Продуктивность бореальных лесов в островном вулканическом ландшафте Северо-Западной Пацифики

Автор: Коломыц Эрланд Георгиевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-1 т.18, 2016 года.
Бесплатный доступ
Проведено эмпирико-статистическое моделирование лесных сообществ экспериментальном полигона «Вулкан Менделеева» на о-ве Кунашир (Курильская островная дуга). Установлено, что темнохвойные и хвойно-широколиственные островодужные леса отличаются от своих внутриконтинентальных аналогов более резко выраженной стратегией создания максимальной биомассы, с высокой продуктивностью, сближающей их с субтропическими лесами Северо-Западной Пацифики. При этом исключительно большая доля зеленой массы в структуре продукционного процесса ведет к ускорению биологического круговорота, что способствует устойчивости лесных сообществ. Соответственно обеспечивается выживание островных лесов бореального пояса в неблагоприятных для них условиях «холодной» океаничности.
Островные вулканические ландшафты, бореальные леса, фитомассы, продуктивность, биологический круговорот, эмпирико-статистические модели
Короткий адрес: https://sciup.org/148204890
IDR: 148204890 | УДК: 581.52/581.524.3/574.9
Productivity of boreal forests in insular volcanic landscape of North-Western Pacific
The empirical statistical models of the island-arc stage of continental biosphere development in the North-West Pacific have been created by the example of experimental testing area near the active Mendeleev Volcano on the Kunashir Island (the Kuril Insular Ridge). An exceptionally important backbone role of forest phytobiota in the establishment and development of young volcanic landscapes was demonstrated. The optimizing strategy program for the development of plant communities, which is aimed at the maximum biomass formation on forest island-arc landscapes, is much more marked compared to their zonal analogs on the neighboring continent. At the same time, an exceptionally high percentage of green mass in the structure of production results in the acceleration of nutrient cycle cycle as a factor of stability of forest community. All the above ensures the survival of insular ecosystems of the boreal Neo-Pacific under unfavorable conditions of «cold» oceanicity.
Текст научной статьи Продуктивность бореальных лесов в островном вулканическом ландшафте Северо-Западной Пацифики
К настоящему времени уже накоплен достаточно обширный эмпирический материал, характеризующий закономерности и условия формирования растительного покрова Курильских островов, а также соседних островных и окраинно-материковых территорий Тихоокеанского мегаэкотона Северной Евразии. Наряду с многочисленными описаниями даются общие аналитические оценки, направленные на познание глубинных процессов флоро-, фитогенеза в Северо-Западной Пацифике [6, 11, 20, 35, 36, 39, 40 и др.ъ. Гораздо менее изученной остается функциональная организация лесных сообществ островов в различных зональных условиях, в том числе продуктивность бореальных лесов. Отметим две крупные обобщающие сводки [3, 5], где приведены преимущественно зарубежные данные по продуктивности лесов севе-рного бореального пояса Тихоокеанского кольца. Однако эти материалы характери-зуют лишь самые крупные зонально-региональные черты функционирования лесного покрова островодужных ландшафтов и не дают достаточно глубоких представлений о его структурно-функциональных связях. Последние могут быть вскрыты лишь на локальном (топологическом) уровне, где биогеоценозы как элементарные хорологические единицы биосферы составляют ее функциональное «ядро» и образуют истоки планетарных вещественноэнергетических круговоротов [29, 32].
В настоящем сообщении представлен опыт изучения продуктивности бореальных лесных
сообществ топологического уровня в островодужном секторе Азиатско-Тихоокеанского мегаэкотона. Анализ проведен на примере стратовулкана Менделеева (абс. высота центрального конуса 850 м, рис. 1), расположенного в южной части о-ва Кунашир Курильской островной дуги и находящегося в состоянии пассивного вулканизма (активной фумарольной деятельности). Здесь на экспериментальном полигоне Менделеевского ландшафта в первой половине августа 1985 г. нами была проведена крупномасштабная ландшафтноэкологическая съемка по методике, изложенной в (Коломыц, 2008), со сбором более чем 50 параметров структурно-функционального состояния лесных биогеоценозов на 32 пробных площадях.
ОБЪЕКТЫ
По последнему ботанико-географическому районированию [2] о-в Кунашир включен в Южно-Курильско-Хоккайдский округ и в ЮжноКурильский район, с господством темнохвойношироколиственных лесов, обогащенных южными восточноазиатскими элементами ( Magnolia hypoleuca, Quercus crispula, Q. Dentate, Acer tschonoskii, A. japonicum, Betula maximowicziana, Daphniphyllum humile ). В районе вулкана сосредоточены весьма разнообразные во флористическом и ботанико-географическом отношениях группировки растительности [21].
На экспериментальном полигоне нами выделены следующие группы типов леса:
-
1 ) хвойно-широколиственные леса (ХШ – ); 2) темнохвойные (елово-пихтовые) леса
(ТХв – ); 3) каменноберезово-еловые леса (КбЕ – ); 4) каменноберезовое криволесье с кедровым стлаником (КбКс – ); 5) кедро-
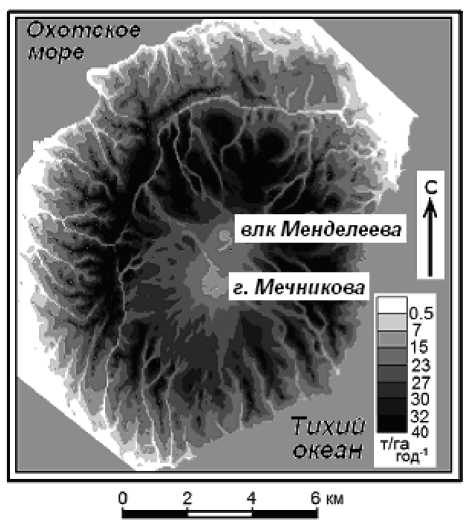
Рис. 1. Растровая карта общей продуктивности хвойно-широколиственных и темнохвойных лесов, каменноберезовых криволесий и кедровых стлаников Менделеевского вулканического ландшафта.
Рассчитана и составлена Л.С. Шарой вый стланик, заросли (Кс – ); 6) смешанные сообщества (каменная береза, ель, кедровый стланик) (БЕКс – ). Эти типологические единицы слагают достаточно хорошо выраженную высотную поясность растительности, несмотря на низкогорный характер вулканогенного рельефа. Самый нижний биоклиматический пояс вулкана Менделеева и его окружения (до 200–250 м. абс.) фрагментарно образуют хвойно-широколиственные леса. Темнохвойные пихтово-еловые ассоциации занимают большую часть площади о-ва Кунашир, в том числе Менделеевского ландшафта. На вулкане они создают следующий высотный пояс (до отметок 580–620 м), а в предгорьях нередко выходят к побережью, главным образом в тихоокеанском секторе ландшафта. Верхний лесной пояс представлен каменно-березовым криволесьем. Выше 670–700 м он сменяется подгольцовым поясом кедрового стланика.
Типы рельефа определяют структуру водных и литодинамических потоков – рассеивающих, транзитных и собирающих. Интегральным выражением этой морфолитодинамики могут служить фоновые типы мезоместоположений . Благодаря достаточно простой геометрической конструкции самого вулканического конуса, описанной высотной поясности соответствуют четыре типа мезоместоположений, по систематике [13]: аккумулятивный (А), трансаккумулятивный (ТА), транзитно–трансэлювиальный (Т–ТЭ) и трансэлювиальный (ТЭ).
Фактор макроэкспозиции склонов в данном эко -регионе отображает известную биоклиматическую асимметрию островодужных систем Северо-Западной Пацифики, обусловленную влиянием теплых и холодных морских течений и соответствующими свойствами воздушных масс, приходящих с Тихого океана и Охотского моря [12, 14]. В соответствии с региональными климатическими контрастами, обусловленными циркуляционной экспозицией макросклонов вулкана Менделеева, нами проведено разделение этого ландшафта на два сектора – охото-морский (Ох, ЮЗ–З–СЗ–С экспозиции) и тихоокеанский (Т/о, СВ–В–ЮВ–Ю экспозиции). Первый сектор является более теплый и менее влажный, второй отличается повышенной «холодной» океаничностью.
МЕТОДЫ
В настоящем анализе использованы следующие функциональные параметры лесных сообществ, измеренные во время ландшафтноэкологической съемки, либо дополнительно рассчитанные по полученным эмпирическим данным: 1) BL – общая живая надземная фитомасса, т/га; 2) PD к – продукция скелетной массы деревьев и кустарников, т/га . год; 3) PV д-к – продукция зеленой массы деревьев и кустарников; 4) PG – продукция зеленой массы напочвенного покрова; 5) PV – общая продукция зеленой массы леса; 6) РС – общая продукция лесного фитоценоза (надземная + подземная).
По данным [3] были рассчитаны зависимости параметров продуктивности ( PD+РV д-к) и РС от запасов живой надземной фитомассы ВL . Использованы 16 пунктов из районов о. Хоккайдо, Приморья, Приохотья и Камчатки. Они характеризуют достаточно разнообразные лесные формации Северо-Западной Пацифики и Субпацифики: елово-пихтовые, лиственничные и каменноберезовые бореальные, пихтовые субальпийские, кедрово-стланиковые субполярные, липово-кленовые с пихтой и дубово-кедровые суббореальные. Получены следующие линейные уравнения связей, с достаточно высокими коэффициентами корреляции (R):
( PD+РV д-к) = 0.953 + 0.027 . ВL ; R = 0.732;
РС = 2.д81 + 0.0455 . ВL ; R = 0.800.
С помощью новых методов геоморфоме-трической статистики [37, 41] Л.С. Шарой была рассчитана и сформирована карта-матрица общей продуктивности ( РС ) лесов Менделеевского вулканического ландшафта (см. рис. 1). В расчетную формулу в качестве значимых предикторов вошли: 1) относительная высота пробной площади; 2) минимальные расстояния от вулкана Менделеева и г. Мечникова; 3) площадь водосбора, замыкающегося на данную пробную площадь. Расчетная модель имеет коэффициент детерминации R2 = 0,78.
Основные закономерности функционирования лесных экосистем Менделеевского вулканического ландшафта охарактеризованы по таксономическим (средневзвешенным) нормам метаболических параметров, а также по их моно-системным (теоретико-информационным) и полисистемным (теоретико-множественным) моделям. Методика построения таких моделей изложена в [1,16, 26]. Получены нормированные коэффициенты сопряженности К(А;В) явления А с фактором В (в каждой паре признаков) и графические модели частных коэффициентов связи C ij групп типов леса с запасами фитомасс и продуктивностью, а также графы мер их отношений сходства (изоморфизма) N ij и орграфы мер отношений включения J ij , по этим функциональным характеристикам лесов. На основе мер отношений включения построены структурные схемы доминирования рассматриваемых лесных образований.
Исходя из этих схем, путем дополнительных расчетов [18] получены два интегральных параметра полисистемного экологического пространства лесов: 1) метаболические индексы лесных сообществ S mk , по которым выделены их экотипы, согласно [8, 24], которые характеризуют общий уровень их функциональной организации; 2) показатель значимости G k данной группы типов леса в отображении высотно-поясного вулканогенного геопространства и приобретения ими соответствующего запаса гомеостатичности.
Аналогичным образом построены теоретико-множественные модели комплексных показателей функционирования лесных экосистем, характеризующие созидательную и частично детритную ветви малого биологического круговорота. Такими показателями являются: 1) коэффициент оборота надземной фитомассы KR = PV/ BL [24]; 2) производный от него аллометрический коэффициент KA = logPV/ logBL [25]; 3) подстилочно-опадный индексKY=ML/PV [4]; здесь Ml - масса лесной подстилки. Параметр KR является показателем эффективности автотрофного биогенеза [13,24], своего рода КПД лесного фитоценоза. Аллометрический коэффициент как инвариантный параметр характеризует степень однородности региональных ландшафтов и служит наиболее объективным критерием выделения их коренных зональных типов [25], находящихся в климаксовом состоянии.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Общая характеристика продуктивности и биологического круговорота
Менделеевский вулканический ландшафт, равно как и о. Кунашир и вся Южно-Курильская гряда, входит в Тихоокеанское окраинно-материковое и островодужное кольцо, отличающееся от внутриконтинентальных территорий в целом более высокими показателями биологической продуктивности и накопления живого органического вещества [3, 5]. Полученные нами эмпирические данные согласуются с этими представлениями относительно бореальных и суббореальных лесов (табл. 1 и 2; рис. 2). Хвойно-широколиственные
Таблица 1. Таксономические нормы фитомасс, продуктивности и комплексных параметров биологического круговорота (КПБК) в группах типов леса Менделеевского вулканического ландшафта
|
Фитомассы, проктивность и КПБК |
Группы типов леса (см. в тексте) |
|||||
|
ШХ М |
ТХв |
КбЕ М |
БКс ^ |
Кс W |
КсБЕ
|
|
|
BL |
340.70 |
223.75 |
163.60 |
89.59 |
89.70 |
135.00 |
|
PG |
11.16 |
14..95 |
11.75 |
8.62 |
4.18 |
14.19 |
|
PD к |
9.52 |
8.14 |
5.72 |
3.81 |
3.16 |
3.86 |
|
PV д-к |
3.99 |
3.21 |
2.41 |
1.83 |
1.62 |
1.94 |
|
( PD+V ) д-к |
13..51 |
11..36 |
8.13 |
5.64 |
4.78 |
5.00 |
|
PV |
15.15 |
18.16 |
14.16 |
10.45 |
5.80 |
16.13 |
|
PC |
24.67 |
26..30 |
19..88 |
14.26 |
8.96 |
20.00 |
|
KR |
0.044 |
0.036 |
0.087 |
0.117 |
0.065 |
0.119 |
|
KA |
0.467 |
0.536 |
0.520 |
0.523 |
0.488 |
0.566 |
|
KY |
1.68 |
0.54 |
1.31 |
0.46 |
5.95 |
0.46 |
Таблица. 2. Таксономические нормы запасов фитомасс, продуктивности и комплексных параметров биологического круговорота (КПБК) в различных высотных ярусах и секторах Менделеевского вулканического ландшафта
|
Фитомассы, продуктивность, и КПБК |
Высотные ярусы и сектора (интервалы абс. высот, м) |
||||||
|
Предгорный - 1 |
Нижний -2 |
Средний - 3 |
Верхний - 4 (> 690– 720) |
||||
|
Ох-1 (60–250) |
Т/о-1 (60–180) |
Ох-2 (250–620) |
Т/о-2 (180–520) |
Ох-3 (620–720) |
Т/о-3 (520–90) |
||
|
BL |
355.58 |
228.5 |
230.8 |
217.9 |
114.2 |
103.4 |
89.6 |
|
PG |
10.30 |
18.36 |
13.55 |
10.95 |
9.42 |
12.90 |
4.18 |
|
PD д-к |
16.25 |
15.44 |
14.90 |
12.20 |
8.38 |
7.83 |
6.50 |
|
PV д-к |
3.91 |
3.90 |
3.36 |
2.98 |
2.05 |
1.80 |
1.62 |
|
PD+ V Д-к |
20.38 |
19.34 |
23.66 |
15.18 |
10.43 |
9.63 |
8.12 |
|
PV |
14.21 |
22.26 |
16.91 |
13.93 |
11.47 |
14.70 |
5.80 |
|
PC |
30.46 |
37.70 |
31.81 |
26.13 |
19.85 |
22.53 |
12.1 |
|
KR |
0.040 |
0.097 |
0.073 |
0.064 |
0.100 |
0.142 |
0.06 |
|
KA |
0.452 |
0.572 |
0.520 |
0.489 |
0.516 |
0.580 |
0.39 |
|
KY |
2.08 |
0.21 |
0.72 |
0.89 |
1.49 |
0.48 |
5.95 |
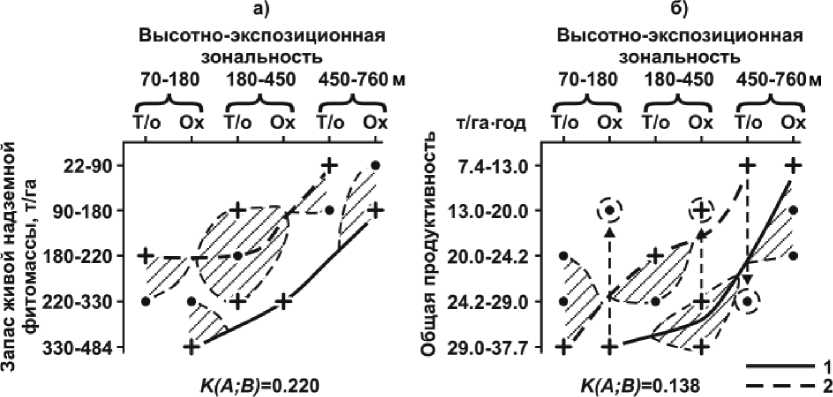
Рис. 2. Распределение запасов фитомасс и продуктивности лесов по трем ступеням абс. высот в охотоморском (Ох – 1) и тихоокеанском (Т/о – 2) секторах Менделеевского ландшафта:
1 – экологический доминант; 2 – «размытая» часть экологической ниши; 3 – траектория, соединяющая экологические доминанты; 4 – пространство экологической ниши; 5 – анклав; 6 – высотный интервал дробления экологических доминантов; 7 – область спорадического распространения явления в пределах данных градаций фактора
леса низменных предгорий Менделеевского ландшафта обладают такими запасами надземной живой фитомассы (таксономическая норма BL ≥ 340 т/га), которые считаются максимальными для Северо-Западной Пацифики (около 300 т/га)
и даже превышают данный максимум (табл. 2). Для сравнения отметим, что в наиболее продуктивных суббореальных (сосново-дубово-липовых) лесах Приокско-Террасного заповедника, находящегося на юге подтаежной зоны Русской равнины, таксономическая норма BL = 376 т/га, а в кедрово-широколиственных лесах бореального экотона Нижнего Приамурья она едва достигает 240 т/га [17]. Пихто-ельники нижнего высотного яруса (250–500 м) по параметру BL = 215–230 т/ га не уступают темнохвойным лесам о. Хоккайдо, Саха лина и Приморья, где значения BL колеблются от 130–150 до 230–280 т/га [3].
Еще более значительно превышение по годичному производству фитобиоты. Подчеркнем в целом феноменально высокую продуктивность южнокунаширских хвойно-широколиственных и пихтово-еловых лесов . Таксономическая норма РС > 24–26 т/га . год (см. табл. 1), с максимальными значениями, достигающими 35–40 т/га . год (см. рис. 2). Эта норма превышает производительность умеренных хвойных и смешанных лесов Приморья, Маньчжурии и Северо-Восточного Китая, где величина общей продукции находится в пределах 11–18 т/га . год, редко достигая 21–24 т/га . год. В южной тайге Западной и Восточной Сибири, она составляют от 2,5 до 7,1 т/га . год [5]. В хвойно-широколиственных лесах Нижнего Приамурья средние значения РС =8,5–9 т/га в год, а на бореальном экотоне Волжского бассейна – около 11–12 т/га . год. Близкие к Южному Кунаширу значения продуктивности (в среднем от 7–10 до 20–25 т/га . год) имеют только широколиственные и хвойные леса о-вов Хонсю и Хоккайдо [3].
Таким образом, «запрограммированность» стратегии развития растительных сообществ на создание максимальной биомассы [30, с. 47] в лесных ландшафтах островной Неопацифики выражена гораздо сильнее, нежели в их зональных аналогах на соседнем материке. По особенностям структуры и функционирования наземные экосистемы Тихоокеанского кольца представляют «… особый класс совокупностей природных образований « [3, с. 208].
Реализация принципа максимальной продуктивности, т.е. достижения наибольшего КПД фитоценоза, возможна лишь при благоприятных условиях абиотической среды. Уже сам этот факт противоречит расхожему представлению о том, что зональные типы растительных формаций на островах Северо-Западной Пацифики существуют при весьма ограниченных теплоэнергетических ресурсах. Далее, для достижения максимальной продуктивности в растительном сообществе необходимо гармоничное сочетание функций фотосинтеза и дыхания [34]. Судя по приведенным материалам по продуктивности, правомерно допустить наличие такой достаточно оптимальной взаимосвязи в функционировании лесных сообществ Менделеевского вулканического ландшафта. Соответственно можно полагать достаточно высокую степень адаптации этих наземных фитоценозов к условиям экстраординарной океанической среды. Это обеспечивает их выживание, а также повышает способность к эволюционным преобразованием. Как известно [33], адаптация лежит в основе всех форм эволюции экосистем, и данный механизм неизменно срабатывал в критические эпохи истории биосферы [10].
Выделяются две основные черты распределения в Менделеевском ландшафте запасов живой надземной фитомассы и продуктивности лесов (см. рис. 1): 1) общее снижение этих показателей с абсолютной высотой и 2) доминирование охотоморского сектора над тихоокеанским по высотно-поясным значениям BL и PC . Если первое явление логично объясняется биоклима-тической высотной поясностью, то причинные механизмы второго, по-видимому, более сложны. Здесь проявляются как циркуляционно-экспозиционная зональность, наиболее благоприятная для фитобиоты охотоморского сектора, так и преобладающее в этом же секторе воздействие вулканических геотерм на эдафотопы.
Суббореальные и бореальные леса молодого вулканогенного ландшафта Неопацифики резко отличаются от своих зональных внутриконтинен-тальных аналогов не только общими величинами фитомасс и продуктивности, но и самой структурой продукционного процесса (рис. 3). Особенно высока на вулканических островах продукция зеленой массы напочвенного покрова ( PG ), где значительную долю составляют, как известно, круп-нотравье ( Filipendula kamchatica , Senecio cannabifolius , Petasites amplus , Cirsium weyrichii , Angelica ursine и др.) и заросли курильского бамбука ( Sasa kurilensis ) . Для южнокунаширских хвойно-широколиственных лесов продукция зеленой массы травостоя PG в таксономической норме превышает 11,0 т/га . год, с максимумом до 28,0 т/га . год. Это составляет около 44% производимой общей надземной фитомассы, в то время как производство скелетной массы деревьев и кустарников ( PD ) не превышает 40%.
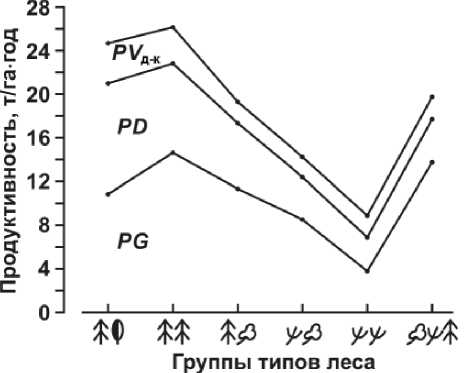
Рис. 3. Распределение продуктивности зеленой массы напочвенного покрова (PG), массы древесины (PD) и зеленой массы деревьев и кустарников (PVд-к) в различных группах типов леса
Еще больший разрыв в данных параметрах (61% против 32%) имеют темнохвойные леса. В целом доля фотосинтезирующей зеленой массы в бореальных и суббореальных лесах Менделеевского ландшафта составляет 60–70% от общей надземной продуктивности.
Для сравнения отметим, что средние и максимальные значения PG в окраинно-материковом Нижнем Приамурье находятся в пределах соответственно 0,5–1,3 и до 3,4 т/га . год, а во внутри-континентальном Приокско-Террасном заповеднике – 5,5–7,5 и до 10,5 т/га . год. При этом резко снижается доля общей зеленой массы, которая составляет в первом случае от 33–36% до 42–47%, а во втором – от 38–40% до 46–49% достигая 61% лишь в сосново-липово-дубовых лесах.
Как видим, островодужном секторе Азиатко-Тихоокеанского мегаэкотона автотрофный биогенез в суббореальных и бореальных лесах направлен главным образом на производство зеленой фотосинтезирующей массы, идущей ежегодно в опад. Это дает толчок существенному ускорению биологического круговорота как фактора функциональной устойчивости лесного биогеоценоза [19], что и подтверждается значе-
Таблица 3. Таксономические нормы комплексных параметров биологического круговорота в лесных сообществах внутриконтинентального и окраинно-материкового экорегионов Северной Евразии
|
Экорегион |
Группы типов леса (типы локальных местоположений) |
Комплексные параметры |
||
|
KR |
KA |
KY |
||
|
s 2 a 0 OJ 0 g С о S ” и а * о м ч а о уда |
Сосновые леса с березой (Э, ТЭ) |
0.027 |
0.437 |
3.7 |
|
Еловые сосновоеловые леса (Э, ТЭ) |
0.021 |
0.422 |
4.8 |
|
|
Сосново-липоводубовые леса |
0.026 |
0.428 |
3.0 |
|
|
Липово-березовые леса (ТЭ, Т) |
0.029 |
0.441 |
3.5 |
|
|
Ельники и сосново-еловые леса (ТА) |
0.018 |
0.403 |
5.2 |
|
|
Л со 2 u S Л Он Ц С о ° |
Широколиственные леса (Э, Saq) |
0.040 |
0.351 |
– |
|
Кедрово-широколиственные леса (Э, ТЭ) |
0.032 |
0.338 |
– |
|
|
Елово-широколиственные леса (А, Saq) |
0.032 |
0.313 |
– |
|
|
Пихтово-еловые леса (Т-ТА, А) |
0.023 |
0.225 |
– |
|
|
Лиственничные леса (Т-ТА, ТЭ) |
0.029 |
0.289 |
– |
|
Примечание. В скобках указаны типы местоположений ниями комплексных параметров метаболизма (см. табл. 1). Суббореальные и бореальные леса Менделеевского островного ландшафта имеют достаточно высокие значения годичного оборота надземной органики (в норме KR = 0,044–0,036). В окраинно-материковом Нижнем Приамурье аналогичные лесные биогеоценозы имеют норму данного параметра 0,023–0,040, а на бореальном экотоне Среднего Поволжья – только 0,017–0,030. По сравнению с Восточноевропейским бореальным экотоном (см. табл. 3) скорость оборачиваемости фитомассы в южнокунаширских хвойно-широколиственных лесах возрастает на 35–40%, а в темнохвойных – на 40–50%.
Весьма показателен также подстилочно-опад-ный индекс KY , служащий, как известно, основным пропускным каналом метаболических процессов в лесной экосистеме. Бореальные и суббореальные леса Менделеевского ландшафта существенно отличаются от своих внутриматериковых аналогов гораздо более оптимальным соотношением прихода и расхода мертвой органики в лесной подстилке, о чем свидетельствуют относительно низкие значения параметра KY (в таксономической норме 0,21–0,44 до 1,68–2,08 лет против 6,42–7,95 лет, например, в Приокско-Террасном заповеднике). Пропускная способность данного канала возрастает в суббореальных лесах в 1,7–2 раза, а в бореальных – в 8–10 раз. Окраинно-материковое Нижнее Приамурье стоит уже гораздо ближе к Южному Кунаширу по указанным показателям биологического круговорота, хотя здесь он также ослаблен (см. табл. 1 и 3).
Таким образом, экстразональные, т.е. выходящие за рамки внутриматериковых зональных норм, показатели продуктивности и биологического круговорота в суббореальных и бореальных лесах молодых островодужных ландшафтов свидетельствуют о высокоэффективной работе их почвенно-биотического ядра . Интенсификация метаболизма направлена на обеспечение функциональной устойчивости и структурного развития лесных сообществ в условиях крайне неблагоприятной для лесообразования холодной в целом океаничности климата Северо-Западной Пацифи-ки. Это позволяет живой надземной фитомассе достичь значений, приближающихся к максимуму в западных внутриконтинентальных экорегионах. Такова, по-видимому, одна из важнейших закономерностей региональной функциональной организации лесного покрова в островодужноом секторе Азиатско-Тихоокеанского мегаэкотона.
Высотно-поясная структура функционального фитоценотического пространства
Наиболее общей чертой этого пространства является его высокая дифференцированность, автономность метаболических процессов в раз- личных группах типов леса. Орграфы отношений включения (рис. 4–6) представлены исключительно слабыми связями, при отсутствии связей толерантности, необходимых для формирования типологического ядра данной системы. Исключительно большое значение критического порога неразличимости (Zкрит = 1) означает весьма высокий нижний предел мер включения элементов системы, при котором еще сохраняется ее структурное, либо функциональное единство [1]. Для сравнения отметим, что в Нижнем Приамурье (окраинно-материковом секторе мегаэкотона) параметр Zкрит = 0,975, а на бореальном экотоне Волжского бассейна он снижается до 0,942–0,937 [16, 17]. Функциональное фитоценологическое пространство молодого островодужного вулканического ландшафта остается еще весьма раздробленным, со слабо выраженными латеральными взаимодействиями между отдельными фрагментами природно-территориальной мозаики. Это свидетельствует о сравнительно низком уровне пространственной организации данной гео(эко-)системы, с отсутствием по существу ее типологического ядра, которое обеспечивало бы ей качественную определенность.
Перейдем к конкретному функциональному анализу. По запасам надземной фитомассы BL предгорно-равнинные хвойно-широколиственные леса резко отличаются от всех других лесных образований Менделеевского ландшафта (рис.
а б
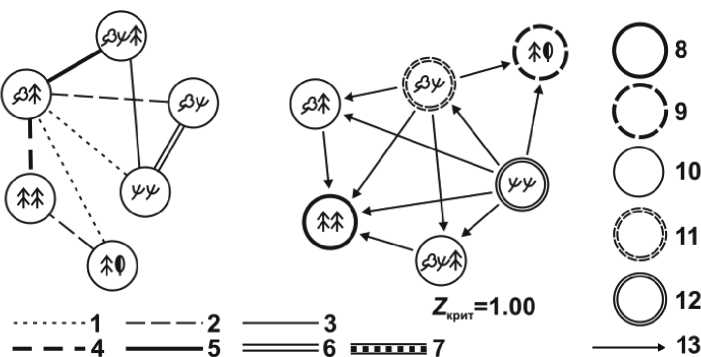
Рис. 4. Схемы нетранзитивных отношений между группами типов леса Менделеевского вулканического ландшафта по запасам фитомасс: граф отношений сходства (а) и орграф отношений доминирования (б). Меры сходства: 1 – 0,66–0,70; 2 – 0,71–0,75; 3 – 0,75–0,80; 4 – 0,81–0,85;
5 – 0,86–0,90; 6 – 0,91–0,95; 7 – 0,96–1,00.
Экотипы лесных сообществ: 8 – макроболиты; 9 – субмакроболиты; 10 – мезоболиты; 11 – субмикроболиты; 12 – микроболиты; 13 – направление включения.
Zкрит – критический порог неразличимости объектов
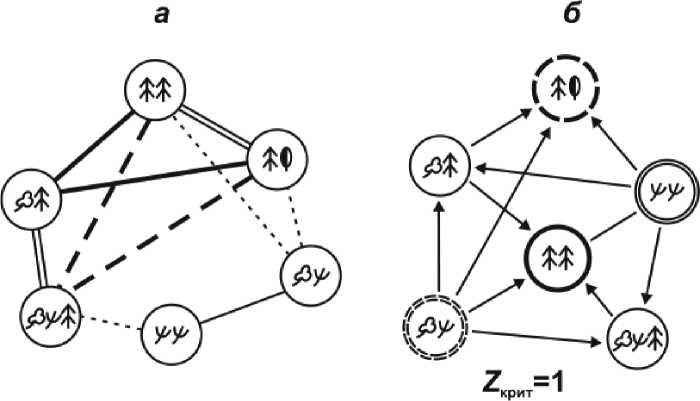
Рис. 5. Схемы нетранзитивных отношений между группами типов леса Менделеевского вулканического ландшафта п о годично й продуктивности: граф отношений сходства (а) и орграф отношений доминирования (б).
Условные обозначения те же, что и на рис. 4
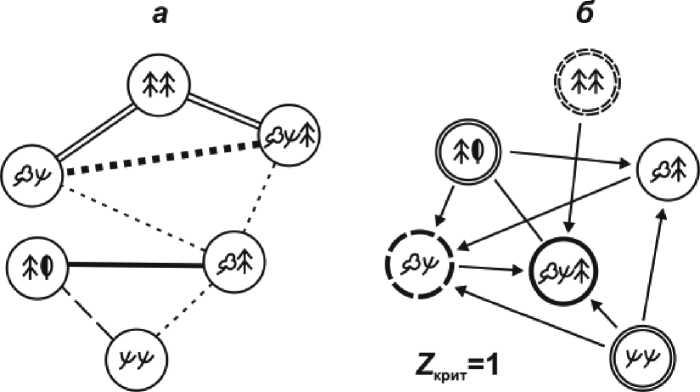
Рис. 6. Схемы нетранзитивных отношений между группами типов леса Менделеевского вулканического ландшафта по биологическому круговороту: граф отношений сходства (а) и орграф отношений доминирования (б).
Условные обозначения те же, что и на рис. 4
4 а ). Среднее значение их мер сходства Nik = 0,58. Характерно также их слабо связное положение субмакроболитов в структурных схемах доминирования не только по фитомассе, но и по общей продуктивности (рис. 5 б , 6 б ), что свидетельствует об определенной уникальности этих суббореальных лесных образований, их функциональной обособленности от общего высотно-зонального биоклиматического пространства Менделеевского ландшафта. Хотя мера сходства хвойношироколиственных лесов с соседними пихто-ельниками по BL и PC достигает соответственно 0,80 и 0,94, их взаимные отношения включения отсутствуют, т.е. они не выстраиваются в соподчиненный ряд автотрофного биогенеза.
При подъеме по склонам вулканического купола в соответствии со сменой климатических условий происходит общее снижение BL и РС , а также парциальных характеристик продуктивности: скелетной древесно-кустарниковой, зеленой общей и травянистого покрова (см. табл. 1 и 2). Тем не менее, по продуктивности бореальные лесные сообщества предгорий, нижнего и среднего ярусов низкогорий вулкана Менделеева имею гораздо больше сходства (рис. 5 а ), нежели по запасам фитомасс.
Уступая существенно по запасам живой надземной органики и незначительно по производству древесно-кустарниковых фитомасс суббореальным лесам (см. табл. 1), темнохвойнолесные сообщества продуцируют наибольшее количество зеленой массы напочвенного покрова и в целом по суммарной продуктивности занимают лидирующее положение макроболитов . Судя по значениям метаболического индекса S mk и меры гомеостатичности G k, они в наибольшей степени отображают высотно-зональное пространство Менделеевского вулканического ландшафта (рис. 4 б ; табл. 4) и их суммарный продукционный процесс обладает максимальной устойчивостью.
В среднем и верхнем ярусах Менделеевского вулканического купола, где на абс. высотах 500–600 м происходит смена темнохвойных пород каменной березой с кедровым стлаником, живая надземная фитомасса имеет примерно такие же средние значения BL (90–110 т/га), что и в предгорном поясе Центральной Камчатки, находящегося у северной границы бореального пояса. Минимальным запасом надземной фитомассы и ее годичным производством отличается подгольцовый пояс кедрового стланика (см. табл. 1 и 2). Исключительно высокая близость по запасам фитомассы каменноберезового криволесья и кедровых стлаников ( Nik = 0,98), а также их существенный отрыв от суббореальных и бореальных лесов по продуктивности (см. рис. 4 а ) свидетельствуют о функциональном единстве этих фитоценотических групп, образующих, как можно полагать, общую охотско-берингийскую биоклиматическую систему подгольцового пояса.
Оби эти фитоценотические группы имеют весьма слабое сходство с лежащими ниже горными пихто-ельниками ( Nik = 0,52–0,58), однако более высокая близость к ним каменноберезо-во-еловых лесов ( Nik = 0,70–0,72) указывает на буферный характер последних, переходный от темнохвойнолесного пояса к субальпийскому. Характерна при этом достаточно тесная связь этих бинарных буферных сообществ по запасам фитомассы с другой более сложной (тернарной) буферной группой – кедровостланиково-березо-во-еловой (см. рис. 4 а ; 5 а ), которая так же формируется у верхней границы темнохвойного пояса.
В отличие от автотрофного биогенеза, который охарактеризован запасами фитомасс и продуктивностью, малый биологический круговорот имеет свои, существенно иные закономерности отношений сходства (изоморфизма) и доминирования лесных фитоценозов. Прежде всего,
Таблица 4. Менделеевский ландшафт. Меры гомеостатичности (Gk ) и структурно-метаболические индексы (Smk) различных групп типов леса
|
Параметры |
Группы типов леса |
||||||
|
м |
|||||||
|
Живые фитомассы |
G k |
0.324 |
0.343 |
0.250 |
0.180 |
0.108 |
0.254 |
|
S m k |
1.18 |
1.36 |
1.08 |
0.74 |
0.55 |
1.09 |
|
|
Продуктивность |
G k |
0.327 |
0.333 |
0.218 |
0.167 |
0.103 |
0.249 |
|
S m k |
1.25 |
1.33 |
1.00 |
0.75 |
0582 |
1.09 |
|
|
Биологический круговорот |
G k |
0.111 |
0.155 |
0.231 |
0.275 |
0.111 |
0.347 |
|
S m k |
0.70 |
0.90 |
1.00 |
1.20 |
0.70 |
1.50 |
|
Примечание. Выделены метаболические экотипы лесных сообществ:
1) макроболиты и субмакроболиты – жирным шрифтом соответственно с подчеркиванием и без него;
2) микроболиты и субмикроболиты – курсивом соответственно с подчеркиванием и без него. Мезоболиты оставлены без выделения. Остальные обозначения см. в тексте
отмечается гораздо более контрастное распределение метаболизма по группам типов леса, о чем свидетельствует сильно разреженный граф мер сходства (рис. 6 а ). Параметр Nik колеблется от 0,41 до 0,99, с основным диапазоном 0,56–0,92. Это говорит о резко выраженной неоднородности механизмов биологического круговорота. При этом хвойно-широколиственные и темнохвойные леса находятся на разных «полюсах» этой контрастности: их Nik = 0,62 – ниже принятого нами уровня минимального сходства, а в структурной схеме доминирования они так же, как и по продуктивности, имеют бинарные отношения ниже порога неразличимости Z крит (рис. 6 б ). Налицо явное расхождение в механизмах метаболизма этих двух фитоценотических групп, составляющих функциональное «ядро» Менделеевского вулканического ландшафта.
Скорость годового оборота надземной живой фитомассы существенно зависит от возраста ( τ ) лесного насаждения. По материалам работ в Нижнем Приамурье получена следующая линейная зависимость KR от τ :
KR = 0,0749 – 0,00037 . τ ; R = 0,765.
С возрастом основных лесообразующих пород скорость годового оборота фитомассы в целом снижается. Следовательно, величина KR характеризует в неявном виде степень зрелости лесного фитоценоза, близость его к климаксово-му состоянию. Исходя из этого, можно считать хвойно-широколиственные леса предгорных равнин Менделеевского ландшафта наиболее возрастными и следовательно максимально приближенными к климаксовому состоянию. В то же время эти суббореальные леса, имея достаточно высокую продуктивность, отличаются минимальными значениями параметров Gk и Smk, т.е. они по существу достигли предела своего функционального развития, уже в минимальной степени отображают высотно-зональное разнообразие биологического круговорота и, таким образом, относятся к наиболее редуцированным экотипам лесных сообществ данного экорегиона – микро-болитам. Соответственно, их биологический круговорот обладает минимальной устойчивостью. К ним же примыкают и наиболее продуктивные предгорные и горные пихто-ельники, составляющие субмикроболитную категорию.
Определенную близость к равнинных суббореальным лесам по биологическому круговороту обнаруживают субальпийские кедровостланиковые сообщества, несмотря на то, что они наименее продуктивны. Это позволяет считать их так же достаточно зрелыми, псевдоклимаксными фитоценозами.
В противоположность сказанному привлекает внимание высокоизоморфная триада горных пихто-ельников с каменноберезовым криволесьем и наиболее сложными буферными кедро-востланиково-березово-еловыми сообществами
( Nik = 0,92). По нашим наблюдениям, тернарная буферная КсБЕ полоса возникла у верхней границы темнохвойнолесного пояса в результате трансгрессии ели и пихты в вышележащие по склону каменные березняки. Об этом свидетельствуют, во-первых, хорошее состояние возобновления и подроста хвойных, а во-вторых, – наличие в подлеске и напочвенном покрове их спутников (тиса, рябины, клена, лиан, папоротников, курильского бамбука). Однако взрослые особи приняли характерные для субальп морфофизиологические черты: общую низкорослость и нередко стланиковую форму, сбежистые и искривленные стволы и ветви, суховершинные и ветровые (плосковершинные) кроны. Присутствует также масса тонкоствольного валежника хвойных. Все это – свидетельства критических климатических условий для развития темнохвойных пород в данных буферных фитоценозах. Налицо пример характерного для Южно-Курильских островов явления снижения бонитета хвойного леса с его возрастом. В то же время для каменной березы эти условия вполне комфортны: она почти не стелится, а дает прямоствольные наклонные жизненные формы.
По биологическому круговороту буферные КсБЕ леса имеют весьма высокий изоморфизм с березовым криволесьем ( Nik = 0,92). Очевидно, приобретя уже определенные структурные черты темнохвойнолесного пояса, эти сообщества еще сохранили функциональные свойства каменных криволесных березняков. Будучи мезоболитами по продуктивности, они выходят в доминанты-макроболиты по скорости биологического круговорота (см. рис. 6 б ). В данном спектре фитоценотических групп эти наиболее сложные буферные образования оказываются, по-видимому, и наиболее молодыми, прогрессивно развивающимися, что придает им, судя по значениям параметров S mk и G k, максимальную функциональную устойчивость. Следуя положению о функциональных критериях эволюции [33], можно полагать, что буферные кедровостла-никово-березово-еловые леса , обладающие наиболее интенсивным (и в этом смысле наиболее совершенным) и устойчивым биологическим круговоротом, стоят на передовом рубеже эволюционного прогресса в лесном покрове Менделеевского вулканического ландшафта.
К субмакроболитам относятся базовые сообщества каменноберезовых криволесий с кедровым стлаником, которые, следовательно, можно считать также сравнительно молодыми и прогрессирующими ассоциациями, с достаточно высоким запасом гомеостатичности, т.е. с устойчивым метаболизмом.
Таким образом, автотрофный биогенез и малый биологический круговорот (в его рассматриваемом нами весьма неполном виде) представляют собой два качественно различных аспекта функционирования островных лесных экосистем, со своими метаболическими параметрами, которые дополняют друг друга, давая более полное представление о сложной системе функциональной организации лесного покрова молодого вулканического ландшафта.
Структура теплового баланса бореальных лесов островодужной Неопацифики
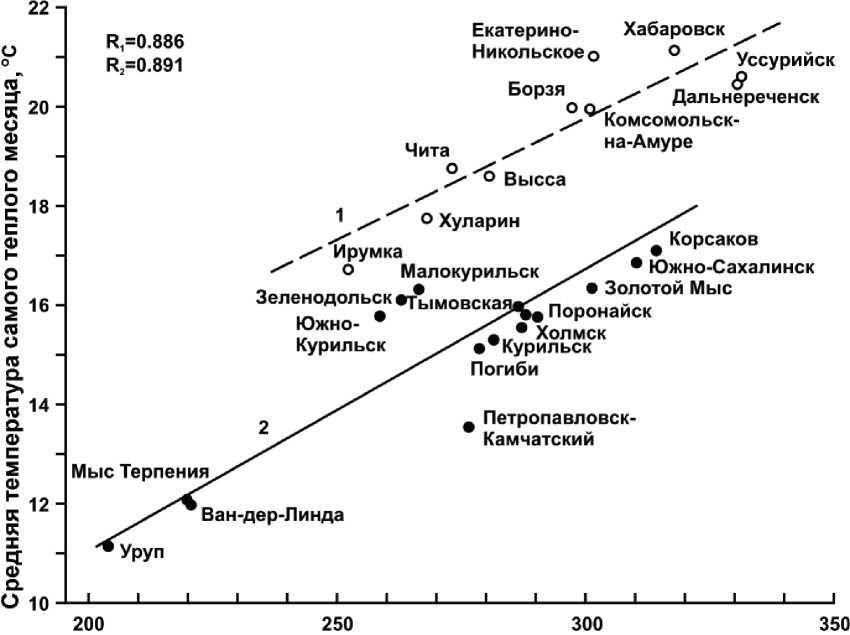
Как известно, одной из характерных географических закономерностей Северо-Западной Неопацифики является общее снижение границ климатических поясов и природных зон на юг по сравнению с соседними материками, с одновременным меридиональным растяжением самих поясов и зон, что объясняется в первую очередь недостатком теплоэнергетических ресурсов [9, 14, 22 и др.]. Приведенные в табл. 5 цифры как будто подтверждают данное положение, однако причины этого кроются в гораздо более сложных ландшафтно-геофизических процессах. Дело в том, что снижение температуры вегетационного периода на островах оказывается значительно большим, чем уменьшение поступающей солнечной радиации и радиационного баланса. Так, по сравнению с южнотаежными и лесостеп-ными районами Забайкалья в на о. Кунашир сумма радиационного баланса за период май–сентябрь снижается на 4–11%, а температура самого теплого месяца – на 16–21%. На Южном Сахалине величина параметра В почти такая же, как и в Среднем Приамурье и значительно выше, чем в Центральном и Южном Забайкалье, тем не менее летняя температура на острове снижается по сравнению с этими регионами соответственно на 4,2о и 3,1о. В целом при одном и том же радиационном балансе вегетационного периода ( В вп) разница в средней температуре самого теплого месяца ( t VII–VIII) между окраинно-материковым (а также внутриконтинентальным) сектором (1) и сектором островным (2) Азиатско-Тихоокеанского мегаэкотона составляет около 3,0–3,5о (рис. 7). Эти различия достаточно четкие и выражаются следующими уравнениями линейной регрессии:
t VII–VIII = 8,14 + 0,039 . В вп; R = 0,886; t VII–VIII = 0,21 + 0,056 . В вп; R = 0, 891.
Подобный контраст в соотношении радиационного и термического параметров обусловлена секторными различиями в структуре расходной части теплового баланса – главным образом в затратах остаточной радиации на эвапотранспирацию и турбулентный теплообмен (последний как раз и определяет температуру приземного слоя воздуха). Как известно, при достаточном и избыточном атмосферном увлажнении суммарное испарение с поверхности сомкнутого растительного покрова представлено главным образом транспирацией [7, 9], которая является необходимым условием первичной биопродуктивности и накопления
Радиационный баланс вегетационного периода, мдж/м2-мес.
Рис. 7. Корреляционные связи средней температуры самого теплого месяца с радиационным балансом вегетационного периода.
Долготные секторы Азиатско-Тихоокеанского мегаэкотона:
1 – окраинно-материковый (Субпацифика – Нижнее Приамурье, Приморье)
и внутриконтинентальный (Палеопацифика – Центральное и Южное Забайкалье, Среднее Приамурье);
-
2 – островной и полуостровной (Неопацифика – Курильские острова, Южный Сахалин, Южная Камчатка). Приведены названия метеостанций
Таблица 5. Сопоставление радиационно-термических параметров внутренних, окраинных и островных территорий Азиатского материка в южной части умеренного пояса
|
Регион (метеостанция) |
Энергетические параметры, МДж/м2 мес. и оС |
||
|
Q |
B |
t VII–VIII |
|
|
Южное Забайкалье (Борзя) |
515 |
297 |
20.0 |
|
Центральное Забайкалье (Чита) |
540 |
268 |
18.8 |
|
Среднее Приамурье (Хабаровск) |
633 |
318 |
21.1 |
|
Сахалин (Южно-Сахалинск) |
451 |
310 |
16.9 |
|
Кунашир (Южно-Курильск) |
450 |
169 |
15.8 |
Примечание. Q и B – соответственно суммарная радиация и радиационный баланс. Эти параметры взяты для горизонтальной поверхности при средних условий облачности; рассчитаны за период май–сентябрь.
tVII–VIII – средняя температура самого теплого месяца (оС)
биомассы [30]. В смешаннолесной и таежной зонах Русской равнины доля транспирации в суммарном испарении лиственных и хвойных лесов составляет 65–70% [27]. На океанических островах бореального пояса эту долю следует, по-видимому, увеличить ввиду высокой влажности воздуха.
Как уже говорилось, островные и прибрежно-морские экосистемы Тихоокеанского кольца отличаются от их зональных аналогов во внутри-континентальных регионах повышенными значениями первичной продуктивности. Отмечены также экстремально высокие значения годичного производства и накопления живой фитомассы в лесных сообществах Менделеевского вулканического ландшафта. Естественно, более высокая продуктивность должна быть обеспечена более значительными затратами тепла на транспирацию. В табл. 6 представлены результаты расчетов таких затрат для двух экорегионов (Нижнего Приамурья и юга о. Кунашир) по сопоставимым лесным равнинно-предгорным сообществам – хвойно-широколиственным и темнохвойным.
При незначительной в общем разнице в радиационном балансе контрасты в затратах тепла на транспирацию между сообществами-аналогами и в целом между самими экорегионами оказались просто поразительными. В Нижнем Приамурье эти затраты на общую продукцию и производство суммарной зеленой массы для смешанных (кедрово- и елово-широколиственных) лесов составляют соответственно 26–28% и 17–19% от радиационного баланса вегетационного периода, а для пихто-ельников снижается до 14% и 7%. В смешанных и хвойных лесах бореального экотона Волжского бассейна величина продуктивной транспирации составляет 260–300 мм [15], с энергетическим эквивалентом 650–750 МДж/м2, что достаточно близко к Нижнему Приамурью (см. табл. 6).
В Менделеевском же островном ландшафте для обеих групп бореальных лесов (смешанных и темнохвойных) транспирационная доля расходов радиационного баланса достигает 71–73% по общей продукции и 52–58% по зеленой массе. Еще более высоки контрасты в затратах тепла на производство зеленой массы напочвенного покрова (33–42% против 1–2%), что и отражает известное явление гигантизма в травостое островных и прибрежно-морских экосистемах Неопацифики. На о-ве Кунашир это заросли бамбучника и крупнотравье, на развитие которых тратится не менее одной трети теплового баланса вегетационного периода.
При таких аномально высоких затратах остаточной радиационной энергии на транспирацию существенно уменьшается доля турбулентного теплообмена земной поверхности с атмосферой, что и вызывает понижение температуры вегетационного периода, с соответствующими изменениями зональных гидротермических параметров в Неопацифическом секторе и с упомянутым выше сдвигом зональных границ на юг. Однако расхожее представление о снижении широтнозональных норм теплоэнергетических ресурсов в Северо-Западной Неопацифике, основанное на температурных показателях, по-видимому, нуждается в корректировке, с учетом структуры расходных статей теплового баланса. По этой структуре внутриконтинентальные и даже окраинно-материковые территории существенно отличаются от островных океанических. В противном случае невозможно объяснить кажущееся парадоксальное явление – аномально высокую продуктивность природных экосистем островной Неопацифики при относительно низкой температуре вегетационного периода, которая выдается за показатель «теплоэнергетических ресурсов». Таким показателем следует считать, несомненно, сам радиационный баланс.
Итак, можно выделить два основных мегатипа структуры теплового баланса наземных экосистем - материковый и океанический ( с возможными промежуточными типами). Эти типы достаточно резко различаются соотноше-
Таблица 6. Затраты радиационной энергии в лесных сообществах окраинно-материкового и островодужного экорегионов на транспирацию в процессе создания массы травостоя, общей зеленой массы и суммарной годичной продукции
|
Энергетические параметры, МДж / м 2 |
Экорегионы (экспериментальные полигоны) и группы типов леса |
|||||
|
Нижнее Приамурье (Комсомольский заповедник) |
О-в Кунашир (Вулкан Менделеева) |
|||||
|
Кедровошироколиственная |
Еловошироколиственная |
Пихтовоеловая |
Темнохвойношироколиственная |
Пихтовоеловая |
||
|
Суммы месячных значений баланса за вегетационный период |
2357 |
2357 |
2422 |
2031 |
2077 |
|
|
S Й Е g н u ПЗ Д ПЗ m m К |
массы травостоя |
34 |
42 |
53 |
672 |
864 |
|
общей зеленой массы |
447 |
404 |
269 |
1184 |
1095 |
|
|
Общей фитомассы |
670 |
610 |
348 |
1440 |
1520 |
|
Примечание. При расчетах использованы данные актинометрических наблюдений на метеостанциях Хабаровск и Южно-Курильск, переводные коэффициенты радиационного баланса при переходе от поля (площадка метеостанции) к лесу [27], а также транспирационные коэффициенты для различных древесных пород [28]
ниями двух основных составляющих расходной части баланса, а именно эвапотранспирации и турбулетного теплообмена. Господство транспирационной составляющей теплового баланса в океаническом типе обусловлено имманентной стратегией развития растительных сообществ островной Неопацифики по принципу максимальной продуктивности (см. выше), что обеспечивает выживание молодых наземных (особенно лесных) экосистем в неблагоприятных условиях экстраокеанического климата на начальном этапе субаэрального биогеогенеза. Оказывая обратное воздействие на расходные статьи теплового баланса, лесные экосистемы уже на данном этапе развития континентальной биосферы оказываются достаточно мощным регулятором гидротермического режима атмосферы через свой биогеохимический круговорот и прежде всего автотрофный биогенез. В дальнейшем этот регулирующий и стабилизирующий механизм лесного покрова приобретает глобальный характер, что достаточно полно описано в работах [23, 31].
Различное сочетание условий радиационного и термического режимов влечет за собой важные экологические следствия [9]. Существенное сокращение на островах Северо-Западной Паци-фики доли остаточной радиационной энергии на турбулентный теплообмен приводит к общему снижению температур деятельной поверхности и приземного слоя воздуха, на фоне чего возникают критические термоэдафотопы для тех или иных растительных сообществ. Абиотическая среда становится территориально более контрастной. Соответственно формируется определенное структурное и функциональное разнообразие региональных и локальных гео(эко-)систем в рамках одних и тех же фоновых (зональных) условий. На примере горных субарктических ландшафтов Южной Чукотки показано, что благодаря контрастности экологических режимов возможно «…. широкое сосуществование разнородных элементов в рамках одной флоры» [38, с. 33] и, добавим, одной географической зоны.
Подобное биогеоценотическое разнообразие определяется тем, что местные геоморфологические и эдафические факторы в различной степени преломляют структуру расходной части теплового баланса: уменьшают расходы тепла на турбулетный теплообмен и увеличивают их на испарение (и частично на прогревание почвы). Соответственно снижаются летние температуры воздуха. В локализованных типах местоположений это снижение идет вплоть до уровня, критического для данной зональной растительной формации. Именно в таких критических условиях возникают новые внутризональные типы экологических систем [9]. В этом, по-видимому, состоят энергетические основы изначальной роли островодужной Неопацифики как очаге возникновения локального и регионального разнообразия наземных экосистем, которое получает дальнейшее развитие на материке. Локальной контрастностью гидротермических условий обусловлены также пространственно-временная изменчивость островных экосистем, их высокая чувствительность к изменениям климата и способность к эволюционным преобразованиям.
ЗАКЛЮЧЕНИЕ
-
1. Установлена исключительно важная системоформирующая роль лесной фитобиоты в становлении и развитии молодых вулканических ландшафтов. «Запрограммированность» стратегии развития растительных сообществ на создание максимальной биомассы в лесных островодужных ландшафтах выражена гораздо сильнее, нежели в их зональных аналогах на соседнем материке. При этом исключительно большая доля зеленой массы в структуре продукционного процесса ведет к ускорению биологического круговорота как фактора устойчивости лесного сообщества. Соответственно обеспечивается выживание островных экосистем бореального пояса в неблагоприятных для них условиях «холодной» океаничности.
-
2. Бореальные леса Северо-Запрадной Не-опацифики характеризуются таким годовым оборотом растительного вещества, который относится к категории интенсивного метаболизма и по зональным меркам свойствен лишь субтропическим лесам. Особенно показательны в этом отношении буферные сообщества: ке-дровостланиково-березово-еловые в среднем ярусе низкогорья и каменноберезово-еловые на верхней границе темнохвойных лесов. Здесь нельзя не видеть зарождения прогрессирующих эволюционных тенденций в буферных лесных образованиях, столь характерных для Азиатско-Тихоокеанского мегаэкотона. Подтверждено известное положение глобальной эволюционной теории о том, что повышение интенсивности работы биогеоценозов и совершенствование биологических круговоротов лежат в основе эволюционного прогресса и составляют главное содержание эволюции. Такие эволюционные тенденции в растительном покрове зарождаются уже на первоначальной, вулканогенной стадии формирования континентальной биосферы.
-
3. Экстразональные, т.е. выходящие за рамки внутриматериковых зональных норм, показатели продуктивности и биологического круговорота в суббореальных и бореальных лесах молодых островодужных ландшафтов свидетельствуют о высокоэффективной работе их почвенно-биотического ядра. Интенсивная годовая оборачиваемость надземной фитобиоты и высокая скорость всего биологического круговорота в южнокурильских ландшафтах служат главными факторами их устойчивости в условиях отно-
- сительно низких температур вегетационного периода и наряду с вулканическими геотермами способствуют формированию суббореальных «климатически неоправданных» лесов. В этом видится один из главных феноменов организации лесных экосистем на начальном, островодужном этапе субаэрального биогеогенеза. С переходом в состояние окраинно-материковых, а затем внутриконтинентальных образований лесные экосистемы в целом снижают интенсивность своего функционирования: годичный оборот надземной фитомассы коренных лесов падает в два–три раза, а их биологический круговорот снижается до умеренной и даже заторможенной категории.
-
4. Проведена количественная оценка энергетических основ функциональной организации лесных экосистем островодужной Неопацифики. Выделены два основных мегатипа структуры теплового баланса наземных экосистем – материковый и океанический. Эти типы существенно различаются соотношениями двух основных составляющих расходной части баланса – эва-потранспирации и турбулетного теплообмена. Господство транспирационной составляющей теплового баланса в океаническом типе создает высокий уровень автотрофного биогенеза в лесных островных сообществах, что обеспечивает их устойчивость на начальном этапе субаэрального биогеогенеза.
РФФИ, 14-05-00032-а
Список литературы Продуктивность бореальных лесов в островном вулканическом ландшафте Северо-Западной Пацифики
- Андреев В.Л. Классификационные построения в экологии и систематике. М.: Наука, 1980. 142 с.
- Атлас Курильских островов/Российская академия наук. Институт географии РАН. Тихоокеанский институт географии РАН/Редкол.: Котляков В.М., Бакланов П.Я., Комедчиков Н.Н. (гл. ред.) и др. М.-Владивосток: ИПЦ «ДИК», 2009. 516 с.
- Базилевич Н.И. Продуктивность, энергетика и биогеохимия наземных экосистем Тихоокеанского кольца//Вопросы географии. Сб. 117. М.: Мысль, 1981. С. 146-208.
- Базилевич Н.И. и Родин Л.Е. Продуктивность и круговорот элементов в естественных и культурных фитоценозах (по материалам СССР)//Биологическая продуктивность и круговорот химических элементов в растительных сообществах. М.: Наука, 1971. С. 5-32.
- Базилевич Н.И., Титлянова А.А. Биологический круговорот на пяти континентах: азот и зольные элементы в природных наземных экосистемах. Новосибирск: Изд-во СО РАН, 2008. 380 с.
- Бобров Е.Г. Некоторые черты новейшей истории флоры и растительности Южной части Дальнего Востока//Бот. журнал. 1980. Т. 65. № 2. С. 172-183.
- Будаговский А.И. Испарение почвенной влаги. М.: Наука, 1964. 244 с.
- Быков Б.А. Геоботанический словарь. Алма-Ата: Изд-во «Наука» Казахской ССР, 1973. 214 с.
- Будыко М.И. Глобальная экология. М.: Высшая школа, 1977. 327 с.
- Будыко М.И. Эволюция биосферы. Л.: Гидрометеоиздат, 1984. 488 с.
- Воробьев Д.П. Растительность Курильских островов. М.-Л.: Изд-во АН СССР, 1963. 92 с.
- Ганзей К.С. Ландшафты и физико-географическое районирование Курильских островов.Владивосток: Дальнаука,2010. 214 с.
- Глазовская М.А. Геохимия природных и антропогенных ландшафтов СССР. М.: Высшая школа, 1988. 328 с.
- Иванов А.Н. Проблемы островного ландшафтоведения//Вопросы ландшафтоведения. Вып. 138. Горизонты ландшафтоведения. М.: «Кодекс», 2014. С. 138-158.
- Коломыц Э.Г. Бореальный экотон и географическая зональность: Атлас-монография. М.: Наука, 2005. 390 с.
- Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
- Коломыц Э.Г. Тихоокеанский мегаэкотон Северной Евразии. Часть I. Эволюционная экология боральных лесов. Raleigh, North Carolina, USA: Lulu Press, 2015. 248 с.
- Коломыц Э.Г., Сурова Н.А. Модели экологического пространства и устойчивости высокогорных лугов (На примере Центрального Кавказа)//География и природные ресурсы. 2014. № 4. С. 120-131.
- Коломыц Э.Г., Шарая Л.С. Количественная оценка функциональной устойчивости лесных экосистем//Экология. 2015. № 2. С. 83-94.
- Крестов П.В. Растительный покров и фитогеографические линии Северной Пацифики. Автореф. дис.. докт. биол. наук. Владивосток: Биолого-почв. ин-т ДВО РАН, 2006. 42 с.
- Манько Ю.И., Розенберг В.А. Высотная поясность растительности на вулкане Менделеева//Биол. ресурсы о. Сахалин и Курильских островов. Владивосток: Биолого-почв. ин-т ДВНЦ АН СССР, 1970. С. 65-71.
- Марков К.К., ред. Физическая география Мирового Океана. Л.: Наука, 1980. 362 с.
- Моисеев Н.Н., Александров В.В., Тарко А.Н. Человек и биосфера. Опыт системного анализа и эксперименты с моделями. М.: Наука, 1985. 271 с.
- Одум Ю. Основы экологии. Пер. с англ. М.: Мир, 1975. 740 с.
- Перельман А.И. Геохимия ландшафта. М.: Высшая школа, 1975. 341 с.
- Пузаченко Ю.Г., Скулкин В.С. Структура растительности лесной зоны СССР: Системный анализ. М.: Наука, 1981. 275 с.
- Раунер Ю.Л. Тепловой баланс растительного покрова. М.: Наука, 1972. 210 с.
- Руднев Н.И. Радиационный и тепловой баланс фитоценозов. М.: Наука, 1984. 111 с.
- Сочава В.Б. Геотопология как раздел учения о геосистемах//Топологические аспекты учения о геосистемах. Новосибирск: Наука, 1974. С. 3-86.
- Сочава В.Б. Введение в учение о геосистемах. Новосибирск: Наука, 1978. 319 с.
- Сукачев В.Н. Избранные труды. Т. 1. Основы лесной типологии и биогеоценологии. Л.: Наука, 1972. 418 с.
- Тимофеев-Ресовский Н.В., Тюрюканов А.Н. Об элементарных биохорологических подразделениях биосферы//Бюлл. Моск. об-ва испытат. природы. Т. LXXI. Вып. 1. М.: Изд-во МГУ, 1966. С. 123-132.
- Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. Краткий очерк теории эволюции. М.: Наука, 1969. 407 с.
- Тооминг Х.Г. Экологические принципы максимальной продуктивности посевов. Л.: Гидрометеоиздат, 1984. 264 с.
- Урусов В.М. Структура разнообразия и происхождение флоры и растительности юга Дальнего Востока. Владивосток: ТИГ ДВО РАН, 1993. 129 с.
- Урусов В.М., Чипизубова, М.Н. Общие закономерности географического распределения формаций и типов растительности. Генезис растительности//Геосистемы Дальнего Востока России на рубеже XX-XXI веков. Том I. Природные геосистемы и их компоненты. Владивосток: Дальнаука, 2008. С. 203-220.
- Шарая Л.С. Предсказательное картографирование лесных экосистем в геоэкологии//Поволжский экологический журнал. 2009, № 3. С. 249-257.
- Юрцев Б.А. Проблемы ботанической географии Северо-Восточной Азии. Л.: Наука, 1974. 159 с.
- Nakamura Y., Krestov P.V. Coniferous forests of the temperate zone of Asia. In Coniferous forests. Ser. Ecosystems of the World. 2005. Vol. 6. P. 163-220.
- Pietsch1 T.W., Bogatov V.V., Amaoka K., et al. Biodiversity and biogeography of the islands of the Kuril Archipelago//Journ. of Biogeogr. 2003.Vol. 30. P. 1297-1310.
- Shary P.A., Sharaya L.S., Mitusov A.V. Fundamental quantitative methods of land surface analysis//Geoderma. 2002. Vol. 107. № 1-2. P. 1-32.
