Прооксидантно-антиоксидантная система растений, выращенных из семян, обработанных постоянным магнитным полем

Автор: Шашурин М.М., Журавская А.Н.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 1 т.19, 2023 года.
Бесплатный доступ
Проведено исследование по выявлению физиолого-биохимических изменений в проростках пяти видов растений Якутии, выращенных из семян, получивших предпосевную обработку сверхслабым постоянным магнитным полем (120 мкТл) в течение 24, 72, 96 и 120 часов. Критерием оценки воздействия сверхслабого постоянного магнитного поля служило формирование у сеянцев настоящего листа. В качестве прооксидантной реакции регистрировали интенсивность процессов перекисного окисления липидов, общее содержание низкомолекулярных антиоксидантов - на примере неферментативной антиоксидантной системы и активность антиоксидантного фермента - супероксиддисмутазы. Влияние сверхслабого постоянного магнитного поля оценивали методом коэффициентов. Результаты обсуждены на основе рабочей гипотезы о том, что формирование настоящего листа зависит от изменений прооксидантно-антиоксидантного баланса в клетках проростков, происходящих под действием сверхслабого постоянного магнитного поля. Результаты показали, что у проростков, выращенных из семян, подвергшихся предпосевной обработке сверхслабым магнитным полем индукцией 120 мкТл в течение до 120 часов, процессы перекисного окисления липидов активируются и зависят от режима адаптации антиоксидантных систем: простая компенсация или гиперкомпенсация, что приводит к увеличению приживаемости сеянцев.
Адаптация, проростки растений, прооксиданты и антиоксиданты, сверхслабое постоянное магнитное поле
Короткий адрес: https://sciup.org/143179378
IDR: 143179378
Prooxidant-antioxidate system of plants grown from seeds treated with a constant magnetic field
A study was conducted to identify physiological and biochemical changes in seedlings of five plant species of Yakutia grown from seeds that received pre-sowing treatment with an ultra-weak constant magnetic field (120 μT) for 24, 72, 96 and 120 hours. The criterion for evaluating the impact of an ultra-weak constant magnetic field was the formation of a true leaf in seedlings. As a pro-oxidant reaction, the intensity of lipid peroxidation processes was recorded, the total content of low molecular weight antioxidants - as an example of a non-enzymatic antioxidant system and the activity of the antioxidant enzyme - superoxide dismutase. The influence of an ultraweak constant magnetic field was evaluated using the coefficient method. The results were discussed based on the working hypothesis that the formation of a true leaf depends on changes in the prooxidant-antioxidate balance in seedling cells that occur under the action of an ultraweak constant magnetic field. The results showed that in seedlings grown from seeds that underwent presowing treatment with an ultra-weak magnetic field with an induction of 120 μT for up to 120 hours, the processes of lipid peroxidation are activated and depend on the mode of adaptation of antioxidant systems: simple compensation or hypercompensation, which leads to an increase in survival of seedlings.
Текст научной статьи Прооксидантно-антиоксидантная система растений, выращенных из семян, обработанных постоянным магнитным полем
Все организмы на нашей планете подвергаются непрерывному действию сверхмалых физических полей искусственного и естественного происхождения. К настоящему времени в области исследования биологического действия слабых магнитных полей накоплен огромный экспериментальный материал, свидетельствующий о высокой чувствительности живых систем к их воздействию (Temuryants et. al., 1992, Bingi, 2003). В определенных пределах интенсивности эти воздействия необходимы для поддержания нормальной жизнедеятельности организма. Растения и животные, постоянно находясь под воздействием сверхслабого магнитного поля, в ходе эволюции выработали механизмы рецепции информации о состоянии окружающей среды посредством взаимодействия с электромагнитным полем Земли. Известно, что малые дозы радиации и магнитного поля могут заметно менять физиологические и биохимические характеристики живых организмов (Bingi, 2003, Burlakova et. al., 1999). Например, рядом исследователей, показано, что ответная реакция растительных организмов на действие магнитного поля (электромагнитного или постоянного) зависит от его характеристики (индукции, частотного диапазона, времени экспозиции и т.д.) и от исходного физиологического состояния растений (семян). Причём в зависимости от характеристик этих факторов их совокупность способна оказывать как стимулирующее, так и ингибирующее действие на ряд параметров организмов. Ранее было показано, что сверхслабые магнитные поля способны выступать в качестве мягкого стресс-агента, поэтому их действие на растения может сопровождаться активацией перекисного окисления липидов, обусловленной усилением генерации активных форм кислорода и свободных радикалов, а также, в качестве адаптивной реакции, изменением активности ферментов антиоксидантной защиты (Serdyukov, Novitskii, 2013; Shabrangi et. al., 2010; Sleptsov, 2019; Baryshev, Kasyanov, 2002; Serdyukov, Novitskii, 2013) В других работах установлено, что магнитное поле и малые дозы радиации не вызывают никаких изменений у некоторых организмов (Garkavi et. al., 1996). Общими чертами публикаций по данной тематике являются разнообразие, неоднозначность, иногда даже противоречивость полученных данных, которые не дают полной картины исследований в магнитобиологии как науки, изучающей биологические эффекты магнитных полей. Таким образом, если воздействие электромагнитных полей на биологические системы значительной напряженности (приводящие к тепловым эффектам в биосистемах) исследованы к настоящему времени достаточно полно (Bingi, 2003), то слабые воздействия низко интенсивных полей (нетеплового характера) пока изучены недостаточно.
Цель данной работы – выявить физиологические и биохимические изменения семенного потомства ряда видов растений Якутии на разную временную предпосевную экспозицию семян сверхслабым постоянным магнитным полем.
MATERIALS AND METHODS
В качестве тест-объектов использовали семена ольховника кустарникового [ Duschekia fruticosa (Rupr) Pouzar] , тысячелистника обыкновенного ( Achilléa millefólium ), полыни чернобыльника ( Artemísia vulgáris ), пижмы обыкновенной ( Tanacétum vulgáre ) и иван-чая узколистного ( Chamaenérion angustifolium ), широко распространённых в Центральной Якутии.
Семена проращивали в чашках Петри на фильтровальной бумаге (d = 110 мм) в климатической камере BINDER KMF (Германия) при температуре 25°С, с длиной светового дня 16 часов, при относительной влажности 60%. Опыт закладывали в четырех повторностях по 50 семян в каждой чашке для каждого варианта. Эксперимент проводили в 4-х вариантах: I - сухие семена, прошедшие предпосевную обработку постоянным магнитным полем (ПМП) с индукцией 120 мкТл в течение 24 часов; II – сухие семена - 72 часа; III-сухие семена - 96 часов и IV вариант – 120 часов при тех же условиях. Контроль – семена не прошедшие предпосевную обработку ПМП. Для создания ПМП использовали катушки Гельмгольца, описанные в работе (Sleptsov et. al., 2019).
Критерием оценки воздействия ПМП было образование настоящего листа (выживаемость) у проростков на 21 день наблюдения (%). Появление настоящего листа у проростков говорит о начале активного функционирования апикальной и корневой меристемы – критических тканей растения, которое позволяет предсказать будущую судьбу проростка: возможные отдаленные последствия или его гибель.
В качестве прооксидантной реакции регистрировали интенсивность процессов перекисного окисления липидов (ПОЛ), определяемую по методике, основанной на реакции между образующимся в этом процессе малоновым диальдегидом (МДА, мкмоль/г пр. ) и тиобарбитуровой кислотой (Ermakov, 1987). Суммарное содержание низкомолекулярных антиоксидантов (ƩНМАО, мг-экв. кверц /г пр. ), в качестве неферментативной антиоксидантной системы, определяли по методике, основанной на окрашивании о-фенантролином восстановленного хлорида железа (Lebedeva et. al. , 1977) Определение активности фермента антиоксидантного действия - супероксиддисмутазы (СОД, КФ 1.15.1.1) в проростках проводили по измерению величины молярного коэффициента экстинкции бисформазановых комплексов (Giannopolitis, Ries, 1977).
Оценку влияния ПМП на образование настоящего листа (ОНЛ), интенсивность ПОЛ, активность антиоксидантных систем (как совокупность содержания НМАО и активности СОД) проводили с использованием метода коэффициентов, нормируя значения соответствующих характеристик при различных временах экспозиции ПМП к их значениям в отсутствие ПМП.
Коэффициент образования настоящего листа у проростков вычисляли по формуле:
k ОНЛ = ОНЛ t /ОНЛ о ;
k ПОЛ = (МДА t /МДА о ), отражает в нормированных единицах активность процесса перекисного окисления липидов;
kАОЗ – (НМАОt/НМАОо+СОДt/СОДо)/2, отражает в нормированных к контрольному значению совокупности изменений активности антиоксидантных систем (ƩНМАО+СОД).
Далее, вычисляли коэффициент антиоксидатной защиты, как kАОЗ/ПОЛ = kАОЗ/kПОЛ, который в нормированных единицах, показывает степень адаптации организма к действию ПМП за счет адаптивных изменений активности антиоксидантных систем в ответ на изменения активности прооксидантных систем, то есть от прооксидантно-антиоксидатного равновесия.
Обсуждение результатов по действию на физиологическое состояние проростков растений, выросших из семян, прошедших предпосевную обработку слабым постоянным магнитным полем, проводили исходя из рабочей гипотезы, заключающейся в том, что образование настоящего листа зависит от изменений прооксидантно-антиоксидатного равновесия в клетках проростков, возникающих в результате действия на них ПМП.
Эксперименты проводили в четырех повторностях. Результаты представлены в виде средней арифметической величины. Абсолютную ошибку рассчитывали из среднеквадратической ошибки с помощью коэффициента Стьюдента при р=0.95 (Lakin, 1980). Значимость отличий между средними значениями определяли, используя критерий Ньюмена-Кейлса для множественных сравнений при уровне p≤0.05. Расчет проводили с помощью пакета AnalystSoft, StatPlus – программа статистического анализа, v.2007.
RESULTS AND DISCUSSIONS
В табл.1 представлены физиологические показатели (образование настоящего листа) и характеристики прооксидантно-антиоксидатной системы проростков растений, выросших из семян, получивших предпосевную обработку в течение разной временно’й экспозиции сверхслабым ПМП. Показано, что с увеличением времени нахождения семян (от 24 до 120 часов) в условиях сверхслабого ПМП образование настоящего листа у проростков тысячелистника обыкновенного, пижмы обыкновенной и иван-чая узколистного статистически значимо не отличались от контрольных значений. Количество проростков ольховника кустарникового и полыни чернобыльника увеличилось на 24-28% при возрастании времени нахождения семян в сверхслабом ПМП (табл.1).
Исследование изменений прооксидантно-антиоксидантного статуса у проростков изучаемых растений при действии на семена сверхслабого ПМП (по накоплению малонового диальдегида, активности супероксиддисмутазы и содержанию низкомолекулярных антиоксидантов) показало, что у проростков, выросших из семян, прошедших предпосевную обработку сверхслабым ПМП в течение 96 и 120 часов статистически достоверно: в 1,6-2,8 раза повысилась интенсивность ПОЛ; активировалась СОД, особенно в клетках проростков ольховника кустарникового, полыни чернобыльника и пижмы обыкновенной в 8.8, 5.5. и 4.8 раза, соответственно; повысилось суммарное содержание НМАО в клетках проростков, особенно ольховника кустарникового, тысячелистника обыкновенного, иван-чая узколистного в 3.1, 2.0 и 2.5 раза, соответственно (табл.1 и 2).
Ранее рядом авторов было показано, что действие сверх- и низкоинтенсивных факторов среды (запыление, сверхслабые статические магнитные поля, малые дозы ионизирующего излучения и др.) приводит к увеличению суммы низкомолекулярных антиоксидантов, концентрации малонового диальдегида, активности пероксидазы и супероксиддисмутазы в клетках организмов растений (Sleptsov I.V. et.al, 2019; Prokop’ev I. A. et.al. 2014). Было предположено, что увеличение суммы НМАО, активности пероксидазы и СОД в клетках вызвано ответной реакцией на повышение интенсивности ПОЛ (Sytar O. et.al., 2013; Mittler R. 2002).
Проведенные расчеты коэффициентов позволили построить кривые, показывающие взаимосвязь между количеством проростков с настоящим листом (выживаемостью) и антиоксидантным/прооксидантным соотношением в клетках этих проростков, выросших из семян, получивших разную временну’ю предпосевную обработку сверхслабым ПМП с индукцией 120 мкТл (рис. 1). Установлено, что проростки тысячелистника обыкновенного, пижмы обыкновенной и иван-чай узколистного, выросшие из семян, находившихся в условиях воздействия сверхслабым ПМП в течение свыше 96 часов, сохранили выживаемость по критерию «образование настоящего листа» на уровне контроля, а проростки ольховника кустарникового и полыни чернобыльник увеличили свою выживаемость по этому критерию на 24-28% (таблица 1).
Полученные результаты по изменению антиоксидантно/прооксидантного соотношения (таблица 2) позволяют предположить, что это произошло благодаря тому, что антиоксидантные системы в клетках проростков ольховника кустарникового, полыни чернобыльник, пижмы обыкновенной и иван-чая узколистного адаптировались к повышению активности ПОЛ в 1,62,8 раза при действии сверхслабого ПМП на семена в течение свыше 96 часов в режиме гиперкомпенсации, за счет и повышения концентрации НМАО до 2,5-3,1 раза (иван-чай узколистный, ольховник кустарниковый) и активности СОД до 4,8-7,8 раз (пижма обыкновенная, полынь чернобыльник, ольховник кустарниковый). Соответственно, k АОЗ/ПОЛ при 120 часах экспозиции семян в сверхслабом ПМП составили для проростков ольховника кустарникового – 2.7, пижмы обыкновенной и иван-чай узколистного – 1.4, полыни чернобыльник – 1,2.
Вместе с тем, антиоксидантные системы в клетках проростков тысячелистника обыкновенного адаптировались к повышению активности ПОЛ в 1,82,0 раза при действии сверхслабого ПМП на семена в течение свыше 96 часов в режиме простой компенсации (k АОЗ/ПОЛ составило 0,8), так как содержание НМАО увеличилось всего в 1,7-2,0 раза, а активность СОД не возросла (рис. 1).
Таблица 1. Образование настоящего листа (%), суммарное содержание антиоксидантов, активность СОД и интенсивность процессов перекисного окисления липидов (МДА) у проростков при разном времени воздействии сверхслабым ПМП (120 мкТл) на семена растений (р≤0.1; р≤0.01)
|
Часы |
0 { |
24 { |
72 { |
96 { |
120 |
|
Ольховник кустарниковый |
|||||
|
Настоящий лист, % |
25.0±2.0 |
28.0±2.8 |
30.0±3.0 |
32.0±3.2 |
32.0±3.2 |
|
ƩНМАО, мкг-экв кверц /г проростка |
32.0±0.3 |
35.0±0.5 |
38.0±0.4 |
42.0 ±0.4 |
98.0 ±0.9 |
|
СОД, мкМ/г∙мин |
9.0 ±0.1 |
10.0 ±0.1 |
9.0 ±0.1 |
30.0 ±0.3 |
70.0 ±0.7 |
|
МДА, мкМ/г проростка |
1.5 ±0.01 |
1.7 ±0.02 |
1.5 ±0.01 |
1.8 ±0.02 |
3.0 ±0.03 |
|
Тысячелистник обыкновенный |
|||||
|
Настоящий лист, % |
70.0±7.0 |
72.0±7.2 |
70.0±7.0 |
72.0±7.2 |
70.0±7.0 |
|
ƩНМАО, мкг-экв кверц /г проростка |
19.0±0.2 |
20.0±0.2 |
20.0±0.2 |
32.0±0.3 |
38.0±0.4 |
|
СОД, мкМ/г∙мин |
68.0±0.7 |
68.0±0.7 |
66.0±0.7 |
70.0±0.7 |
72.0±0.7 |
|
МДА, мкМ/г проростка |
2.0±0.02 |
2.4±0.02 |
2.5±0.02 |
3.6±0.04 |
4.0±0.04 |
|
Полынь чернобыльник |
|||||
|
Настоящий лист, % |
45.0±4.5 |
50.0±5.0 |
54.0±5.4 |
54.0±5.4 |
56.0±5.6 |
|
ƩНМАО, мкг-экв кверц /г проростка |
50.0±0.5 |
52.0±0.5 |
55.0±0.5 |
56.0±0.6 |
56.0±0.6 |
|
СОД, мкМ/г∙мин |
6.5±0.06 |
7.8±0.08 |
8.2±0.08 |
20.0±0.2 |
36.0±0.4 |
|
МДА, мкМ/г проростка |
10.0±0.1 |
12.0±0.1 |
12.0±0.1 |
22.0±0.2 |
28.0±0.3 |
|
Пижма обыкновенная |
|||||
|
Настоящий лист, % |
45.0±4.5 |
46.0±4.6 |
45.0±4.5 |
44.0±4.4 |
48.0±4.8 |
|
ƩНМАО, мкг-экв кверц /г проростка |
40.0±0.4 |
42.0±0.4 |
42.0±0.4 |
44.0±0.4 |
50.0±0.5 |
|
СОД, мкМ/г∙мин |
10.0±0.1 |
12.0±0.1 |
12.0±0.1 |
34.0±0.3 |
48.0±0.5 |
|
МДА, мкМ/г проростка |
15.0±0.2 |
16.0±0.2 |
16.0±0.2 |
28.0±0.3 |
32.0±0.3 |
|
Иван-чай узколистный |
|||||
|
Настоящий лист, % |
56.0±5.6 |
66.0±6.6 |
60.0±6.0 |
62.0±6.2 |
60.0±6.0 |
|
ƩНМАО, мкг-экв кверц /г проростка |
96.0±1.0 |
95.0±1.0 |
98.0±1.0 |
98.0±1.0 |
240.0±2.4 |
|
СОД, мкМ/г∙мин |
86.0±0.9 |
86.0±0.9 |
88.0±0.9 |
120.0±1.2 |
160.0±1.6 |
|
МДА, мкМ/г проростка |
1.5±0.01 |
1.7±0.02 |
1.5±0.02 |
1.6±0.02 |
2.4±0.02 |
Таблица 2. Расчетные значения (коэффициенты) образования настоящего листа, суммарного содержания антиоксидантов, активности СОД и интенсивности перекисного окисления липидов у проростков при разном времени воздействии сверхслабым ПМП (120 мкТл) на семена растений (р≤0.1; р≤0.01)
|
Часы |
0 { |
24 { |
72 { |
96 { |
120 |
|
Ольховник кустарниковый |
|||||
|
k ОНЛ |
1 |
1.12 |
1.20 |
1.28 |
1.28 |
|
k НМАО |
1 |
1.09 |
1.90 |
1.31 |
3.06 |
|
k СОД , |
1 |
1.11 |
1.00 |
3.33 |
7.78 |
|
k ПОЛ |
1 |
1.13 |
1.00 |
1.20 |
2.00 |
|
k АОЗ/ПОЛ |
1 |
0.97 |
1.10 |
1.93 |
2.71 |
|
Тысячелистник обыкновенный |
|||||
|
k ОНЛ |
1 |
1.03 |
1.0 |
1.03 |
1.00 |
|
k НМАО |
1 |
1.05 |
1.05 |
1.68 |
2.00 |
|
k СОД , |
1 |
1.00 |
0.97 |
1.03 |
1.06 |
|
k ПОЛ |
1 |
1.20 |
1.25 |
1.80 |
2.00 |
|
k АОЗ/ПОЛ |
1 |
0.85 |
0.81 |
0.75 |
0.77 |
|
Полынь чернобыльник |
|||||
|
k ОНЛ |
1 |
1.11 |
1,20 |
1.20 |
1.24 |
|
k НМАО |
1 |
1.04 |
1.10 |
1.12 |
1.12 |
|
k СОД , |
1 |
1.20 |
1.26 |
3.08 |
5.54 |
|
k ПОЛ |
1 |
1.20 |
1.20 |
2.20 |
2.80 |
|
k АОЗ/ПОЛ |
1 |
0.93 |
0.98 |
0.95 |
1.19 |
|
Пижма обыкновенная |
|||||
|
k ОНЛ |
1 |
1.02 |
1.00 |
0.98 |
1.07 |
|
k НМАО |
1 |
1.05 |
1.05 |
1.10 |
1.25 |
|
k СОД , |
1 |
1.20 |
1.20 |
3.40 |
4.80 |
|
k ПОЛ |
1 |
1.07 |
1.07 |
1.87 |
2.13 |
|
k АОЗ/ПОЛ |
1 |
1.05 |
1.05 |
1.20 |
1.42 |
|
Иван-чай узколистный |
|||||
|
k ОНЛ |
1 |
1.18 |
1.07 |
1.11 |
1.07 |
|
k НМАО |
1 |
0.99 |
1.02 |
1.02 |
2.50 |
|
k СОД , |
1 |
1.00 |
1.02 |
1.40 |
1.86 |
|
k ПОЛ |
1 |
1.13 |
1.00 |
1.07 |
1.60 |
|
k АОЗ/ПОЛ |
1 |
0.88 |
1.02 |
1.13 |
1.36 |
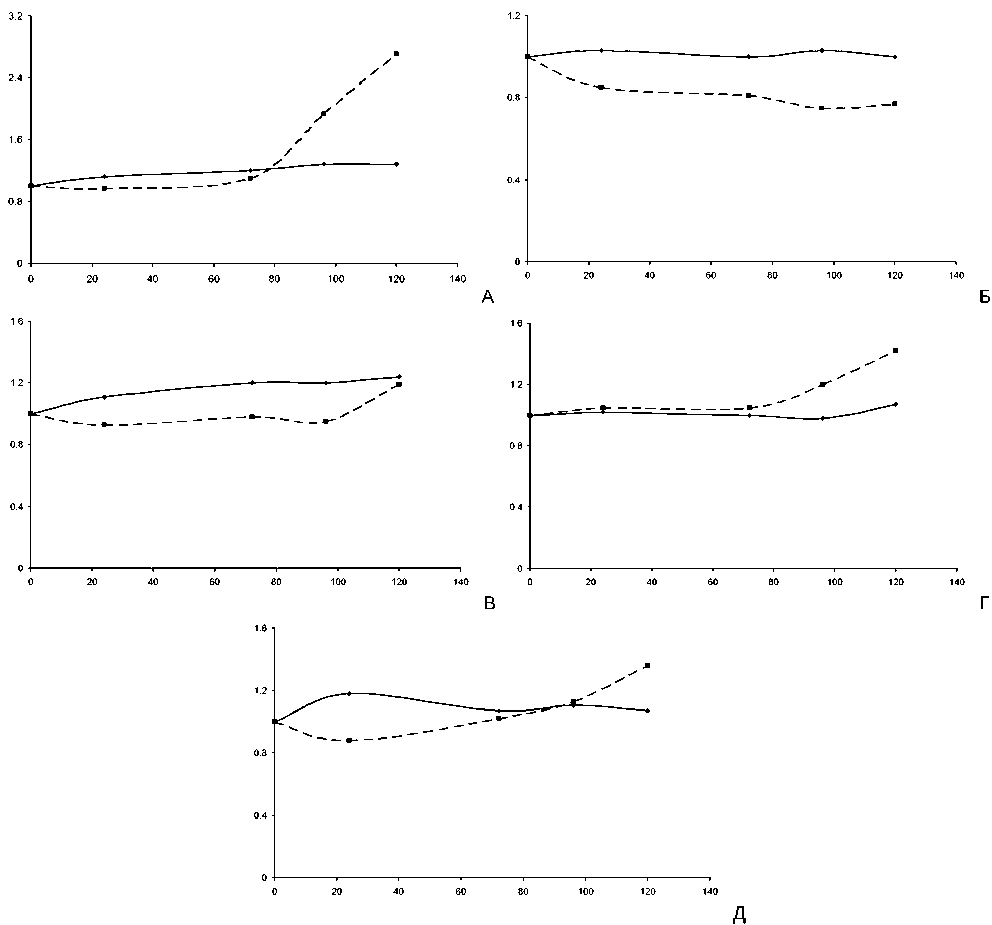
Figure 1. Выживаемость по критерию образования настоящего листа и антиоксидантное/прооксидантное соотношение в клетках проростков, семена которых прошли предпосевную обработку сверхслабым ПМП с индукцией 120 мкТл, в зависимости от времени нахождения семян в условиях сверхслабого ПМП.
А – ольховник кустарниковый;
Б – тысячелистник обыкновенный;
В – полынь чернобыльник;
Г – пижма обыкновенная;
Д – иван-чай узколистный по оси абсцисс – время нахождения семян в условиях ПМП, часы; по оси ординат – значения коэффициентов
—•—настоящий лист —•- Каоз/пол
CONCLUSION прошедших предпосевную обработку сверхслабым магнитным полем с индукцией 120 мкТл в течение до Полученные результаты свидетельствуют о том,
120 часов, которая приводит к активации процессов что для изученных видов дикорастущих травянистых перекисного окисления липидов, зависит от того в растений выживаемость проростков (по критерию каком режиме протекают процессы адаптации образования настоящего листа), выросших из семян, антиоксидантных систем (суммы низкомолекулярных антиоксидантов и активности супероксиддисмутазы): простой компенсации или гиперкомпенсации. Во втором случае действие сверхслабого ПМП приводит к повышению выживаемости по критерию образования настоящего листа.
ACKNOWLEDGMENT
Работа выполнена в рамках проекта «Физиологобиохимические механизмы адаптации растений, животных, человека к условиям Арктики/Субарктики и разработка биопрепаратов на основе природного северного сырья, повышающих эффективность адаптационного процесса и уровень здоровья человека в экстремальных условиях среды», № научной темы 0297-2021-0025, регистрационный номер ЕГИСУ НИОКТР АААА-А21-121012190035-9
CONFLICTS OF INTEREST
The authors declare that they have no potential conflicts of interest.
Список литературы Прооксидантно-антиоксидантная система растений, выращенных из семян, обработанных постоянным магнитным полем
- Baryshev M.G., Kasyanov G.I. (2002) Vozdeistvie elektromagnitnykh polei na biokhimicheskie protsessy v semenakh rastenii. Izvestiya vuzov. Pishchevaya tekhnologiya.. (1). 21-23.
- Bingi V. N. (2003) Fizicheskie problemy deistviya slabykh magnitnykh polei na biologicheskie sistemy // Uspekhi fizicheskikh nauk. 173(3). 265-300.
- Burlakova E.B., Konradov A.A., Goloshchapov A.N., Zhizhina G.P. (1999) Novye aspekty zakonomernostei deistviya nizkointensivnogo oblucheniya v malykh dozakh //Radiatsionnaya biologiya. Radioekologiya. 39(1) 26-34.
- Ermakov A.I. (1987) Metody biokhimicheskogo issledovaniya rastenii. L.: Agropromizdat,. 430 p.
- Garkavi JI.X., Kvakina E.B., Shikhlyarova A.I., Kuzmenko T.S., Barsukova L.P., Maryanovskaya G.Ya., Sheiko E.A., Evstratova O.F., Zhukova G.V. (1996) Magnitnye polya, adaptatsionnye reaktsii I samoorganizatsiya zhivykh sistem. Biofizika, 41(4). 898-905.
- Giannopolitis C.N., Ries S.K. (1977) Superoxide Dismutases: I. Occurrence in Higher Plants Plant Physiol.. V. 59. P. 309–314.
- Lakin G.F. (1980) Biometriya. M.: Vysshaya shkola.. 293 p.
- Lebedeva O.V., Ugarova N.N., Berezin I.V. (1977) Kineticheskoe izuchenie reaktsii okisleniya odianizidina perekisyu vodoroda v prisutstvii peroksidazy iz khrena Biokhimiya. 42. 1372–1379.
- Mittler R. (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 7. 405–410.
- Prokop’ev I. A., Filippova G. V., Shein A. A., Gabyshev D. V. (2014) Impact of urban anthropogenic pollution on seed production, morphological and biochemical characteristics of chamomile, Matricaria chamomila L Russian journal of ecology. 45(1). 18-23.
- Serdyukov Yu.A., Novitskii Yu.I. (2013) Deistvie slabogo postoyannogo magnitnogo polya na aktivnost antioksidantnykh fermentov u prorostkov redisa Fiziologiya rastenii. 60. 66-75.
- Serdyukov Yu.A., Novitskii Yu.I. (2013) Deistvie slabogo postoyannogo magnitnogo polya na aktivnost antioksidantnykh fermentov u prorostkov redisa Fiziologiya rastenii. 60. 66-75.
- Shabrangi A., Majd A., Sheidai M., Nabyouni M., Dorranian D. (2010) Effects of Extremely Low Frequency Electromagnetic Fields on the Antioxidant Enzymes Activity of C3 and C4 Plants Progress In Electromagnetics Research Symposium Proceedings, Cambridge, USA,. 1083–1087.
- Sleptsov I.V. Shashurin M.M., Zhuravskaya A.N. (2019) Kratkovremennoe vozdeistvie postoyannogo magnitnogo polya na fiziologicheskie, morfologicheskie i biokhimicheskie kharakteristiki prorostkov Amaranthus retroflexus, Agastache rugosa i Thlaspi arvense Fiziologiya rastenii. 66(1). 66-72.
- Sytar O., Kumar A., Latowski D., Kuczynska P., Strzałka K., Prasad M. N. V. (2013) Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol. Plant. 35. 985–999.
- Temuryants N.A., Vladimirskii B.M., Tishkin O.G. (1992) Sverkhnizkochastotnye elektromagnitnye signaly v biologicheskom mire. Kiev:Naukova Dumka, 187 p.
