Проявление хозяйственно важных признаков у яровых гибридов мягкой пшеницы, отобранных с помощью MAS-технологии при скрещивании озимых сортов с яровыми донорами устойчивости к бурой ржавчине

Автор: Стасюк А.И., Леонова И.Н., Салина Е.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Зерновые: генетический потенциал, селекционное улучшение, агробиотехнологии
Статья в выпуске: 3 т.52, 2017 года.
Бесплатный доступ
В западносибирском регионе России мягкая яровая пшеница - основная зерновая культура, занимающая больше 40 % посевных площадей. Расширение генетического разнообразия этой культуры и создание более продуктивных и устойчивых к неблагоприятным факторам среды сортов всегда было ключевой проблемой селекции. В качестве перспективного источника хозяйственно важных признаков можно рассматривать генофонд озимой мягкой пшеницы, которая характеризуется разнообразием по толерантности к абиотическим стрессовым факторам, продуктивной кустистости и урожайности. Несмотря на то, что интрогрессия генного пула озимых культур в яровые давно используется для повышения урожайности яровых сортов, в настоящее время мало доступной информации о проявлении других хозяйственно важных признаков в потомстве от скрещивания озимых и яровых культур. Однако такие результаты важны для оценки комбинационной способности сортов озимой и яровой пшеницы и отбора потенциально ценных образцов по комплексу признаков. При применении классических методов селекции для идентификации озимых и яровых генотипов в потомстве от скрещивания озимых и яровых родительских форм требуется большой объем выборки и длительный период культивирования растительного материала. Использование MAS-технологий и маркеров, сцепленных с целевыми локусами, позволяет значительно сократить время получения новых форм. В рамках программы по созданию новых селекционных линий мягкой пшеницы по генам устойчивости к бурой ржавчине была использована схема гибридизации, где материнскими формами служили сорта озимой пшеницы Филатовка и Бийская озимая, адаптированные к условиям западносибирского региона. Интрогрессивную яровую линию 5366-180 ( Triticum aestivum / T. timopheevii ) и яровой сорт мягкой пшеницы Тулайковская 10, содержащий генетический материал пырея Thinopyrum intermedium, использовали в качестве доноров эффективных генов устойчивости к бурой ржавчине LrTt2 и Lr6Ai#2. Целью данной работы была оценка разнообразия яровых форм мягкой пшеницы, полученных в результате такой схемы гибридизации, по хозяйственно важным признакам и анализ влияния генотипа на их проявление. Отбор потомства с озимым и яровым образом жизни проводили в поколении F2 с помощью аллель-специфичных маркеров генов VRN-1. Для идентификации генотипов, содержащих гены устойчивости к бурой ржавчине, использовали микросателлитный маркер Xbarc232 и праймеры МF2 / MR1r2 и MF2 / MR4, специфичные соответственно для фрагментов транслокаций T. timopheevii и Th. intermedium. В результате гибридизации озимых сортов с яровыми донорами получили 122 растения F2, из которых отобрали 30 гомозиготных яровых растений с различным аллельным составом генов VRN-1. Из них 12 растений согласно данным молекулярного маркирования содержали чужеродные транслокации с генами Lr. Для сравнительного анализа количественных признаков было использовано 6 потомков F2, которые имели такой же аллельный состав генов VRN-1, как у ярового родителя, но отличались наличием или отсутствием Lr генов. Семейства F3-4, полученные в результате самоопыления соответствующих растений F2, были изучены в течение двух полевых сезонов по следующим признакам: период всходы-колошение, продуктивная кустистость, число и масса зерна с колоса, масса 1000 зерен. Результаты оценки периода всходы-колошение показали, что при одинаковом аллельном составе генов VRN-1 наблюдается значительная разница во времени колошения между гибридами F3-4 и яровыми донорами. В зависимости от года проведения эксперимента различия составляли от 3 до 8 сут, при этом было установлено, что наличие чужеродного генетического материала не оказывает влияния на анализируемый признак. Анализ гибридов F3-4 по признакам продуктивности показал, что на выраженность большинства из них в основном влияют факторы внешней среды. Значительный вклад генотипа в фенотипическое проявление признаков число зерен с колоса и масса 1000 зерен выявлен у гибридного образца 70-12, содержащего транслокацию от Th. intermedium. В работе показана эффективность маркер-ориентированного отбора гомозиготных генотипов в малых выборках на ранних этапах селекции. Вариация по признакам, которая наблюдается в семействах F3-4, позволяет подобрать селекционные образцы с оптимальной выраженностью хозяйственно важных признаков для дальнейшего использования.
Мягкая яровая пшеница, vrn-1 гены, lr гены, маркер-ориентированный отбор, признаки продуктивности
Короткий адрес: https://sciup.org/142214049
IDR: 142214049 | УДК: 633.11:631.523.4:631.527:577.21 | DOI: 10.15389/agrobiology.2017.3.526rus
Variability of agronomically important traits in spring wheat hybrids obtained by marker-assisted selection from crosses of winter wheat with spring wheat donors of resistance genes
Spring wheat is the main cereal crop in the West Siberian region of Russia and occupies more than 40 % of the acreage. The widening of the genetic diversity of this culture and the creation of new varieties with high productivity and resistance to environmental factors have always been a key problem of breeding. Winter wheat varieties which are characterized by a wide diversity of useful characters such as tolerance to abiotic stressors, productive tillers and yield can be used as a promising source of agronomically important traits. Despite the fact that the methodological approach to the introgression of the gene pool of winter wheat into the genome of spring wheat has been in use for a long time to increase the yield of spring varieties, at the moment there is little available information concerning the manifestation of valuable traits in offspring obtained from the winter and spring wheat hybridization. However, such results are important for assessing the combining ability of winter and spring wheat varieties and for the selection of potential parents by a complex of characters. For the identification of the winter and spring genotypes in the offspring resulting from the hybridization of winter and spring parents with the help of convenient breeding methods, a large sample size populations and a long period of cultivation of plant material are required. The application of MAS technologies and markers linked to the target loci can significantly decrease the time of development of new breeding lines. In creation of breeding lines resistant to leaf rust, the winter wheat varieties Filatovka and Biyskaya ozymaya, adapted for cultivation in the West Siberian region, were involved in a scheme of hybridization. Spring wheat introgression line 5366-180 ( Triticum aestivum / T. timopheevii ) и spring wheat cultivar Tulaikovskaya 10, containing genetic material of Thinopyrum intermedium were the donors of effective leaf rust resistance genes LrTt2 and Lr6Ai#2. The aim of this work was to evaluate the diversity of spring wheat forms obtained from these crosses on economically important traits and analysis of the genotype influence on their manifestation. The plants differing in winter and spring habits were selected in F2 progeny by means of allele-specific markers to the VRN-1 genes. To identify genotypes containing leaf rust resistance genes, a microsatellite marker Xbarc232 and primers MF2/MR1r2 and MF2/MR4 specific for T. timopheevii and Th. intermedium translocations, respectively, were used. As a result of hybridization of winter varieties with spring donors, 122 F2 plants were obtained, of which 30 homozygous spring plants with different allelic composition of VRN-1 genes were selected. Of these, 12 plants according to molecular data with Lr -specific primers contained alien translocations. For the comparative analysis of agronomic traits, six F2 plants were used which possessed the same allelic composition of VRN-1 genes as in the initial spring donors but differed in the presence or absence of Lr genes. F3-4 families obtained by selfing of the corresponding F2 plants were studied during two field seasons on the following traits: heading date, productive tillers, grain number per spike, grain weight per spike, and 1000-grain weight. The results of heading date evaluation showed that, despite the same allelic composition of the VRN-1 genes, there was a significant difference in the heading date between F3-4 hybrids and spring donors. Depending on the year of the field evaluation, the differences were three to eight days, and it was found that the presence of alien genetic material does not affect this trait. Analysis of the productivity traits of F3-4 hybrids indicated that the main impact to the manifestation of the traits was made by environmental factors. A significant contribution of the genotype to the phenotypic manifestation of the traits grain number per spike and 1000-grain weight was detected in the hybrid 70-12 containing the translocation from Th. intermedium. The obtained data demonstrated the effectiveness of MAS for the identification of homozygous genotypes with desired traits in the early breeding stages using small sample population. Agronomic trait variations observed in the F3-4 progenies allows to select breeding lines with the optimal expression of valuable traits for further breeding.
Текст научной статьи Проявление хозяйственно важных признаков у яровых гибридов мягкой пшеницы, отобранных с помощью MAS-технологии при скрещивании озимых сортов с яровыми донорами устойчивости к бурой ржавчине
В России яровая мягкая пшеница относится к наиболее распространенным злаковым культурам. При поиске перспективных источников цен-
∗ Работа выполнена при финансовой поддержке гранта РНФ ¹ 16-16-00011.
ных признаков для яровой пшеницы и их использования в разнообразных схемах гибридизации особый интерес вызывает генофонд озимой пшеницы как исходный материал для обогащения яровых форм по хозяйственно важным свойствам. Известно, что озимая мягкая пшеница в сравнении с яровой обладает большим разнообразием по толерантности к абиотическим факторам, продуктивной кустистости, фертильности колоса и урожайности (1, 2). Озимая пшеница издавна и по настоящее время привлекается для расширения разнообразия яровых сортов в мировой селекционной практике (3, 4). Советские ученые применяли скрещивание озимых и яровых сортов для изучения и отбора потомства по срокам колошения (5, 6). Было показано, что в поколениях F2 и F3 появились растения с более ранним, чем у ярового родителя, колошением, однако они были лишены способности к кущению и имели только один стебель.
Продуктивные и засухоустойчивые сорта яровой пшеницы получены на основе гибридизации озимых сортов Безостая 1, Аврора, Кавказ с яровыми (7, 8). В Международном центре по улучшению кукурузы и пшеницы (International Maize and Wheat Improvement Center — CIMMYT) при гибридизации озимых и яровых форм созданы селекционные линии и сорта яровой мягкой пшеницы с высокой урожайностью, короткостебельностью, устойчивостью к грибным болезням (9). От скрещивания ярового сорта с озимым в Швеции получен сорт яровой пшеницы Extrakolben, урожайность которого на 10 % выше, чем у лучших исходных селекционных сортов, что указывает на возможность значительно увеличить продуктивность яровых сортов при скрещивании с озимыми формами (10). Используя озимые сорта Безостая 1, Мироновская 808, Ранняя 12 в качестве материнской формы и яровой сорт Варденик 9 как опылитель, получили яровые гибридные формы, превосходящие по массе 1000 зерен ярового родителя (11). Кроме того, среди гибридных поколений были выделены константные линии, у которых высокая продуктивность сочеталась с хорошим качеством зерна.
Сообщалось об изучении комбинационной способности (12, 13) и гетерозиса по признакам, определяющим продуктивность, у гибридов от скрещивания форм пшеницы, которые различаются образом жизни (14-17). Однако до настоящего времени мало доступной информации о проявлении хозяйственно важных признаков в потомстве от таких скрещиваний.
В последние годы при анализе селекционного материала в дополнение к традиционным методам используют маркер-ориентированный отбор (marker-assisted selection, MAS). К его основным преимуществам можно отнести выявление генотипов, гомозиготных по целевому признаку, с помощью хромосом-специфичных маркеров и возможность обнаружения целевых генотипов на выборках малого размера (18).
Для создания нового селекционного материала и переноса эффективных генов устойчивости к бурой ржавчине в геном озимой и яровой мягкой пшеницы нами была использована схема гибридизации, где в качестве реципиентов взяты озимые сорта мягкой пшеницы, адаптированные для произрастания в западносибирском регионе. Донорами служили яровые формы, содержащие чужеродный генетический материал с генами резистентности, унаследованными от Triticum timopheevii и Thynopyrum intermedium (19, 20). Получение озимых генотипов с помощью молекулярных маркеров для выявления генов, контролирующих озимый или яровой образ жизни и устойчивость к бурой ржавчине, и их оценка по зимостойкости и восприимчивости к патогену, описаны нами ранее (21). В настоящем сообщении впервые представлены результаты изучения яровых потомков от скрещивания озимых и яровых родителей по признаку период всхо- ды—колошения и компонентам урожайности, а также исследовано влияние генетического материала, унаследованного от Triticum timopheevii и Thynopy-rum intermedium, на проявление анализируемых признаков.
Цель работы — сравнение продуктивных свойств у яровых гибридов пшеницы с интрогрессированной устойчивостью к бурой ржавчине.
Методика. При гибридизации материнскими формами послужили озимые сорта мягкой пшеницы Филатовка и Бийская озимая, донорами — яровая интрогрессивная линия 5366-180 ( T. aestivum / T. timopheevii ) и сорт мягкой пшеницы Тулайковская 10 ( T. aestivum / Th. intermedium ), несущие локусы устойчивости к бурой ржавчине — соответственно LrTt2 и Lr6Ai#2 (19, 20). Перед началом гибридизации озимые сорта яровизировали в течение 60 сут при температуре 3 ° С. Скрещивание с яровыми донорами генов устойчивости и выращивание гибридов F1 и F2 проводили в условиях гидропонной теплицы (Центр коллективного пользования «Лаборатория искусственного выращивания растений», ФИЦ Институт цитологии и генетики).
Полученные в результате гибридизации растения F2 генотипирова-ли с помощью аллель-специфичных маркеров для генов VRN-1 и чужеродного генетического материала, содержащего гены Lr . Геномную ДНК выделяли из листьев молодых растений с использованием бисульфита натрия Na2S2O5 согласно описанию (22). Яровые и озимые растения отбирали с помощью маркеров доминантных и рецессивных аллелей Vrn-A1 , Vrn-B1 и Vrn-D1 . Для отбора растений, несущих ген LrTt2 от Triticum timopheevii , использовали SSR маркер Xbarc232 (23), для идентификации локуса Lr6Ai#2 от Thinopyrum intermedium — разработанные нами две пары праймеров МF2 / MR1r2 и MF2 / MR4 (24). Структура праймеров к генам VRN-1 , условия ПЦР для праймеров к генам VRN-1 и праймеров МF2 / MR1r2 и MF2 / MR4 (25), а также протокол ПЦР для маркера Xbarc232 (22) описаны ранее. ПЦР проводили на амплификаторе Bio-Rad T-100 Thermal Cycler, разделение продуктов ПЦР — в 1,5 % агарозном геле в присутствии бромистого этидия, визуализацию — с помощью системы гель-документи-рования Gel Doc XR+ («Bio-Rad», США).
Потомство F3-4 от самоопыления яровых растений F2, отобранных с помощью молекулярных маркеров, оценивали в условиях опытного поля (Новосибирская обл., пос. Краснообск, 2014 и 2016 годы) по признакам период всходы—колошение, продуктивная кустистость, число зерен в колосе, масса зерна с колоса и масса 1000 зерен. Посев (в 2014 году 14 мая, в 2016 году — 17 мая) проводили вручную в 2 повторностях (контроль — исходные яровые формы). Растения выращивали на делянках шириной 1 м по 40-60 зерен в ряду с расстоянием между рядами 20 см. Периодом всходы— колошение считали время от появления всходов до выхода колоса из трубки. Число продуктивных побегов определяли как число всех озерненных побегов растения. Массу 1000 зерен рассчитывали по 2-3 колосьям для 20 растений каждого образца. Для статистической обработки данных по остальным признакам случайным образом выбирали 25 растений.
При определении достоверности различий между средними значениями двух выборочных совокупностей использовали t- критерий Стьюдента. Образцы сравнивали с помощью факторного дисперсионного анализа, достоверность оценивали по F -критерию Фишера, вклад факторов рассчитывали согласно Л.А. Васильевой (26). Статистическую обработку данных проводили с помощью пакета программ Statistica 10.0 («StatSoft, Inc.», США). В таблицах и на рисунках представлены средние арифметические ( x ) и ошибка средней ( sx ) .
Результаты. В годы испытаний метеоусловия в период вегетации растений различались по количеству осадков и температурному режиму. В 2014 году они были близки к среднемноголетним, при этом в мае наблюдался дефицит тепла на фоне избыточного увлажнения. В 2016 году отмечалось снижение влагообеспеченности в течение всего полевого сезона при более высоких температурах, чем в 2014 году.
Озимые сорта мягкой пшеницы Филатовка и Бийская озимая, адаптированные к почвенно-климатическим условиям Западной Сибири, обладают высокой зимостойкостью, высокопродуктивны, устойчивы к абиотическим факторам, но чувствительны к большинству рас бурой ржавчины (2). При гибридизации сортов Филатовка и Бийская озимая с интрогрессивной линией 5366-180 и сортом Тулайковская 10 получили 122 потомка, из которых на стадии F2 с помощью молекулярного маркирования отобрали 30 гомозиготных яровых растений с различным аллельным составом генов VRN-1 . Из них 12 растений содержали чужеродные транслокации с генами Lr . В качестве примера приведена электрофореграмма продуктов ПЦР-амплификации геномной ДНК родителей и потомства F2 с SSR маркером Xbarc232 (рис. 1). У озимых сортов Бийская озимая и Тулай-ковская 10 в присутствии Xbarc232 синтезировался фрагмент 200 п.н., специфичный для длинного плеча хромосомы 5B, а у интрогрессивной линии 5366-180 и растения ¹ 6 из популяции F2 он отсутствовал (нуль-аллель), что свидетельствует о наличии чужеродной транслокации с геном LrTt2 .
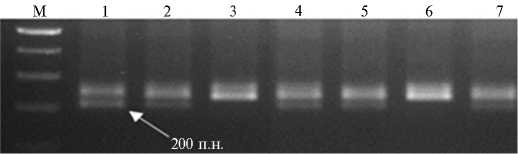
M — маркер длины фрагментов (100 bp ladder, ЗАО «Биосан», зывает на диагностический фрагмент 200 п.н.
Рис. 1. Пример использования ДНК-маркера Xbarc232 для идентификации генотипов с геном устойчивости к бурой ржавчине LrTt2 : 1 и 2 — соответственно сорта Бийская озимая и Тулайковская 10; 3 — интрогрессивная линия 5366-180; 4-7 — растения F 2 ; . Новосибирск). Стрелка ука-
1. Характеристика растений F2 мягкой пшеницы от разных комбинаций скрещивания по аллельному составу генов VRN-1 и наличию генов Lr
|
Комбинация скрещивания, сорт, образец |
Номер растения |
Аллельный состав генов VRN-1 |
Наличие генов Lr |
|
Филатовка ½ 5366-180 |
90-7 |
Vrn-A1a Vrn-B1c vrn-D1 |
LrTt2 |
|
88-11 |
Vrn-A1a Vrn-B1c vrn-D1 |
LrTt2 |
|
|
88-3 |
Vrn-A1a Vrn-B1c vrn-D1 |
- |
|
|
89-12 |
Vrn-A1a Vrn-B1c vrn-D1 |
- |
|
|
5366-180 |
Контроль |
Vrn-A1a Vrn-B1c vrn-D1 |
LrTt2 |
|
Бийская озимая ½ Тулайковская 10 |
70-12 |
Vrn-A1a Vrn-B1a vrn-D1 |
Lr6Ai#2 |
|
71-12 |
Vrn-A1a Vrn-B1a vrn-D1 |
- |
|
|
Тулайковская 10 |
Контроль |
Vrn-A1a Vrn-B1a vrn-D1 |
Lr6Ai#2 |
|
П р и м е ч а н и е . Прочерки означают отсутствие генов Lr. |
|||
Для сравнительного анализа хозяйственно ценных признаков использовали 6 потомков F2, которые имели такой же аллельный состав генов VRN-1 , как у ярового родителя, но отличались от него наличием или отсутствием Lr генов (табл. 1). Проверка гибридов, содержащих, согласно результатам молекулярного анализа, гены Lr , выявила у всех низкую восприимчивость к западносибирской популяции возбудителя бурой ржавчины Puccinia triticina Erikss. (данные не представлены).
Известно, что комбинация доминантных и рецессивных аллелей генов VRN-1 не только определяет тип развития, но и влияет на скорость развития и время колошения растений (27, 28). Также существует предположение о более раннем колошении и более высокой урожайности у сор- тов мягкой пшеницы, несущих два доминантных гена Vrn-A1 и Vrn-B1 (29). Оценка времени колошения у яровых потомков F3-4, полученных от скрещивания озимых сортов и яровых доноров генов устойчивости, показала, что гибридные растения выколашивались значительно позже ярового родителя (рис. 2). Так, для растений из комбинации скрещивания Филатов-ка ½ 5366-180 разница в сроках колошения по сравнению с яровым родителем составляла в среднем 3,3-4,5 сут в 2014 году и 4,2-8,6 сут — в 2016 году (рис. 2, А). Следует отметить отсутствие различий по времени колошения у образцов, содержащих и не содержащих чужеродные транслокации с генами Lr. В потомстве от скрещивания сортов Бийская озимая и Тулайковская 10 (см. рис. 2, Б) линии 70-12 и 71-12 по аллельному составу VRN-1 генов были идентичны родительскому сорту Тулайковская 10, различия заключались в присутствии чужеродной интрогрессии у линии 70-12. Оценка времени колошения показала, что независимо от года полевых испытаний обе линии выколашивались позднее родительского сорта.
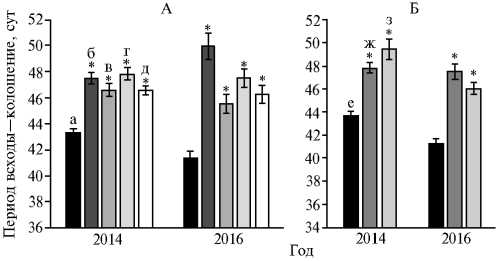
ное поле, Новосибирская обл.).
при p < 0,001.
Рис. 2. Длительность периода всходы—колошение в разные годы у яровых растений F 3-4 из комбинаций сорт Филатовка ½ интрогрессивная линия 5366-180 (донор LrTt2 гена) (А) и сорт Бийская озимая ½ сорт Тулай-ковская 10 (донор Lr6Ai#2 гена) (Б) в сравнении с родительскими формами: а — 5366-180 (контроль), б, в, г, д — линии 90-7, 88-11, 88-3, 89-12, е — Тулайковская 10 (контроль), ж, з — линии 70-12, 71-12 (опыт-
* Различия между гибридными линиями и родительскими формами статистически значимы
Число зерен в колосе в комбинации Филатовка ½ 5366-180 у всех изучаемых гибридов достоверно увеличилось в 2014 году, но в 2016 году разницы не наблюдали, а в скрещивании Бийская озимая ½ Тулайковская 10 образец 70-12 с транслокацией от Th. intermedium достоверно превосходил исходный сорт в оба года, тогда как линия 71-12 без чужеродной транслокации не отличалась от ярового родителя (табл. 2). По массе зерна с колоса в первой комбинации достоверно превосходили контроль линии 90-7, 88-11 и 88-3 в 2014 году, во второй — линия 70-12 (в 1,2 раза) в 2016 году. Масса 1000 зерен у образцов 88-11 и 88-3 в потомстве Филатовка ½ 5366-180 была соответственно 35,7 и 35,1 г, что достоверно ниже, чем у линии 5366180 (38,2 г), а образцы 90-7 и 89-12 не отличались от контроля. В 2016 году у всех образцов из первой комбинации не проявились достоверные различия с контролем по массе 1000 зерен, а во второй комбинации его выраженность была значительно ниже у образца 70-12 в оба года (см. табл. 2). Следует также отметить, что почти у всех исследованных образцов, включая родителей, показатели по количественным признакам снижались в 2016 году, что может быть обусловлено сложившимися погодными условиями. О значительном влиянии факторов внешней среды на количественные признаки свидетельствовали и данные факторного анализа (табл. 3).
Факторный дисперсионный анализ, выполненный на основании результатов оценки хозяйственно-важных признаков, установил, что в проявление признака период всходы—колошение значительный вклад вносит генотип гибридных линий, в то время как продуктивная кустистость зависела только от условий внешней среды (см. табл. 3). Данные факторного анализа
-
2. Выраженность хозяйственно важных признаков у семейств F3-4 и родительских форм яровой мягкой пшеницы в разные годы в двух комбинациях скрещивания ( x ± sx , опытное поле, Новосибирская обл.)
Образец, сорт, линия
п Признак
2014 год
1 2016 год
С о р т Ф и л а т о в к а ½ о б р а з е ц 5366-180
5366-180 (контроль)
Продуктивная кустистость, шт.
2,50±0,14
3,13±0,21
Число зерен в колосе, шт.
28,79±0,71
23,45±1,21
Масса зерна с колоса, г
1,15±0,04
0,64±0,04
Масса 1000 зерен, г
38,15±0,60
25,79±0,79
90-7
Продуктивная кустистость, шт.
3,23±0,21**
2,31±0,26*
Число зерен в колосе, шт.
32,74±1,23*
25,58±2,22
Масса зерна с колоса, г
1,32±0,06*
0,65±0,07
Масса 1000 зерен, г
36,64±0,57
24,17±1,58
88-11
Продуктивная кустистость, шт.
2,46±0,19
2,60±0,23
Число зерен в колосе, шт.
36,40±1,52***
26,50±1,45
Масса зерна с колоса, г
1,38±0,07**
0,70±0,04
Масса 1000 зерен, г
35,65±0,70**
25,63±0,72
88-3
Продуктивная кустистость, шт.
2,64±0,17
2,17±0,25**
Число зерен в колосе, шт.
36,03±1,34***
20,27±1,69
Масса зерна с колоса, г
1,37±0,06**
0,55±0,06
Масса 1000 зерен, г
35,09±0,72**
24,34±1,14
89-12
Продуктивная кустистость, шт.
2,41±0,15
2,68±0,36
Число зерен в колосе, шт.
31,59±1,16*
22,06±1,83
Масса зерна с колоса, г
1,26±0,06
0,61±0,06
Масса 1000 зерен, г
36,93±0,93
26,15±1,25
Сорта Бийская озимая ½
Тулайковская 10
Тулайковская 10 (контроль)
Продуктивная кустистость, шт.
2,28±0,13
2,71±0,17
Число зерен в колосе, шт.
33,52±1,14
28,98±1,30
Масса зерна с колоса, г
1,18±0,05
0,87±0,04
Масса 1000 зерен, г
33,93±0,57
28,96±0,58
70-12
Продуктивная кустистость, шт.
2,03±0,22
1,64±0,16***
Число зерен в колосе, шт.
41,19±2,39**
38,94±1,93***
Масса зерна с колоса, г
1,33±0,12
1,04±0,08*
Масса 1000 зерен, г
29,22±1,30***
26,48±1,08*
71-12
Продуктивная кустистость, шт.
2,27±0,19
2,19±0,20*
Число зерен в колосе, шт.
35,22±2,11
32,30±1,62
Масса зерна с колоса, г
1,21±0,09
0,79±0,06
Масса 1000 зерен, г
32,68±1,02
23,27±0,99***
*, **, *** Различия между гибридными линиями и родительскими формами статистически значимы соот-
ветственно при p < 0,05; p < 0,01; p < 0,001.
-
3. Факторный дисперсионный анализ, выполненный по результатам оценки хозяйственно ценных признаков у гибридов F3-4 и родительских форм яровой мягкой пшеницы в двух комбинациях скрещивания (опытное поле, Новосибирская обл., 2014 и 2016 годы)
Признак
Филатовка ½ 5366-180
Бийская озимая ½ Тулайковская 10
интрогрессии
без интрогрессий
интрогрессии
без интрогрессий
r wg r we
r wg r we
r wg r we
r wg r we
Всходы—колошение
0,94*****
0,05
97,1*****
2,70
0,91*****
0,08
0,99*****
0,01
Продуктивная кустистость
0,01
0,98*****
0,05
0,92*****
0,12
0,86****
0,01
0,98*****
Число зерен в колосе
0,22**
0,75****
0,09
0,78****
0,83****
0,16*
0,23
0,74**
Масса зерна с колоса
0,05
0,92*****
0,04
0,93*****
0,20
0,80***
0,03
0,95**
Масса 1000 зерен
0,02
0,96***
0,02
0,98***
0,55***
0,41*
0,26
0,68**
П р и м е ч а н и е . rwg и rwe — коэффициенты корреляции, определяющие вклад соответственно генотипа и внешней среды в фенотипическое проявление признака.
*, **, *** , ****, ***** Степень влияния генотипа и внешней среды статистически значимы соответственно при p < 0,05, p < 0,01, p < 0,001, p < 0,0001 и p < 0,00001.
позволяют предположить, что наличие фрагментов транслокаций в геноме гибридных образцов не влияет на признаки период всходы—колошение и продуктивная кустистость. Наоборот, у образца 70-12 из комбинации Бийская озимая ½ Тулайковская 10 на число зерен в колосе и массу 1000 зерен в большей степени влияет генотип, тогда как у образца 71-12 — условия окружающей среды. Не исключено, что вариации признаков у этих гибридов связаны с влиянием чужеродного генетического материала, поскольку линия 70-12 содержит фрагмент транслокации от Th. intermedium .
У всех изученных яровых линий, отобранных из потомства от скре- щивания с озимыми сортами, происходила задержка в сроках колошения относительно ярового родителя, независимо от их гаплотипа по генам VRN-1 и присутствия чужеродных транслокаций. Следует отметить безуспешность попыток ученых в начале XX века получить более ранние яровые образцы от скрещивания яровых и озимых сортов пшеницы (3, 5, 6). В первую очередь, причина в том, что если яровость обеспечивается присутствием доминантного гена VRN-1, то сроки колошения зависят от взаимодействия многих регуляторных и структурных генов, которые в обсуждаемом случае могут быть привнесены от озимого генотипа (30, 31). Кроме того, внутри линий наблюдалось варьирование как по срокам колошения, так и по остальным хозяйственно важным признакам (см. рис. 2, табл. 2), что дает возможность отбора яровых форм растений, превосходящих родительские формы по продуктивности и близких по срокам колошения. Нельзя исключить вероятность того, что при использовании в скрещивании с яровыми пшеницами озимых сортов европейского региона, задержка в сроках колошения потомства ярового образа жизни может быть существенно меньше.
Таким образом, представленные результаты свидетельствуют об эффективности аллель-специфичных маркеров при отборе генотипов по целевым локусам. Генотипирование образцов одновременно по нескольким генетическим локусам позволяет существенно уменьшить размер выборки и идентифицировать гомозиготное потомство на стадии популяции F2. Значительное удлинение сроков колошения у яровых потомков, полученных в результате гибридизации озимых и яровых форм мягкой пшеницы, по-видимому, не связано, с эффектом генов VRN-1 , детерминирующих сроки колошения. Оценка яровых семейств F3-4 по количественным признакам не выявила у полученных образцов достоверных отличий от родительских форм. Изучение образцов в течение двух полевых сезонов свидетельствует, что основной вклад в проявление этих признаков вносят факторы внешней среды. Исключение составляет образец 70-12, у которого достоверное влияние генотипа на проявление количественных признаков можно объяснить наличием чужеродного генетического материала.
Список литературы Проявление хозяйственно важных признаков у яровых гибридов мягкой пшеницы, отобранных с помощью MAS-технологии при скрещивании озимых сортов с яровыми донорами устойчивости к бурой ржавчине
- Sharma S., Chaudhary H.K. Combining ability and gene action studies for yield contributing traits in crosses involving winter and spring wheat genotypes. Acta Agron. Hungarica, 2009, 57: 417-423 ( ) DOI: 10.1556/AAgr.57.2009.4.4
- Козлов В.Е. Агротехнические и селекционные слагаемые успеха внедрения мироновских сортов озимой пшеницы в СССР как основа для работы по внедрению в Сибири вновь созданных сортов, зимостойких в условиях региона. Вавиловский журнал генетики и селекции, 2013, 17(3): 541-557.
- Nilsson-Ehle H. Selection of spring wheat in Sweden. Sveriges Utsadesforenings Tidskrift, Malmo, 1917, 28: 51-76 (Цит. по Вавилов Н.И. Научные основы селекции пшеницы. М., 1935).
- Mishra C.N., Venkatesh K., Kumar S., Singh S.K., Tiwari V., Sharma I. Harnessing winter wheat variability for enhancement of yield in spring wheat. International Journal of Bioresource and Stress Management, 2013, 4(2 special): 375-377.
- Вавилов Н.И., Кузнецова Е.С. О генетической природе озимых и яровых растений. Изв. агрон. фак. Саратовского ун-та, 1921, 1: 1-25.
- Лысенко Т.Д., Презент И.И. Селекция и теория стадийного развития растения. М., 1935.
- Лукьяненко П.П. Селекция и семеноводство озимой пшеницы. Изб. тр. М., 1973.
- Дорофеев В.Ф., Удачин Р.А., Семенова Л.В., Новикова М.В., Градчанинова О.Д., Шитова И.П., Мережко А.Ф., Филатенко А.А. Пшеницы мира. Л., 1987.
- Rajaram S., Borlaug N.E., van Ginkel M. CIMMYT international wheat breeding. In: Bread wheat: improvement and production/B.C. Curtis, S. Rajaram, G.H. Macpherson (eds.). FAO, Rome, 2002.
- Вавилов Н.И. Научные основы селекции пшеницы. М., 1935.
- Никогосян Е.Е., Сафарян Г.Е. Качество зерна пшеницы, полученной методом гибридизации озимых с яровыми формами. Биологический журнал Армении, 1977, ХХХ(9): 80-85.
- Kant L., Mahajan V., Gupta H.S., Pandey M.C. Winter ½ spring wheat hybridization: a potential source for yield advancement. International Journal of Plant Breeding, 2008, 2(2): 69-73.
- Kapoor E., Mondal S.K., Dey T. Combining ability analysis for yield and yield contributing traits in winter and spring wheat combinations. Journal of Wheat Research, 2011, 3(1): 52-58.
- Рутц Р.И., Леонтьев С.И. Проявление гетерозиса гибридов яровой пшеницы с озимой в зависимости от условий выращивания. Научные труды Омского сельскохозяйственного института им. С.М. Кирова, 1967, 69(4): 75-82.
- Grant M.N., McKenzie H. Heterosis in F1 hybrids between spring and winter wheat. Can. J. Plant Sci., 1970, 50: 137-140.
- Baric M., Sarcevic H., Keresa S. Analysis of yield сomponents of F1 hybrids of crosses between spring and winter types (Triticum aestivum L.). Agriculturae Conspectus Scientificus, 2004, 69(1): 11-15.
- Моисеенко Л.М., Клыков А.Г., Коновалова И.В., Богдан П.М. Использование озимых сортов в селекции яровой мягкой пшеницы с целью повышения генетического потенциала продуктивности. Достижения науки и техники АПК, 2011, 12: 28-30.
- Leonova I.N. Molecular markers: Implementation in crop plant breeding for identification, introgression and gene pyramiding. Rus. J. Genetics: Applied Research, 2013, 3(6): 464-473 ( ) DOI: 10.1134/S2079059713060051
- Timonova E.M., Leonova I.N., Röder M.S., Salina E.A. Marker-assisted development and characterization of a set of Triticum aestivum lines carrying different introgressions from the T. timopheevii genome. Mol. Breed., 2013, 31: 123-136 ( ) DOI: 10.1007/s11032-012-9776-x
- Salina E.A., Adonina I.G., Badaeva E.D., Kroupin P.Yu., Stasyuk A.I., Leonova I.N., Shishkina A.A., Divashuk M.G., Starikova E.V., Khuat T.M.L., Syukov V.V., Karlov G.I. A Thinopyrum intermedium chromosome in bread wheat cultivars as a source of genes conferring resistance to fungal diseases. Euphytica, 2015, 204: 91-101 ( ) DOI: 10.1007/s10681-014-1344-5
- Стасюк А.И., Леонова И.Н., Щербань А.Б., Салина Е.А. Использование молекулярных маркеров для создания озимых форм мягкой пшеницы (Triticum aestivum L.), устойчивых к грибным болезням. Мат. 2-й Межд. конф. «Генофонд и селекция растений». Новосибирск, 2016: 59.
- Kiseleva A.A., Shcherban A.B., Leonova I.N., Frenkel Z., Salina E.A. Identification of new heading date determinants in wheat 5B chromosome. BMC Plant Biol., 2016, 16(Suppl 1): 8 ( ) DOI: 10.1186/s12870-015-0688-x
- Song Q.J., Shi J.R., Singh S., Fickus E.W., Costa J.M., Lewis J., Gill B.S., Ward R., Cregan P.B. Development and mapping of microsatellite (SSR) markers in wheat. Theor. Appl. Genet., 2005, 110: 550-560 ( ) DOI: 10.1007/s00122-004-1871-x
- Салина Е.А., Леонова И.Н., Щербань А.Б., Стасюк А.И. Способ создания линий озимой мягкой пшеницей с комплексной устойчивостью к бурой и стеблевой ржавчине и мучнистой росе. Федеральный исследовательский центр Институт цитологии и генетики СО РАН (РФ). Патент РФ № 2598275 от 20.09.2016. Заявл. 29.07.2015. Опубл. 30.08.2016.
- Лихенко И.Е., Стасюк А.И., Щербань А.Б., Зырянова А.Ф., Лихенко Н.И., Салина Е.А. Изучение аллельного состава генов Vrn-1 и Ppd-1 у раннеспелых и среднеранних сортов яровой мягкой пшеницы Сибири. Вавиловский журнал генетики и селекции, 2014, 18(4/1): 691-703.
- Васильева Л.А. Статистические методы в биологии. Новосибирск, 2004.
- Стельмах А.Ф. Генетика типа развития и продолжительность вегетационного периода мягких пшениц. Селекция и семеноводство, 1981, 48: 8-15.
- Trevaskis B., Hemming M.N., Dennis E.S., Peacock W.J. The molecular basis of vernalization-induced flowering in cereals. Trends in Plant Science, 2007, 12(8): 352-357 ( ) DOI: 10.1016/j.tplants.2007.06.010
- Файт В.И., Стельмах А.Ф. Генетический контроль типа и скорости развития яровой пшеницы Западной Сибири. Сибирский вестник сельскохозяйственной науки, 1993, 2: 32-41.
- Chen A., Li C., Hu W., Lau M.Y., Lin H., Rockwell N.C., Martinc S.S., Jernstedt J.A., Lagariasc J.C., Dubcovsky J. Phytochrome C plays a major role in the acceleration of wheat flowering under long-day photoperiod. PNAS USA, 2014, 111: 10037-10044 ( ) DOI: 10.1073/pnas.1409795111
- Pearce S., Kippes N., Chen A., Debernardi J.M., Dubcovsky J. RNA-seq studies using wheat phytochrome B and phytochrome C mutants reveal shared and specific functions in the regulation of flowering and shade-avoidance pathways. BMC Plant. Biol., 2016, 16: 141 ( ) DOI: 10.1186/s12870-016-0831-3
