Проростки паразитного растения Cuscuta japonica как модель для изучения апоптоза

Автор: Леусова Н.Ю.
Журнал: Международный журнал гуманитарных и естественных наук @intjournal
Рубрика: Биологические науки
Статья в выпуске: 12-2 (75), 2022 года.
Бесплатный доступ
По-нашему мнению, проросток паразитного растения повилики является привлекательной моделью для изучения молекулярных механизмов ростовых процессов у растений. В статье приводятся данные цитологического и биохимического исследования проростков повилики. Сделан вывод, что отмирание базального полюса, происходит по пути апоптоза, поскольку наблюдались его типичные признаки - уменьшение в объеме клетки в целом, самого протопласта, появление замкнутых везикул, фрагментирование ДНК. Сведения, полученные нами, свидетельствуют в пользу участия активных форм кислорода (АФК)-в частности О2-- как в процессах пролиферации, так и в запрограммированной гибели клеток базального полюса проростка повилики японской. Результаты позволяют предложить новую модель для исследования ростовых процессов и естественного неиндуцированного метаболизма апоптозной растительной клетки.
Проросток повилики cuscuta japonica choisy, ростовые процессы, активные формы кислорода (афк), апоптоз
Короткий адрес: https://sciup.org/170197010
IDR: 170197010 | DOI: 10.24412/2500-1000-2022-12-2-9-12
Seedlings of a parasite plant Cuscuta japonica as a model for studying apoptosis
In our opinion, the seedling of the parasitic dodder plant is an attractive model for studying the molecular mechanisms of growth processes in plants. The article presents the data of cytological and biochemical studies of dodder seedlings. It was concluded that the death of the basal pole occurs along the path of apoptosis, since its typical signs were observed - a decrease in the volume of the cell as a whole, the protoplast itself, the appearance of closed vesicles, DNA fragmentation. The data obtained by us testify in favor of the participation of reactive oxygen species - in particular, O2 - both in the processes of proliferation and in the programmed cell death of the basal pole of the Japanese dodder seedling. The results allow us to propose a new model for the study of growth processes and natural non-induced metabolism of apoptotic plant cells.
Текст научной статьи Проростки паразитного растения Cuscuta japonica как модель для изучения апоптоза
Повилики (Cuscuta L.) относят к группе лишенных фотосинтеза паразитных покрытосеменных растений, полностью питающихся за счет растения – хозяина. В процессе эволюции выработавшийся механизм приспособления роста и развития этих стеблевых паразитов к изменениям окружающей среды отличается и от фото-трофных растений, и от других паразитных растений, большинство из которых являются корневыми паразитами.
По-нашему мнению, проросток паразитного растения повилики является привлекательной моделью для изучения молекулярных механизмов ростовых процессов у растений, поскольку проростки других растений имеют точки ветвления, так что трудно в достаточных количествах выделить ткани с зонами деления, растяжения, зрелые. Большой популярностью для исследования ростовых процессов пользуются колеоптили и корни злаков. Проростки повилики имеют преимущество перед этими моделями – наличие однозначной полярности (униполярности). Рост происходит за счет верхушки проростка – апикальной меристемы, по мере удлинения проростка питательные вещества передви- гаются из нижней части в верхнюю, базальная часть отмирает, но достаточно длительное время (до 3 недель) сохраняется постоянная длина живого стебля проростка повилики.
Апоптоз является обязательной составляющей процесса развития и без него эмбриогенез невозможен [1]. Апоптозная элиминация клеток является необходимым условием и развития растений, в частности, в развивающихся пыльниках ячменя, в эндосперме формирующейся зерновки кукурузы, в алейроновом слое ячменя при прорастании, в колеоптиле и листьях пшеницы и во многих стареющих клетках и тканях растений [2]. Апоптоз у растений сопровождается чередованием структурноморфологических изменений клетки: происходит выраженная конденсация хроматина с последующим распадом ядра, меж-нуклеосомальной фрагментацией ядерной ДНК, в клетках образуются гигантские вакуоли. Тем не менее, сведения об апоптозе у ратений все еще очень фрагментарны [1]. Некоторые авторы даже подвергают сомнению явления апоптоза у растений [3]. Поэтому, на наш взгляд, выбор еще одной модели для исследования явлений апопто- за может пополнить картину, происходящую в растительной клетке при этом процессе.
Мы поставили перед собой задачу - выяснить, происходит ли в базальной отмирающей части проростка характерные для апоптозной клетки изменения тканей, и характерна ли для этой части проростка межнуклеосомальная фрагментация ДНК.
Для изучения ростовых процессов использовали сеянцы повилики разного возраста. Проростки выращивали 1-2 недели при комнатной температуре на свету. При исследовании клеток разных зон проростка мы предпочли метод измельченных препаратов с окрашиванием ацетокармином.
Выделение и электрофорез полученной ДНК проводили согласно [4].
Результаты. Базальный полюс 4-х дневного проростка повилики начинает отмирать: изменяется окраска препарата, – в результате деструктивных процессов содержимое клеток сжимается, происходит отделение участков цитоплазмы с органеллами в вакуоль клетки в виде замкнутых везикул (рис. 1). Ядра красителем не прокрашиваются вследствие разрушения хроматина.
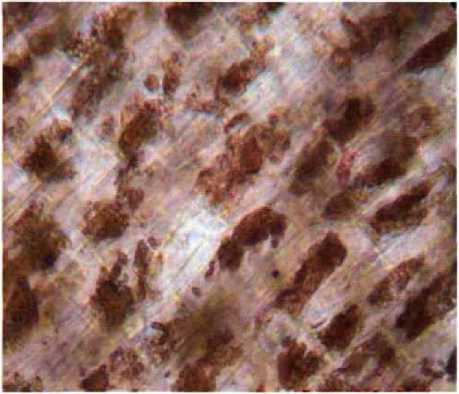
Рис. 1. Базальный полюс проростка повилики японской (х660)
Выделенную ДНК проростков повилики японской подвергали электофорезу. Электрофореграмма ДНК, полученная из проростков повилики показана на рис.
Электрофорез ДНК 4-дневных проростков (рис. 2, а) показал, что верхушка и средняя часть проростка не характеризуются апоптозной фрагментацией ДНК, в отличие от базального полюса, где обнаружены легкие фрагменты (происходит полная фрагментация ДНК). Однако в данном случае мы не обнаружили промежуточного звена - где были бы как тяжелые, так и легкие цепи ДНК. В последую- щем мы разделили 4-дневный проросток повилики на 4 части (рис. 2 (б)-1,2, 3,4). Апоптозная фрагментация ДНК наблюдалась уже в живой части проростка ближе к базальному полюсу (рис. 29, 6-2). У 2 -дневных проростков повилики с не отмирающим еще базальным полюсом наблюдалась следующая картина: фрагментация ядерной ДНК не характерна для верхушки 2-дневного проростка повилики, в базальном полюсе уже отмечается наличие легких фрагментов ДНК (начинается фрагментация ДНК по типу апоптоза).
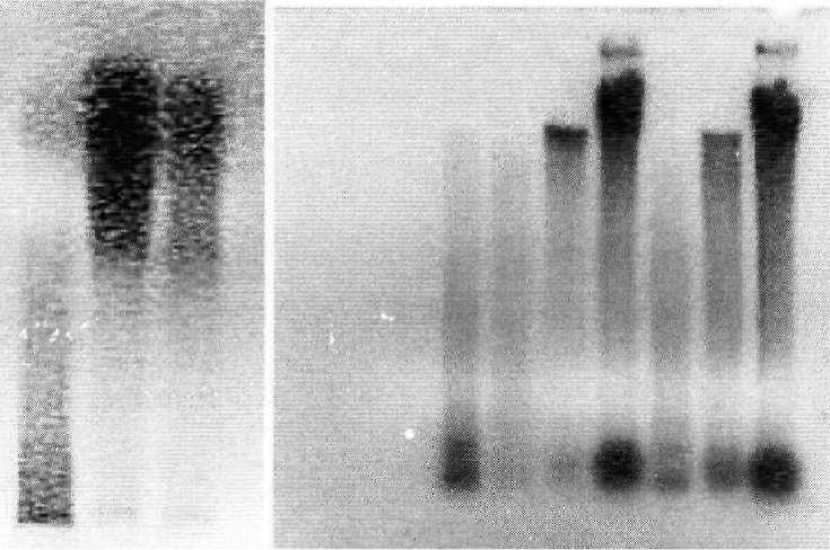
(а) (б)
1 2 3 1234567
Рис. 2. Электрофореграммы ДНК, выделенной из проростков повилики японской
а - ДНК 4-х дневных проростков с отмирающим базальным полюсом: 1- базальный полюс; 2 - средняя часть проростка; 3- верхушка б - ДНК проростков повилики различного возраста: 1- базальный полюс 4 - дневного проростка; 2- средняя часть (ближе к базальному полюсу) 4— дневного проростка; 3- средняя часть 4дневного проростка (ближе к верхушке); 4- верхушка 4- дневного проростка; 5 - базальный полюс 2- дневного проростка; 6-средняя часть 2- дневного проростка; 7-верхушка 2- дневного проростка.
Таким образом, получено еще одно свидетельство в пользу апоптоза базального полюса у проростков повилики японской. В базальном полюсе 4- дневного проростка происходит апоптозная фрагментация ДНК. У 2-дневных проростков начинается фрагментация я-ДНК базального полюса.
На основании проведенных нами цитологических исследований обнаружены изменения клеточных структур в различных участках проростка повилики; и выделены четкие зоны: меристематическая – зона деления, средняя – зона растяжения, и базальная – зона отмирания . Сделан вывод, что отмирание базального полюса, происходит по пути апоптоза, поскольку наблюдались его типичные признаки – уменьшение в объеме клетки в целом, самого протопласта, появление замкнутых везикул, фрагментирование ДНК.
Список литературы Проростки паразитного растения Cuscuta japonica как модель для изучения апоптоза
- Замятнина В.А., Бакеева Л.Е., Александрушкина Н.И., Ванюшин Б.Ф. Апоптоз у этиолированных проростков пшеницы.2. Влияние антиоксиданта (ВНТ) и перекисей // Физиология растений. - 2003. - Т. 50. - С. 280-290.
- Кирнос М.Д., Александрушкина Н.И., Ванюшин Б.Ф. Апоптоз в клетках первого листа и колеоптиля проростков пшеницы: межнуклеосомная фрагментация ДНК и синтез "тяжелых" фрагментов ДНК нуклеосомной длины // Биохимия. - 1997. - Т. 62. - С. 1008-1014.
- Doom W.G., Woltering E.J. Many ways to exit? Cell death categories in plants // Trends in Plant Science. 2005. Vol. 10, № 3. P. 117-122.
- Александрушкина Н.И., Коф Э.М., Середина А.В., Борзов А.А., Ванюшин Б.Ф. Cвязанные со старением деградация ДНК и эндонуклеазная активность в листьях гороха нормального и афильного генотипов // Физиология растений. - 2008. - Т. 55. - С. 27-36.
