Произвольная ритмическая активность пальцев руки в условиях задержки дыхания

Автор: Ашуркова Елена Сергеевна, Рыжов Анатолий Яковлевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Физиология труда
Статья в выпуске: 8, 2008 года.
Бесплатный доступ
Выявлены возможности индивидуального анализа периодических процессов, произвольного движения кинематического звена пальцев рук в условиях обычного дыхания и при его задержке с оксигемометрическим контролем. Произвольная задержка дыхания сопровождается изменениями гипоксемического характера, определенным образом воздействующими на выполнение ритмической сенсомоторной работы пальцами руки. Это обусловлено некоторым дефицитом кислорода в работающих мышцах и в соответствующих нервных центрах.
Теппингтест, оксигенация крови, оксигемометр, статистические параметры
Короткий адрес: https://sciup.org/146116154
IDR: 146116154 | УДК: 612.822.3
Текст научной статьи Произвольная ритмическая активность пальцев руки в условиях задержки дыхания
Физиологическое понятие произвольности движения обусловлено осознанным действием, которое человек может выполнить или затормозить по собственному приказу и дать о нем словесный отчет (я делал так-то) [13]. Истинно произвольным действием являются двигательные акты, инициированные моторной сферой коры больших полушарий головного мозга, согласно многоуровневому построению движений [1; 2; 5]. Известно, что многие произвольные двигательные акты трудового и спортивного характера могут выполняться в экстремальных условиях или близких к таковым. Примером может служить гипоксия различных форм и режимов, индивидуальная устойчивость к которой достаточно вариабельна, поскольку человек в процессе трудовой и исследовательской деятельности осваивает космос, высотные, подземные и подводные регионы, характеризующиеся соответственно различными условиями газовой среды [7].
Система дыхания вместе с другими физиологическими системами обеспечивает двигательные акты, доставляя в нервную систему и работающие мышцы кислород, выводя избыток углекислого газа и тем самым сохраняя газовый гомеостазис [6]. Достаточно подробно изученным примером произвольных режимов дыхания является произвольная его задержка, обязательно сопровождающаяся сдвигами в газовом составе альвеолярного воздуха и артериальной крови [9; 20; 21].
Цель данной работы – физиологическая оценка ритмической активности пальцев рук при произвольной задержке дыхания в аспекте сенсомоторной работоспособности.
Методика . В исследовании в качестве испытуемых участвовали 15 практически здоровых мужчин 18–23 лет, предварительно проинструктированных об условиях эксперимента и соответствующей обстановке. В первой серии опытов испытуемые, находящиеся в положении сидя, производили ритмические движения пальцами при фиксированных предплечье и пясти (теппинг-тест), с максимально возможной частотой нажимов на рычаг со свободным ходом, как было подробно представлено в одной из наших предыдущих работ [3; 4; 16; 17; 19]. Во второй серии с помощью оксигемометра 057-М у испытуемых, находящихся в прежнем положении, определяли содержание в крови оксигемоглобина при произвольной задержке дыхания по общепринятой методике [7; 9; 10; 13; 19; 20]. Практически экспериенты данных серий были контрольными и служили своеобразным фоном для дальнейших основных исследований, имеющих комплексный характер.
Работа выполнена при поддержке гранта РГНФ 07-06-57607 а/ц
В третьей серии испытуемые совершали работу пальцами рук с максимальной частотой при фиксированной пясти на фоне задержки дыхания с оксигемометриче-ским контролем и выборочной графической регистрацией кривых оксигенации посредством фотооксигемографа.
Математическая обработка результатов, осуществляемая по компьютерной программе «Impulse», включала интервалографический, гистографический, автокорреляционный и фрактальный (по Харсту) анализ с оценкой репрезентативности выборок и достоверности различий результатов методами параметрической статистики по Стьюденту. Опыты проводились в помещении с постоянным микроклиматом, нормативным световым и шумовым режимами в первой половине дня, то есть на фоне неутомленных центральной нервной и нервно-мышечной систем.
Результаты и их обсуждение . Регистрация произвольных движений пальцев рук в первой серии показала, что в начале опыта длительность двигательного цикла (удар-замах) составляет 0,172±0,006 с, в середине – 0,188±0,006 с (Р<0,05), а в конце опыта отмечено увеличение длительности двигательного цикла до 0,198±0,007 с (Р<0,01). Подобная динамика двигательного цикла соответствует периодам врабатывания, оптимальной работоспособности и утомления. Аналогично изменяется мода (Мо) гистограммы движений от 0,161±0,008 с до 0,186±0,005 с (Р<0,01) и до 0,200±0,008 с (Р<0,01), причем амплитуда ее в начале опыта составляет 32,9±2,12% с тенденцией к постепенному увеличению к концу опыта до 33,7±2,1% [3]. В целом кривая теппинг-теста напоминает «кривую усталости», полученную еще Д. Уотсоном [20] при эргографической регистрации ритмической работы пальца. На ней сначала регистрируется максимальная высота поднятия груза, затем наблюдается снижение высоты с последующей некоторой задержкой данного процесса и дальнейшим, еще более крутым падением вплоть до наступления усталости и прекращения работы. Регистрируемая нами работа является не силовой, а сенсомоторной, и закономерности ее периодизации, с нашей точки зрения, скорее, всего иные.
На основании анализа автокорреляционных функций данного периодического процесса выявлено, что коэффициент r1, получаемый по первому сдвигу коррелируемого массива, в начале опыта составляет 0,357±0,063 О.Е., в середине – 0,109±0,055 О.Е. (Р<0,01) и к моменту его окончания – 0,156±0,066 О.Е. (Р<0,01). Таким образом, развитию утомления предшествует ослабление связи смежных интервалов ритмических движений, что в определенной мере характеризует повышение хаосогенности процесса. Это подтверждается фрактальными характеристиками теппинг-теста по показателю Харста [4], составившего в начале опыта 0,984±0,059 О.Е., в середине – 0,914±0,071 и в конце опыта – 0,857±0,077.
Во второй серии при произвольной задержке дыхания на вдохе мы смогли построить усредненную кривую оксигенации (рис. 1, А) с типичной фазностью [9; 15]. Следует отметить, что в условиях произвольной задержки дыхания существенную роль может играть сопутствующая гиперкапния, по величине которой чаще всего и осуществляется регулирующая стимуляция [11]. Кислород, пройдя альвеолярнокапиллярную мембрану, попадает в плазму крови, а затем диффузионно, вследствие градиента парциального давления, проходит через мембрану эритроцитов, где связывается с гемоглобином до полного его насыщения. Последующая оксигенация тканей, естественно, осуществляется с участием небольшой остаточной части растворенного в плазме кислорода (порядка 2%). Акт вдоха (инспирация), как известно, связан с расширением грудной клетки, в результате чего снижается давление в плевральной полости, что и создает условия для уменьшения давления в альвеолах. На уровне дыхательного центра продолговатого мозга важнейшая роль принадлежит информации от механорецепторов тканей легких, а также стенок бронхов и трахеи. Эти рецепторы растяжения возбуждаются при вдохе, а импульсы от них по афферентным волокнам блуждающего нерва поступают в дыхательный центр. Под их влиянием тормозится активность инспираторных нейронов продолговатого мозга, поскольку по мере растяжения легких вдох прекращается и начинается выдох [7; 8; 11; 14; 15;
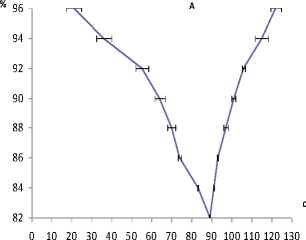
Рис. 1. Усредненные кривые оксигенации крови при произвольной задержке дыхания в условиях относительного покоя (А) и ритмической работы пальцами (Б)
Отмеченная в наших исследованиях фаза устойчивой оксигенации (96%), обусловленная кооперативными свойствами гемоглобина, т.е. способностью молекулы оксигемоглобина удерживать кислород пока парциальное напряжение его в артериальной крови не снизится до 80 мм рт. ст., составила 20,6±1,92 с. Фаза медленного снижения оксигенации (с 96,0 до 92%) составила 34 c, фаза быстрого снижения оксигенации (с 92,0 до 84%) – 29 c. Совместное действие гипоксемии и гиперкапнии в этих условиях формирует императивный стимул (непреодолимый позыв к вдоху) и задержка дыхания срывается, хотя, несмотря на начало дыхания оксигенация продолжает снижаться до 84–82% в течение 7 с, а в целом время обычной задержки дыхания у наших испытуемых составляет 89 с. В восстановительный период, состоящий из фаз быстрого (82–92%) и медленного (92–96%) повышения оксигенации, все показатели насыщения крови кислородом приближались к исходному уровню (рис. 1, А).
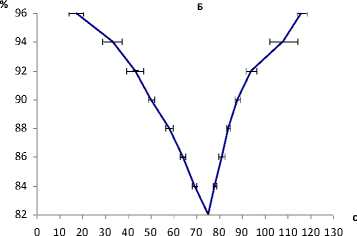
В третьей серии опытов анализировались ритмические движения пальцев в период задержки дыхания и, соответственно, снижения насыщения крови кислородом, где длительность фазы устойчивой оксигенации (96%) за счет расхода энергии и действия нейрогенного стимула несколько уменьшилась – до 16,5±1,61 с. Обусловленные вовлечением гуморальных факторов фазы медленной и быстрой гипоксемии, развитие которых, в общем синхронно с периодом локального снижения газообмена в работающих мышцах, составили 96–90 и 90–84% с длительностью их соответственно 33 и 19 с. Период снижения оксигенации после начала дыхания (84–82%) также имел тенденцию к укорочению до 6 с (рис. 1, Б). В итоге общее время задержки дыхания составило 75 с, что несколько меньше аналогичного времени в состоянии относительного покоя (рис. 1, Б). Общее время выполнения произвольных ритмических движений в условиях относительного покоя составило 105,0±15,751 с, при задержке дыхания – 51,733±4,733 с.
Следовательно, произвольная задержка дыхания в процессе выполнения работы сопровождается ускоренными изменениями гипоксемического характера. Это, на наш взгляд, связано с понижением содержания кислорода в крови, а также с некоторой кислородной недостаточностью и нарастающей гиперкапнией в двигательных центрах и, естественно, в самих работающих мышцах. Первичным же источником дыхательного дискомфорта, побуждающего человека прервать работу, следует считать усиленную афферентацию от рецепторов дыхательных мышц, преимущественно межреберных, напряжение которых особенно возрастает, когда нагрузка превышает анаэробный порог [11; 12; 13].
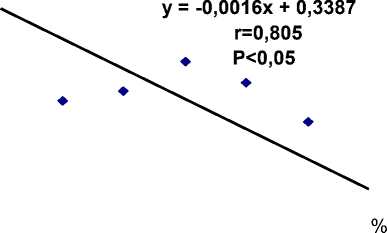
Уравнение регрессии (рис. 2), составленное на основе лонгитудинальной корреляции массивов, включающих данные оксигенации, демонстрирует общую и вполне прогнозируемую тенденцию к линейной зависимости, где коэффициент корреляции (r1), демонстрирующий сопряженность изменений оксигенации и длительности двигательного цикла, составляет 0,805 при Р<0,05. В результате на каждый процент понижения кислорода приходится прирост длительности двигательного цикла (ДДЦ) приблизительно в 2 мс (0,0018 с), от периода врабатывания до периода утомления с прекращением движений. Характерно, что ДДЦ, в устойчивый оксигенационный период (96%) составившая 0,181±0,003 с, по мере снижения оксигенации крови до 94% удлиняется до 0,194±0,003 и на протяжении нескольких периодов, составивших фазу медленной оксигенации, до 0,198±0,004 с (92%) и 0,201±0,003 с (90%). Затем к моменту окончания работы длительность двигательного цикла несколько уменьшается до 0,197±0,002 с (88%), 0,196 ± 0,001 с (86%), после чего прогрессивно растет до уровня 0,210 с по мере понижения оксигенации до 82–80%, четко соответствуя периоду утомления.
с0,21
0,2
0,19
0,18
82 84 86 88 90 92 94 96
Рис. 2. Регрессионная зависимость длительности двигательного цикла (ордината) от уровня оксигенации (абсцисса) при выполнении ритмической работы пальцами во время задержки дыхания
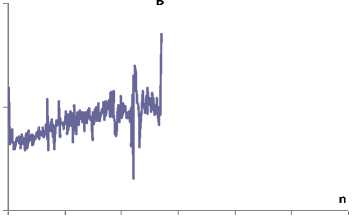
Индивидуальные показатели в первой серии (регистрация произвольных движений – теппинг-тест) эксперимента на примере испытуемого В., 20 лет (рис. 3, А), показали, что усредненная (ДДЦ), по данным всей работы, составляет 0,183±0,0004 с. В начале работы длительность двигательного равна 0,174±0,001 с, в середине – 0,183±0,001 с, конце 0,187±0,001 с. Аналогично изменяется Мо от 0,166 с до 0,189 с, что в принципе соответствует среднегрупповым значениям. Амплитуда Мо в начале опыта составляет 27%, в середине увеличивается до 35% и к концу опыта достигает 47%, что свидетельствует о развивающемся процессе нервно-мышечного утомления. Теснота смежных интервалов по величине первого сдвига (r1) имеет тенденцию к повышению от 0,245 О.Е. до 0,420 О.Е., что вписывается в динамику всей статистической выборки. Показатель Харста у данного испытуемого в начале опыта несколько повышен и составляет 0,837±0,034 О.Е., затем наблюдается тенденция к понижению – 0,680±0,018 О.Е. и по мере окончания опыта повышается до 0,745±0,016 О.Е.
0,3
с
А
0,3
0,2
)#м^^
0,1
n
с
0,2
0,1
0 100 200 300 400 500 600
0 100 200 300 400 500 600 700 800 900 1000 1100 1200
Рис. 3 Индивидуальные интервалограммы ритмических движений пальцев испытуемого В., 20 лет, в условиях свободного дыхания (А) и при произвольной его задержке (Б)
0,3
с
А
0,3
0,2
^^
n
0,1
0 40 80 120 160 200 240 280 320 360 400
0,2
0,1
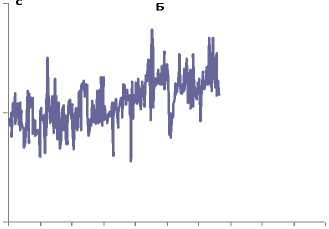
0 40 80 120 160 200 240 280 320 360 400
n
Рис. 4 Индивидуальные интервалограммы ритмических движений пальцев испытуемого Х.,19 лет, в условиях свободного дыхания (А) и при произвольной его задержке (Б)
Усредненная длительность двигательного цикла в третьей серии опытов (рис. 3, Б) во время работы с произвольной задержкой дыхания составила 0,187±0,001 с, практически не отличаясь от показателей первой серии, однако, число движений уменьшилось до 144 за время 26,9 с. Характерно, что и ДДЦ, составляющая в начале опыта 0,176±0,001 с, прогрессивно увеличивается до 0,189±0,001 с в середине и по мере окончания до 0,198±0,001 с. Показатель Мо менялся от 0,168 с до 0,184 и 0,189 с. Значение аМо увеличилось (по сравнению с первой серией) до 38 %, затем незначительно уменьшилось до 35 %, к окончанию работы данный показатель увеличился почти вдвое – до 66%. Если в первой серии величина коэффициента автокорреляции по первому сдвигу массива имела тенденцию к повышению, то здесь наблюдается ее снижение от 0,326 до 0,276 О.Е. Аналогично снижается и показатель Харста от 1,003±0,024 О.Е. до 0,801±0,024 О.Е., что свидетельствует о явном росте хаотичности данного процесса.
У испытуемого Х., 19 лет, (рис. 4, А) можно видеть несколько иные показатели времени теппинг-теста (29,9 С) и числа двигательных циклов (145). Усредненное значение ДДЦ, по данным всей работы, в первой серии экспериментов составила 0,206±0,001 с. Длительность двигательного цикла в начале работы 0,196±0,002 с, середине – 0,200±0,002 с и в конце работы 0,226±0,002 с. Мода увеличивается от 0,208 с до 0,217 с. Показатель амплитуды Мо в начале опыта увеличивается до 19 %, в середине опыта уменьшается до 13% и в момент окончания работы увеличивается до 17%. Величина коэффициента автокорреляции по первому сдвигу (r1) увеличивается от -0,034 О.Е. до 0,141 О.Е., тогда как показатель Харста, равный в начале опыта 1,327±0,054 О.Е., к середине его несколько повышается – до 1,417±0,034 О.Е. и в период окончания опыта снижается до 1,267±0,054 О.Е.
В третьей серии эксперимента в период задержки дыхания время теппинг-теста снижается до 26,3 с, число двигательных циклов – до 125. При этом усредненное значение ДДЦ, составившее 0,210±0,001 с), существенно не отличалось от аналогичных данных первой серии. В начале опыта ДДЦ – 0,186±0,001 с, середине – 0,206±0,002 и в конце опыта – 0,214±0,003 с. Аналогично изменяется показатель Мо повышается от 0,181 с до 0,203 с. Амплитуда Мо постепенно увеличивается от 14%, 16% до 18 %. Величина r1 в начале работы составляет 0,153 О.Е., затем увеличивается до 0,319 О.Е. и к моменту окончания работы уменьшается до -0,166 О.Е.. Показатель Харста увеличивается в начале работы до 1,461±0,038 О.Е., уменьшается в середине до 1,232±0,036 О.Е. и вновь увеличивается к окончанию работы – 1,442±0,052 О.Е. (рис. 4, Б), демонстрируя, таким образом, разнонаправленность хаотичности периодического процесса.
В итоге нашего исследования можно констатировать, что как групповой, так и индивидуальный анализ периодических произвольных движений кинематического звена пальцев рук в условиях обычного дыхания и при его задержке на фоне типичных оксигемометрических изменений позволяют установить некоторые особенности произвольной ритмической активности рук в гипоксически-гиперкапнических условиях. Опыты в определенной мере моделируют соответствующие экстремальные ситуации, и их результаты могут быть использованы при подготовке и реализации различного вида работ, осуществляемых в гипоксических условиях.
Выводы. 1. На примере молодых мужчин выявлены возможности индивидуального анализа периодических процессов произвольного ритмического движения кинематических звеньев пальцев рук в условиях обычного дыхания, а также на фоне его задержки под оксигемометрическим контролем.
-
2. Произвольная задержка дыхания, сопровождаемая изменениями гипоксически-гиперкапнического характера, определенным образом воздействует на характер ритмической сенсомоторной работы пальцев руки, уменьшая время теста и число двигательных циклов наряду с хаосогенными изменениями ритма интервалограммы.
-
3. Анализ теппинг-тестовых интервалограмм при произвольной задержке дыхания выявляет увеличение продолжительности двигательного цикла, моды распределения циклов и уменьшения их автокорреляционных взаимоотношений, что мы объясняем растущим дефицитом кислорода в работающих мышцах и в соответствующих нервных центрах.
VOLUNTARY RHYTHMIC ACTIVITY OF FINGERS DURING
Tver State University
The possibilities of individual analysis of periodical processes of voluntary movement of kinematical loop of fingers during the ordinary breathing and breath-holding with oxihemometric control were revealed. Voluntary breath-holding conducted with changes of hypoxemic character, influenced the execution of rhythmic sensomotor work of the fingers. It is caused by some oxygen deficiency in working muscles and related nerve centers.
