Проявление эффекта гормезиса у дрожжевых клеток, облученных ионизирующим излучением

Бесплатный доступ
Проведено сравнительное изучение количественных закономерностей отмирания гаплоидных и диплоидных дрожжевых клеток в непитательной среде, облученных ионизирующим излучением. Отмирание клеток наблюдали в процессе хронического облучения (γ-кванты 137Cs) и после острого облучения (γ-кванты 60Со) в различных дозах. Установлено, что хроническое (в течение одного месяца при 30 0C) действие ионизирующего излучения в диапазоне мощностей доз, составляющих 310-2 - 102 естественных радиационных фонов (ЕРФ), приводило к уменьшению скорости отмирания дрожжевых клеток. При этом отмечался обратный эффект мощности дозы: меньшие дозы и мощности доз ионизирующего излучения были более эффективны по сравнению с более высокими дозами и мощностями доз ионизирующего излучения. Однократное острое облучение дрожжей также замедляло отмирание выживших после облучения клеток, происходившее в термостате при 37 и 40 0C. Процесс этот регистрировался не только в области малых доз ионизирующего излучения, снижающих выживаемость клеток до 70-90 %, но и в области больших доз, когда доля выживших клеток составляла несколько процентов. Показано, что эффекты радиационного гормезиса были слабо выражены для гаплоидных штаммов. Полученные результаты интерпретируются в рамках традиционной гипотезы о роли систем репарации в проявлении гормезиса.
Короткий адрес: https://sciup.org/170169907
IDR: 170169907
Hormesis effect display in yeast cells exposed to ionizing radiation
A comparative study of quantitative regularities of haploid and diploid yeast cell dying in unnu-trient media was carried out for various strains exposed to ionizing radiation. Cell dying was observed in control and cells treated to chronical (γ-rays of 137Cs) and acute (γ-rays of 60Co) irradiation. It was established that after chronical (for a month at 30 0C) treatment of ionizing radiation at the dose rate range 3-10-2 - 102 of the natural radiation background resulted in the reduction of the cell dying velocity. In these experiments the reverse dose rate effect was obtained - the lower doses and dose rates of ionizing radiation were more effective in comparison with the higher doses and dose rates. The single acute treatment at room temperature resulted in the reduction of dying of cells surviving after irradiation. The cell dying has taken place at 37 и 40 0C. This process was registered not only at low doses decreasing cell survival to 70-90 % but also at high doses when cell survival was only a few percents. The radiation hormesis effect was shown to be slightly expressed for haploid yeast cells. The results obtained in this paper are interpreted within the traditional hypothesis about the role of reparation systems in hormesis effects.
Текст научной статьи Проявление эффекта гормезиса у дрожжевых клеток, облученных ионизирующим излучением
Изучение закономерностей биологических эффектов малых доз ионизирующих излучений становится все более актуальным в связи с облучением больших популяций людей и биосферы во все возрастающих дозах ионизирующего излучения. В литературе отсутствует целостное представление о механизмах запуска и реализации различных эффектов малых доз. Поэтому решение задач, связанных с выявлением закономерностей проявления биологических эффектов малых доз, является перспективным направлением в современной радиобиологии. Среди эффектов малых доз в последнее время особенное внимание уделяется феномену радиационного гормезиса. Термин «гормезис» принят для обозначения положительного действия малых доз ионизирующего излучения [1]. Гормезисные эффекты при действии ионизирующего излучения известны давно и их наблюдали как для простейших клеточных систем [1], так и для животных [2] и человека [3]. Одним из проявлений гормезиса является снижение скорости отмирания облученных клеток по сравнению с интактными [4]. Изменения в живых системах, зависящие от времени, многие ученые определяют как старение, которое сопровождается утратой или гибелью клеток [5, 6]. Существует несколько теорий старения клеток, самой распространенной из которых является теория тепловых повреждений и структурной неустойчивости клеток [5]. Согласно этой теории, живая клетка, подвергаясь воздействию различных факторов окружающей среды, претерпевает разрушительные изменения. Эти изменения варьируют от мельчайших изменений мембран и пластинок до глубоких изменений в генетической структуре клеток. Ранее были проведены эксперименты по влиянию рентгеновского облучения Drosophila melanogaster на продолжительность их жизни [5]. Было обнаружено, что облученные дрозофилы жили доль- ше контрольных. Аналогичные результаты были получены при облучении Campanulara flexuosa [5]. Понятие «старение клеток в культурах» появилось недавно и часто берется в кавычки, т.к. не всегда ясны механизмы старения клеток [7]. Опубликовано сравнительно мало данных, демонстрирующих благоприятное действие ионизирующего излучения для бактериальных [8] и дрожжевых клеток [9] различного генотипа, детерминирующего их способность к восстановлению от повреждений, индуцированных ионизирующим излучением. В связи с этим представляло интерес изучить закономерности влияния хронического и острого действия ионизирующего излучения на процессы старения и отмирания дрожжевых клеток, различающихся по способности к пострадиационному восстановлению.
Материалы и методы
В экспериментах использованы гаплоидные (штамм S288C) и диплоидные дрожжевые клетки ( Saccharomyces cerevisiae ) дикого типа (штамм T1, RAD/RAD ) и радиочувствительный мутант (штамм XS1898, rad52/rad52 ), выращенные на полной питательной среде до стационарной стадии роста в течение 5 суток при температуре 30 ºС. Для изучения влияния мощности дозы на старение и отмирание клеток использовали образцы клеточных суспензий, находящихся в непитательной среде в концентрациях, значительно превышающих максимально возможную концентрацию клеток для данных условий культивирования (106 клеток/мл). Хроническое облучение (137Cs) происходило при разных мощностях доз при 30 °С. 30-кратное снижение мощности дозы по сравнению с интенсивностью естественного радиационного фона (ЕРФ) обеспечивали специальной камерой, выполненной из свинца, выплавленного в 19 веке до открытия радиоактивности. Для острого облучения клеток использовали γ-кванты 60Со (установка «Исследователь») при мощности дозы 40 Гр/мин. Облучаемая суспензия не содержала комков клеток и состояла из одиночных клеток примерно одинакового размера, число почкующихся клеток в ней не превышало 1-2 %. Соответствующие разведения как контрольных, так и облученных клеток высевали на питательную среду. Выживаемость клеток определяли путем подсчета видимых невооруженным глазом макроколоний в чашках Петри после 5-суточной инкубации при 30 ºС на твердой питательной среде. Затем как облученную, так и контрольную суспензии выдерживали в термостате при 37 или 40 ºС. Каждый опыт повторяли 3-5 раз. Результаты измерений обрабатывали и оценивали с применением методов вариационной стати стики.
Результаты и обсуждение
На рис. 1 приведена кривая зависимости выживаемости диплоидных дрожжей S. cere-visiae, штамм XS1898, инкубированных в непитательной среде в течение одного месяца, от мощности дозы хронического облучения, нормированной к интенсивности редко ионизирующего компонента ЕРФ (0,1 мкГр/ч). Как видно из рисунка, самая низкая выживаемость дрожжевых клеток была зарегистрирована при их отмирании в условиях 30-кратного снижения ЕРФ. С увеличением мощности дозы хронического облучения выживаемость клеток возрастает, т.е. старение и отмирание клеток замедляется при повышении интенсивности ионизирующего излучения в диапазоне от 3-10-2 до 100 ЕРФ. Эти данные демонстрируют наличие т.н. обратного эффекта мощности дозы. Данные, приведенные на рис. 1, подтверждают точку зрения о функциональной значимости ЕРФ. В течение миллионов лет эволюция биосферы происходила в условиях повышенных уровней воздействия различных природных химических и физических факторов окружающей среды [10]. Многие физические факторы, включая силу тяжести, давление атмосферы, магнитное поле Земли, естественные потоки корпускулярных заряженных частиц, ионизирующих и неионизирующих электромагнитных излучений, постоянно воздействуют на биоту. Биосфера адаптировалась к действию естественных уровней этих физических и многих химических агентов. Известно, что в процессе абиотической и биотической эволюции интенсивность ЕРФ ионизирующих излучений претерпевала значительные изменения как в пространстве, так и во времени [11]. Значимость ЕРФ в возникновении, эволюции и существовании живых систем детально обсуждается в работах А.М. Кузина [2, 12]. В этих публикациях показано, что снижение ЕРФ может оказывать неблагоприятное действие на скорость деления клеток, рост и развитие организмов. Наоборот, облучение многих биологических объектов при мощностях доз, превышающих ЕРФ в 10-100 и даже в 1000 раз, оказывало благоприятное действие на многие характеристики живых систем. Было высказано предположение [12], что ЕРФ может использоваться клетками для запуска процессов восстановления клеток от повреждений, индуцируемых различными факторами, включая ионизирующее излучение. Эта идея может быть применена для интерпретации обратного эффекта мощности дозы, наблюдавшегося в наших экспериментах (рис. 1). Можно предположить существование некоторой минимальной дозы для запуска систем репарации клеток, которые могут участвовать в ликвидации повреждений, ответственных за старение и отмирание клеток. Тогда повышение мощности дозы в диапазоне 3·10–2 – 102 ЕРФ может приводить к постепенному повышению поглощенной дозы и все более полному запуску систем восстановления, поддерживающих надежность функционирования клеток [13] и приводящих к соответствующему снижению скорости их отмирания.
ЕРФ
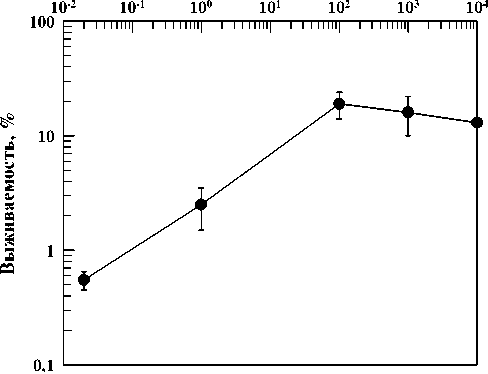
Рис . 1. Кривая зависимости выживаемости диплоидных дрожжей Saccharomyces cerevisiae , штамм XS1898, инкубированных в непитательной среде в течение одного месяца, от мощности дозы хронического облучения, нормированной к интенсивности редкоионизирующего компонента естественного радиационного фона (ЕРФ).
Микродозиметрические оценки частоты прохождения треков через объем облучаемых клеток, основанные на результатах работы [14], позволили нам приближенно оценить потоки частиц через дрожжевые клетки, облучавшиеся в течение одного месяца при различных мощностях доз. Оказалось, что при мощностях доз 3⋅10–2 – 102 ЕРФ накопленная за месяц облучения доза соответствует прохождению значительно менее одного трека электрона через ядра дрожжевых клеток. Это означает, что при наименьшей мощности дозы только через одно из тысячи клеточных ядер хронически облучаемой популяции проходит трек ионизирующей частицы. Следовательно, большая часть популяции дрожжевых клеток каким-то образом получает информацию о необходимости замедления скорости отмирания клеток с увеличением мощности дозы в пределах диапазона 3⋅10–2 – 102 ЕРФ. Следовательно, эти результаты демонстрируют наличие «эффекта свидетеля».
Парадоксальность явлений гормезиса и «эффекта свидетеля», регистрируемых при сверхнизких мощностях доз ионизирующего излучения, состоит в том, что вероятность прохождения треков электронов, формируемых при взаимодействии γ-квантов с веществом, через каждую клетку, а тем более ее чувствительную мишень, ничтожно мала. Было высказано предположение [15], что явление гормезиса может быть связано со свечением Вавилова-Черенкова, большая часть которого излучается в объеме облучаемой среды в виде квантов УФ-света, которые более равномерно распределены по объему облучаемой суспензии из-за большей проникающей способности квантов УФ-света [8] по сравнению с треками электронов, ответственных за ионизацию.
В связи с полученными данными, демонстрирующими большую гибель дрожжевых клеток в условиях хронического воздействия низкими интенсивностями ионизирующего излучения, представляло интерес изучить динамику отмирания клеток после их острого облучения в различных дозах.
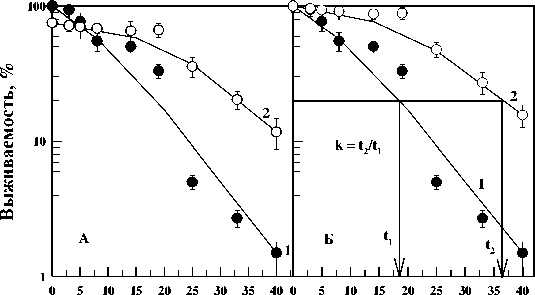
На рис. 2А показана динамика отмирания контрольных (кривая 1) и облученных (кривая 2) в дозе 130 Гр диплоидных дрожжевых клеток Saccharomyces cerevisiae дикого типа (штамм Т1) при температуре инкубации 37 ºС. Видно, что отмирание выживших после острого облучения дрожжей в дозе, снижающей выживаемость клеток до 75 %, приводило к значительному замедлению скорости отмирания выживших после облучения клеток по сравнению с необлученным контролем. По аналогии с фактором изменения дозы можно ввести коэффициент замедления отмирания клеток (k), определяемого отношением продолжительностей отмирания облученных и контрольных клеток, рассчитанного для некоторого изоэффективного уровня (например, для выживаемости клеток, равной 10 %). С этой целью выжившую часть популяции нормировали к 100 %. Данные приведены на рис. 2Б. Такое представление данных будет использовано и при дальнейшем описании полученных результатов. Для данных, представленных на рис. 2, коэффициент замедления отмирания k=1,9 для 20 % уровня выживаемости. Пример нахождения этого коэффициента приведен на рис. 2Б стрелками.
Представляет интерес сравнить динамику отмирания контрольных клеток и клеток, выживших после облучения в еще больших дозах. На рис. 3 и 4 приведены кривые отмирания контрольных дрожжевых клеток и клеток, облученных в дозах 670 и 1200 Гр, снижающих выживае- мость клеток примерно до 20 (рис. 3) и 5 % (рис. 4). Из этих рисунков также видно, что выжившая после облученная популяция дрожжевых клеток отмирает медленнее контрольной. Рассчитанные на основе этих данных коэффициенты замедления отмирания клеток k=1,7 (рис. 3) и k=2,3 (рис. 4) для 10 % уровня выживаемости. Поскольку никакого закономерного изменения этого параметра с дозой не отмечено, не исключено, что полученные значения отражают статистическую вариабельность коэффициента замедления отмирания. Усредняя все полученные значения этого коэффициента (1,9; 1,7 и 2,3), получаем, что k=2,0±0,2. Это означает, что скорость отмирания выживших после облучения клеток не зависела от дозы облучения и была в 2 раза медленнее скорости отмирания контрольных клеток. Эти данные показывают, что горме-зисные эффекты дрожжевых клеток, проявляющиеся в замедлении отмирания выживших после облучения клеток могут проявляться не только в области малых доз как это традиционно предполагало
Продолжительность оI мирания, сутки
Рис . 2. Отмирание клеточной популяции диплоидных дрожжей Saccharomyces cerevisiae (штамм Т1) в непитательной среде при 37 °С без предварительного облучения (кривые 1) и после однократного острого облучения γ-квантами 60Со в дозе 130 Гр (кривые 2).
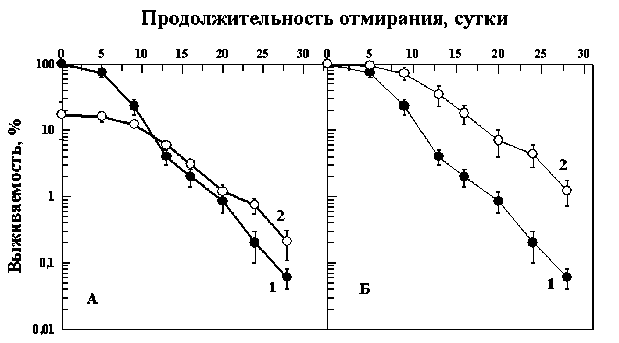
Рис . 3. Отмирание клеточной популяции диплоидных дрожжей Saccharomyces cerevisiae (штамм Т1) в непитательной среде при 37 °С без предварительного облучения (кривые 1) и после однократного острого облучения γ-квантами 60Со в дозе 670 Гр (кривые 2).
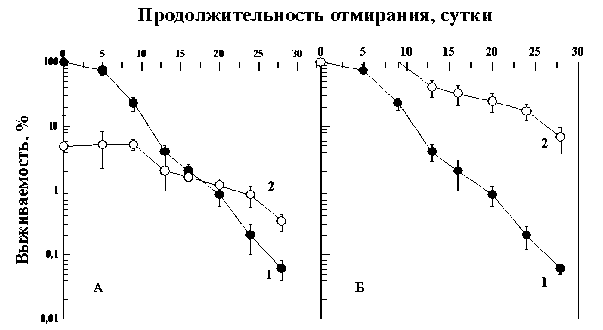
Рис . 4. Отмирание клеточной популяции диплоидных дрожжей Saccharomyces cerevisiae (штамм Т1) в непитательной среде при 37 °С без предварительного облучения (кривые 1) и после однократного острого облучения γ-квантами 60Со в дозе 1200 Гр (кривые 2).
На рис. 5 приведены данные, отражающие динамику отмирания дрожжевых клеток (штамм Т1) при инкубировании их при 40 ºС. Из рисунка видно, что скорость отмирания при повышенной температуре культивирования была больше, чем при более низкой температуре инкубирования (37 °С). При продолжительности инкубирования клеток более 15 суток коэффициент замедления отмирания (k) составлял 1,5±0,2. Это означает, что и при повышенной температуре инкубации эффекты гормезиса наблюдаются, хотя и выражены в меньшей степени. Известно, что механизм положительного действия радиации связывают с запуском систем восстановления клеток, поддерживающих надежность функционирования клеток [13]. Тогда наблюдаемое снижение эффекта гормезиса при 40 °С могло быть связано с меньшей вероятностью восстановления клеток при этой температуре. Дополнительное подтверждение этому факту получено на гаплоидных дрожжевых клетках, способность которых к восстановлению выражена в значительно меньшей степени. Результаты нашего эксперимента приведены на рис. 6. Расчеты показывают, что коэффициент замедления отмирания клеток составлял всего лишь 1,1. Эти данные подтверждают известную точку зрения, что механизм радиационного гормезиса связан с восстановлением клеток от радиационных повреждений.

Рис . 5. Отмирание клеточной популяции диплоидных дрожжей Saccharomyces cerevisiae (штамм Т1) в непитательной среде при 40 °С без предварительного облучения (кривые 1) и после однократного острого облучения γ-квантами 60Со в дозе 1200 Гр (кривые 2).
Продолжительность отмирания, сутки
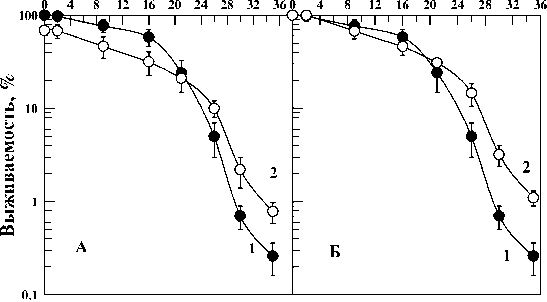
Рис . 6. Отмирание клеточной популяции гаплоидных дрожжей Saccharomyces cerevisiae (штамм S288C) в непитательной среде при 30 °С без предварительного облучения (кривые 1) и после однократного острого облучения γ-квантами 60Со в дозе 130 Гр (кривые 2).
В целом, представленные в данной работе результаты указывают на общебиологическую значимость радиационного гормезиса и поддерживают идею [2, 12] о необходимости пересмотра парадигмы о беспороговом вредном действии ионизирующего излучения.
Список литературы Проявление эффекта гормезиса у дрожжевых клеток, облученных ионизирующим излучением
- Luckey T.D. Hormesis with ionizing radiation. -Florida: CRC Press, 1980. -222 p.
- Кузин A.M. Идеи радиационного гормезиса в атомном веке. -М: Наука, 1995. -156 с.
- Kant К., Chauhan R.P., Sharma G.S. et al. Hormesis in humans exposed to low-level ionising radiation//Int. J. Low Radiation. -2003. -V. 1, N 1. -P. 76-88.
- Петин В.Г., Морозов И.И., Кабакова Н.М. и др. Некоторые эффекты радиационного гормезиса бактериальных и дрожжевых клеток//Радиационная биология. Радиоэкология. -2003. -Т. 43, № 2. -С. 176-178.
- Стрелер Б. Время, клетки и старение. -М.: Мир, 1964. -251 с.
- Clark A.M., Rubin M.A. The modification by X-rays of the life span of haploids and diploids of the wasp, Habrobracon//Radiation Research. -1961. -V. 15, N 2. -P. 244-253.
- Михельсон В.М. Старение клеток в культурах//Надежность и элементарные события старения биологических объектов. -Киев: Наукова Думка. -1986. -С. 116-123.
- Морозов И.И., Петин В.Г., Морозова Г.В. Влияние низкоинтенсивного ионизирующего излучения на процессы размножения, старения и отмирания бактерий Е. coli//Радиационная биология. Радиоэкология. -2002. -Т. 42, № 2. -С. 159-163.
- Петин В.Г., Цепенко В.В. Значимость естественного радиационного фона и малых доз ионизирующего излучения для старения и отмирания дрожжевых клеток//Биосфера и человечество. -Обнинск, 2000. -С. 233-238.
- Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. Краткий очерк теории эволюции. -М.: Наука, 1969.
- Неручев С.Г. Уран и жизнь в истории Земли. -П.: Недра, 1982.
- Кузин A.M. Природный радиоактивный фон и его значение для биосферы Земли. -М.: Наука, 1991.
- Надежность клеток и тканей/Под ред. Гродзинского Д.М. -Киев: Наукова думка, 1980. -211 с.
- Спитковский Д.М. Биологическое действие малых доз ионизирующей радиации//Радиационная биология. Радиоэкология. -1992. -Т. 32, № 3. -С. 382-400.
- Морозов И.И. О биологической роли свечения Вавилова-Черенкова//Радиационная биология. Радиоэкология. -1996. -Т. 36, Вып. 6. -С. 921-925.
