Продукционный процесс как отражение функционирования целостной экосистемы

Автор: Зеленская Надежда Николаевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-4 т.15, 2013 года.
Бесплатный доступ
Структурно-функциональное единство растительности и почвы в экосистеме базируется на метаболизме (циклической смене двух процессов: синтеза органического вещества из минеральных элементов с помощью солнечной энергии автотрофами и деструкции отработавшей ресурс и отмершей биомассы гетеротрофами). Целесообразность такого единства состоит в производстве живой органики, максимально возможной в климатических условиях каждой природной зоны.
Экосистемы, продукционный процесс, фотосинтез, факторы среды
Короткий адрес: https://sciup.org/148201979
IDR: 148201979 | УДК: 581.524
Productyvity process as reflection of functioning of complete ecosystem
The structure-functional unity of vegetable and soil in ecosystem is based on a metabolism (cyclic change of two processes: synthesis life organic substance by autotrophic vegetable and destruction dead organic substance by heterotrophic organism). The aim of ecosystem is a production of a life organic, at maximal volume according to local climatic factors.
Текст научной статьи Продукционный процесс как отражение функционирования целостной экосистемы
где все виды растений выполняют общую функцию по созданию нового органического вещества – чистой продукции экосистемы.
Цель нашей работы – рассмотреть процесс производства продукции в изолированной степной экосистеме с позиций функционирования растительного единства. В частности, изучить ритмику прироста надземной продукции растительным сообществом экосистемы под влиянием гидротермических условий конкретных вегетационных сезонов.
МАТЕРИАЛ И МЕТОДЫ
В основу представлений о функционировании целостной экосистем положен принцип «экосистемного круговорота» или метаболизма экосистем [7]. Он заключается в циклической смене синтеза органического вещества из минеральных элементов автотрофами с помощью солнечной энергии и деструкции отработавшей ресурс и отмершей биомассы гетеротрофами, с возвратом минеральных элементов в новый цикл фотосинтеза. Таким образом, экосистема представляется практически замкнутой системой, лишь малая часть органики которой выходит за пределы биологического круговорота, попадая в большой геологический круговорот [2, 5]. Концептуально модель экосистемы можно представить как круговорот вещества, находящегося в экосистеме в различных состояниях и позволяющих воспроизводить новый цикл синтеза (рис. 1). По качественному состоянию вещество в экосистеме можно представить как «живое вещество» (BM – биомасса), «разлагающееся вещество» (NM – некромасса), «минерализованное вещество» (MM – минеральная масса). Переход вещества из одного состояния в другое регулируют физиологические процессы: анаболизм (ANB, ассимиляция органического вещества из неорганического зелеными растениями в процессе фотосинтеза), некроболизм (NKB, постепенное отмирание и передача части органики в жизненно важные части системы) и катаболизм (KTB, деструкция сложных органических молекул до минеральных компонентов для нового цикла синтеза). Причем, любой из этих процессов имеет двойственную природу: биосинтез (bsn) все- гда подразумевает частичный отток образованной органики, например на дыхание, а деструкция или минерализация (min) всегда сопровождается промежуточным синтезом, например, гуминовых веществ в почве (gum). Каждый из магистральных потоков вещества в свою очередь связан процессами синтеза и распада органического вещества внутри потока.
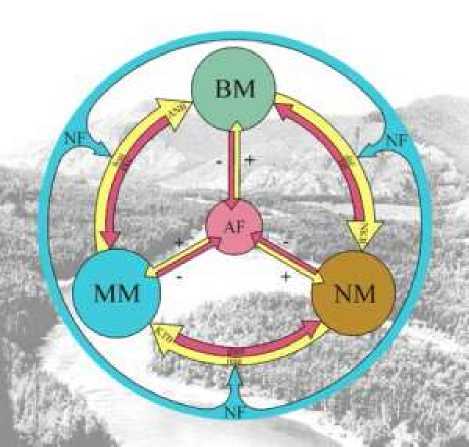
Рис. 1. Концептуальная модель экосистемы (по [5])
В процессе эволюции подбирается такое сообщество живых организмов, которое позволяет автотрофам синтезировать максимально возможное количество органического вещества в данных условиях, а гетеротрофам более бережно сохранять его в системе. Почва является тем механизмом, который накапливает, разлагает и дозирует органику так, чтобы экосистема (биогеоценоз) могла существовать почти автономно, т.е. за счет собственных ресурсов. Цикличность круговорота вещества между фитоценозом и педоценозом обеспечивает устойчивость экосистемы как относительно автономного образования. Тип растительного сообщества (автотрофный компонент экосистемы) складывался в результате длительной эволюции «подогнанных к совместному существованию» видов, максимально полезно преобразующих солнечную энергию в энергию химических связей органического (живого) вещества при данных условиях местообитания [8]. В этом смысле, все виды растительного сообщества предстают не как конкуренты за ресурсы, а как функционально связанные компоненты экосистемы по производству продукции. Отмершее органическое вещество – опад; его количество, скорость и полнота преобразования микробной биотой определяют формирование почвенного профиля, складывающегося в той или иной экосистеме. Таким образом, процесс производства продукции в экосистеме представляется согласованной работой сообщества видов по максимизации метаболизма.
Объектом исследования явилась лугово-степная экосистема урочища Долы, Приокско-Террасный биосферный заповедник (ПТБЗ). Она расположена изолированно от основного ареала, на северозападном пределе произрастания степей и функционирует в широком диапазоне факторов среды. Травяные сообщества Долов идентичны разнотравно-злаковой луговой степи, эталоном которой является Стрелецкая степь Центрально-Черноземного заповедника [4]. Однако крайнее местоположение степной экосистемы ПТБЗ приводит к значительной адаптационной изменчивости основных характеристик, что позволяет рассматривать ее как природную модель экосистемы. Доминирующее положение в сообществе занимают плотнодерновинные злаки. Содоминантами выступают бобовые и виды разнотравья. Для многолетних наблюдений выделены три площадки по 100 м2 каждая, расположенные вдоль градиента влажности: мезоксерофитное сообщество с доминированием Stipa pennata , ксе-ромезофитное – с доминированием Festuca valesiaca , гигромезофитное – с доминированием Phleum phleoides . В течение вегетационного сезона проводятся детальные геоботанические описания и учет надземной продукции по каждому из трех фитоценозов. Измерение годовой продукции проводят по стандартной методике. Статистическая обработка данных проводится при помощи программы MXL. Метеорологические наблюдения ведутся станцией комплексного фонового мониторинга заповедника (СФМ). Общая выборка метеорологических показателей составляет более 35 лет, продуктивности – более 30 лет. Непрерывные ряды наблюдений, полученные нами за последнее 14-летие и архивные данные с 1975 по 1985 гг., укладываются в синхронизацию 11-летних циклов, что позволяет нивелировать различия, связанные с естественными циклами солнечной активности. Сравнение этих двух циклов показательно, т.к. последний цикл совпал с глобальным потеплением, а контрольный цикл попадает в диапазон, оцениваемый климатологами как «норма».
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Многолетняя динамика показателей продуктивности отражает широкую амплитуду флуктуационных изменений лугово-степной экосистемы ПТБЗ. Полный диапазон продуктивности данной экосистемы составил 85,0±5.5–595,0±7,5 г/м2/год, с 7-кратным превышением максимума над минимумом. При этом, в теплом цикле средняя продуктивность составила 332,0 г/м2/год, в холодном – 209,0 г/м2/год. Надземная фитомасса всех годов теплого цикла превышает годовую массу холодного цикла в 1,5-2 раза. В рекордный сезон (2001) отмеченный фактором непромерзания почвы, надземная продукция превысила среднемноголетнюю в 2,2 раза, а продукцию холодного цикла – в 2,8 раз.
Известно, что на продуктивность экосистемы влияют обеспеченность питательными элементами и влагой, солнечный свет и тепло. Традиционно годичную продукцию степных сообществ увязывают с усредненными условиями температуры и влажности каждого сезона [1]. Опыт исследований в ПТБЗ показал, что продуктивность экосистемы не только изменяется в широком диапазоне, но далеко не всегда конечная продукция соответствует средним показателям условий сезона (корреляция не превышает 80%). Так, например, в наиболее благоприятном по гидротермическим условиям 1977 г. надземная фитомасса мезофитного сообщества (Festuca vale-siaca – разнотравье) составила 229,2 г/м2, столько же, как в холодном, но избыточно влажном 1976 году (299,4 г/м2). 1980, 1985 и 1993 годы характеризовались близкими значениями гидротермических условий, а фитомасса была очень разной – от 329,8 г/м2 в 1980-м, до вдвое меньшей (154,6 г/м2) в 1985-м. В сухом и жарком 1999 г. фитомасса оказалась вдвое больше (168,9 г/м2), чем в сухом и жарком 1979 г. (83,5 г/м2).
Чтобы понять механизм прироста надземной продукции, мы исследовали подекадную ритмику прироста, которая совпадает с основными фенологическими изменениями в экосистеме. Учет подекадного прироста продукции (ритмики прироста) проводили на мезофитном стационаре, где условия наиболее типичны для региона. Сезон 2005 г. выбран как наиболее близкий к среднемноголетним климатическим условиям.
Было исследовано влияние температуры и влажности на подекадный прирост надземной фитомассы Долов. Исследования показали, что влияние температуры на прирост продукции в сообществе повторяет ту же закономерность (рис. 2, ряд 1), что и влияние температуры на скорость фотосинтеза отдельного растения [9]. На начальном этапе (в экосистеме – этап фенологической весны) продукция возрастает линейно, как и в случае с фотосинтезом, когда процесс прямо лимитируется фактором внешней среды. В данном случае, прирост фитомассы лимитируется температурой, так же, как нарастание фотосинтеза лимитируется на начальной стадии температурой (или освещенностью). На следующих этапах прирост фитомассы происходит более плавно, т.к. появляются точки частичного оттока продукции: при фотосинтезе – на дыхание и запас; в экосистеме – на «переключение» фенофаз (цветение, созревание семян). Пик прироста зеленой массы приходится в данной экосистеме на последнюю декаду июня. После этого наступает период насыщения, как и в реакции фотосинтеза на температуру (или освещенность). В степной экосистеме период насыщения (с первой декады июля до второй декады августа) прирост фитомассы незначителен. Со второй декады августа, как правило, происходит усыхание растений.
В критических точках темпы прироста снижаются, поскольку часть первичной продукции идет не на рост вегетирующих органов, а на более энергоемкие, но мало заметные в массе вещества (фито- гормоны и запасающие вещества). В отдельные годы в фазу перехода к цветению фитомасса вегетирующих органов может даже снижаться по сравнению с предыдущей декадой. Так происходит, когда сильная засуха приводит к замедлению поставки минеральных элементов из почвы; тогда процесс переключения фенофаз вызывает частичный отток органического вещества из листьев. В период насыщения вся новая органика идет на созревание семян, так что даже при благоприятных условиях для прироста, заметного увеличения зеленой массы не происходит. Возможно поэтому, многие степные виды имеют крупные сочные прикорневые листья (которые к середине вегетации усыхают) и небольшие стеблевые листья (которые вполне справляются с функцией поддержания фотосинтеза при созревании семян).
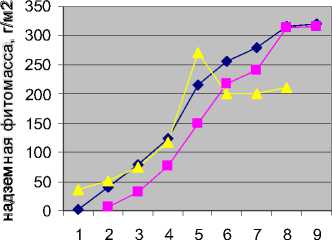
♦ Ряд1
■ Ряд2
Ряд3
дек ады
Рис. 2. Ритмика прироста продукции в мезофитной ассоциации Долов в различные вегетационные сезоны. Ряд 1 – 2005 г., ряд 2 – 2006 г., ряд 3 – 2007 г.
Было изучено влияние гидротермических показателей на каждый этап онтогенеза в различные по благоприятности для роста растений сезоны: 2005 (близкий к норме), 2006 (прохладный и влажный), 2007 (жаркий и сухой). Оказалось, что и ритмика прироста фитомассы и конечная продукция экосистемы в эти годы существенно различаются (рис. 2). Для одних сезонов характерен ранний и резкий старт вегетации (2007), по сравнению с «нормой» (2005); для других (2006) – ее запаздывание. На характере кривой сказывается как переключение фенофаз, так и длительные засухи. При ранжировании сезонов (сумма баллов) по их благоприятности для нарастания фитомассы учитывались следующие параметры: а) гидротермические условия всего вегетационного сезона, б) гидротермические условия наиболее критического периода, в) условия, предшествующие вегетации.
В среднем, надземная продукция в эталонных луговых степях достигает 330 г/м2/год [1]. В нашем регионе прошедшее 15-летие отмечено как самое теплое за всю историю наблюдений. Оно характеризуется увеличением средней годовой продукции лугово-степных сообществ Долов до эталонных значений, что почти на 25,0% больше по сравнению со среднемноголетними данными и более чем на 45,0% больше по сравнению с контрольным 11-летием. Такой рост продуктивности можно объяснить выраженным потеплением климата и более ранним наступлением вегетации растений (что компенсирует для экосистемы влияние весенних засух). Сравнительный анализ продуктивности экосистемы и суммарной оценки сезонов показал, что менее половины сезонов оказались близкими по условиям к среднемноголетней норме региона (913 баллов). Надземная фитомасса (среднее по трем стационарам) в эти сезоны достигла значений в 235,0 – 315,0 г/м2 сухого вещества в год (рис. 3).
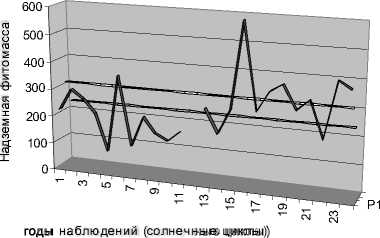
□ Ряд2
□ Ряд3
□ Ряд1
Рис. 3. Динамика продуктивности экосистемы Долов в сравнении с условной нормой.
Ряд 1 – продуктивность холодного и теплого циклов, ряды 2, 3 – интервал условной нормы
Существенные отклонения реальных условий от нормы (5-8 баллов) приводят к значительному падению (на 25-55%) надземной фитомассы. Это отмечалось в каждый четвертый сезон. Наиболее экстремальными оказались два сезона (1-4 балла). Они отличались крайней засушливостью на протяжении всего вегетационного сезона, так что экосистема попала в условия, более характерные для сухих степей. Закономерно, что фитомасса этих сезонов оказалась минимальной для Долов, достигнув лишь трети от условной нормы. Однако экосистема может попасть в условия не только хуже, но и лучше среднемноголетних значений в регионе. Наиболее благоприятными для прироста фитомассы стали пять сезонов теплого цикла (14-18 баллов). Они же дали наибольшую надземную фитомассу, которая в разы превысила условную норму.
ВЫВОДЫ
-
1. Ритмика прироста надземной фитомассы целостного растительного сообщества повторяет закономерности нарастания фотосинтеза отдельного растения.
-
2. Фитоценотический блок экосистемы в своем
-
3. Реальный ход прироста надземной продукции отличается от идеального (среднемноголетнего) и регулируется гидротермическими условиями конкретного вегетационного сезона. Для полной оценки влияния условий на прирост продукции следует учитывать влияние каждого фенологического этапа на конечный результат.
функционировании проходит несколько критических точек (старт вегетации, переключение на цветение, переключение на созревание семян), которые сопровождается уменьшением зеленого прироста за счет создания более энергоемких соединений метаболизма.
Список литературы Продукционный процесс как отражение функционирования целостной экосистемы
- Базилевич Н.И., Титлянова А.А. Биологический круговорот на пяти континентах: азот и зольные элементы в природных наземных экосистемах. Новосибирск: Изд-во СО РАН 2008. 381 с.
- Вернадский В.И. Биосфера и ноосфера. М.: Наука, 1989. 260 с.
- Дылис Н.В. Основы биогеоценологии. М.: Изд-во МГУ, 1978. 172 с.
- Зеленская Н.Н., Керженцев А.С., Аблеева В.А., Терешонок Н.А. Динамика продуктивности луговых степей на северо-западном пределе их ареала (бассейна Оки)//Известия РАН. Сер. Геогр. 2012. № 6. С. 47-54.
- Керженцев А.С. Бесконфликтный переход биосферы в ноосферу -разумный выход из экологического кризиса//Вестник РАН. 2008. Т. 78. № 6. С. 1-8.
- Керженцев А.С. Новое перспективное научное направление//Вестник РАН. 2012. Т. 82. № 5. С. 1-9.
- Ковда В.А., Бугровский В.В., Керженцев А.С., Зеленская Н.Н. Модель трансформации органического вещества в почве для количественного изучения функций почвы в экосистеме//Доклады АН СССР, 1990. 312 (3). С. 759-762.
- Мазинг В.В. Системы биогеоценотического уровня и их усложнение в эволюции//Развитие концепции структурных уровней в биологии. М., 1972. С. 349-356.
- Холл Д., Рао К. Фотосинтез/Пер. с англ. М.: Мир, 1083. 134 с.
