measured in Siberian larch tree-rings / O.V. Churakova (Sidorova) [et al.] // Dendrochronologa. - 2016. - Vol. 39. - P. 51-59. doi:10.1016/j. dendro.2015.12.008.
- Fritts, H.C. Tree Rings and Climate / H.C. Fritts. - London: Academic Press, 1976. - 567 p.
- Румянцев, Д.Е. Дендроклиматические исследования и лесоведение / Д.Е. Румянцев // Проблемы природопользования и экологическая ситуация в Европейской России на сопредельных территориях. - Белгород: БелГУ, 2017. - C. 58-60.
- Матвеев, С.М. Дендрохронология / С.М. Матвеев, Д.Е. Румянцев. - Воронеж: ВГЛТА, 2013. - 139 с.
- Судебно-ботаническая экспертиза с применением методов дендрохронологии при расследовании незаконной рубки лесных насаждений: учебно-практическое пособие / В.И. Воронин, О.П. Грибунов, Ю.М. Жаворонков [и др.]. - Иркутск: ФГКОУ ВО ВСИ МВД России, 2016. - 200 с.
- Шиятов, С.Г. Дендрохронология верхней границы леса на Урале / С.Г. Шиятов. - Москва: Наука, 1986. - 136 с.
- McCarroll, D. Stable isotopes in tree rings / D. McCarroll, N.J. Loader // Quat. Sci. Rev. - 2004. - Vol. 23. - № 7-8. - Р. 771-801.
- Updating the dual C and O isotope - Gas-exchange model: A concept to understand plant responses to the environment and its implications for tree rings / R.T.W. Siegwolf [et al.] // Plant Cell and Environment. - 2023. -Vol. 46. - Р. 2606-2627. doi:10.1111/pce.14630.
- Stable Isotopes in Tree Rings of Boreal Forests. - Текст: электронный / R.T.W. Siegwolf, J.R. Brooks, J. Roden, M. Saurer. - Corvallis: Springer - 2022. - Vol. 8. - Р. 773. - Режим доступа: https://doi. org/10.1007/978-3-030-92698-4_20.
- Stable isotopes in tree rings inferring physiological, climatic and environmental responses / R.T.W. Siegwolf, J.R. Brooks., J. Roden, M. Saurer // Stable isotope in tree rings. Corvallis: Springer - 2022. - Vol. 8. - Р. 773. doi:10.1007/978-3-030-92698-4.
- Farquhar, G.D. Carbon isotope discrimination and photosynthesis / G.D. Farquhar, J.R. Ehleringer, K.T. Hubick // Annu. Rev. Plant Phisiol. Plant Mol. Biol. - 1989. - Vol. 40. - P. 503-537.
- Farquhar, G.D. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves / G.D. Farquhar, M.H. O'Leary, J.A. Berry // Australian Journal of Plant Physiology. - 1982. - Vol. 9. - P. 121-137.
- The Stable Hydrogen Isotopic Signature: From Source Water to Tree Rings / M.M. Lehmann [et al.] // Stable Isotopes in Tree Rings. Tree Physiology. Corvallis: Springer - 2022. - Vol. 8. Springer doi:10.1007/978-3-030-92698-4_11.
- Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: a conceptual model / Y. Scheidegger, M. Saurer, M. Bahn, R. Siegwolf // Oecologia. - Corvallis: Springer - 2000. - Vol. 125. -Р. 350-357. doi:10.1007/s004420000466.
- Roden, J.S. A mechanistic model for interpretation of hydrogen and oxygen isotope ratios in tree-ring cellulose / J.S. Roden, G.G. Lin, J.R. Ehleringer // Geochim Cosmochim Acta. - 2000. - Vol. 64. - P. 21-35. doi:10.1016/ S0016-7037(99)00195-7.
- Roden, J.S. Hydrogen and oxygen isotope ratios of tree ring cellulose for field-grown riparian trees / J.S. Roden, J.R. Ehleringer // Oecologia. - 2000. - Vol. 123. - P. 481-489.
- Yakir, D. The use of stable isotopes to study ecosystem gas exchange / D. Yakir, L. da S.L. Sternberg // Oecologia. - 2000. - Vol. 123. - P. 297-311.
- Методы дендрохронологии: учебно-методическое пособие. - Ч. I / С.Г. Шиятов, Е.А. Ваганов, А.В. Кирдя-нов [и др.]. - Красноярск: ИЦ КрасГУ, 2000. - 80 с.
- Simultaneous determination of stable carbon, oxygen, and hydrogen isotopes in cellulose / N.J. Loader [et al.] //Anal. Chem. - 2015. - Vol. 87 (1). - Р. 376-380. 10.1021/ac502557x
- A high-temperature water vapor equilibration method to determine non-exchangeable hydrogen isotope ratios of sugar starch and cellulose / P. Schuler [et al.] // Plant Cell Environ. - 2022. - Vol. 45 (1). - Р. 12-22. 10.1111/pce.14193
- Comparison of 518O and 513C values between tree-ring whole wood and cellulose in five species growing under two different site conditions / R.B. Weigt [et al.] // Rapid Commun. Mass Spectrom. - 2015. - Vol. 29 (23). - Р. 2233-2244.
- Wood cellulose preparation methods and mass spectrometric analyses of 513C, 518O, and nonexchangeable 52H values in cellulose, sugar, and starch: an interlaboratory comparison / T. Boettger [et al.] // Anal. Chem. - 2007. - Vol. 79 (12). - Р. 4603-4612.
- A 1000-year high precision record of 513C in atmospheric CO2 / R.J. Francey [et al.] // Tellus Ser. B. - 1999. - Vol. 51. - Р. 170-193.
- Craig, H. Deuterium and oxygen 18 variations in the ocean and the marine atmosphere / H. Craig, L.I. Gordon. -University of California, 1965. - Р. 122.
- Craig, H. Isotopic variations in meteoric waters / H. Craig // Science. - 1961. - Vol. 133. - Р. 1702-1703. doi:10.1126/science.133.3465.1702.
- Reconstructing relative humidity from plant 518O and 5D as deuterium deviations from the global meteoric water line / S.L. Voelker [et al.] // Ecol. Appl. - 2014. - Vol. 24. - Р. 960-975.
- Bowen, G.J. Revenaugh interpolating the isotopic composition of modern meteoric precipitation / G.J. Bowen, J. Revenaugh // Water Resour. Res. - 2023. - Vol. 39. - № 10. - Р. 1299. doi:10.129/2003WR002086
- Dansgaard W. Stable isotopes in precipitation / W. Dansgaard // Tellus. - 1964. - Vol. 16. - P. 436-468.
- The impact of an inverse climate-isotope relationship in soil water on the oxygen-isotope composition of Larix gmelinii in Siberia / M. Saurer [et al.] // New Phyt. - 2016. - Vol. 209. - P. 955-964.
- Importance of permafrost as a source of water for plants in East Siberian taiga / A. Sugimoto, N. Yanagisawa, D. Naito, N. Fujita, T.C. Maximov // Ecol. Res. - 2002. - Vol. 17. - № 4. - Р. 493-503. doi:10.1046/j.1440-1703.2002.00506.x.
- Korner, Ch. The cold range limit of trees/ Ch. Korner // Trends Ecol. Evol. - 2021. - Vol. 36 (11). - Р. 979-989.
- Globally coherent water cycle response to temperature changes during the past two millennia. - Текст: электронный / B.L. Konecky [et al.] // Nat. Geosci. - 2023. - Vol. 16. - Р. 997-1004. - Режим доступа: https://doi. org/10.1038/s41561-023-01291-3.
- A triple tree-ring constraint for tree growth and physiology in a global land surface model / J. Barichivich [et al.] // Biogeosciences. - 2021. - Vol. 18. - Р. 3781-3803. doi:10.5194/bg-18-3781-2021.
