Пространственная и демографическая структура ценопопуляций сарсазана шишковатого (Halocnemum strobilaceum (Pall.) Bieb., Chenopodiaceae) в Республике Калмыкии
 Bieb., Chenopodiaceae) в Республике Калмыкии")
Автор: Зенкина Т.Е., Сагалаев В.А.
Журнал: Природные системы и ресурсы @ns-jvolsu
Рубрика: Биология и биотехнология
Статья в выпуске: 1 (3), 2012 года.
Бесплатный доступ
Обсуждаются результаты исследования ценопопуляций сарсазана на солончаках в Кумо-Манычской впадине (Республика Калмыкия). Описывается динамика возрастной и пространственной структуры популяций, характеризуется роль семенного и вегетативного размножения в поддержании численности и структуры популяций.
Ценопопуляции, возрастные (онтогенетические) группы, демографические спектры, пространственная структура, динамика
Короткий адрес: https://sciup.org/14967350
IDR: 14967350 | УДК: 582.661.15(470.47)
Regional and demographical structure of cenopopulations of Halocnemum strobilaceum (Pall.) Bieb. (Chenopodiaceae) in the Republic of Kalmykia
The investigation results of cenopopulations of Halocnemum strobilaceum (Pall.) Bieb. in salt marshs in Kumo-Manychskaya hollow (republic of Kalmykia). The dynamics of age and regional structure of populations are described, the role of seed and vegetative reproduction in keeping the number and structure of the population is characterized.
Текст научной статьи Пространственная и демографическая структура ценопопуляций сарсазана шишковатого (Halocnemum strobilaceum (Pall.) Bieb., Chenopodiaceae) в Республике Калмыкии
Исследования ценотических популяций растений являются основой для разработки научных рекомендаций по сохранению биологического разнообразия и природных экосистем в условиях антропогенного стресса. Изучение организации популяций позволяет оценить их состояние, определить способы самоподдержа-ния и устойчивого существования вида. Знание биологии вида и структуры его популяций – необходимая основа для прогноза дальнейшего развития популяций и реакции растений на факторы среды обитания [2; 3; 5; 10; 11; 13; 14].
В условиях аридного юго-востока Европейской России заметную роль играют гало-фитные сообщества. Одним из доминантов таких группировок является Сарсазан шишковатый [ Halocnemum strobilaceum (Pall.) Bieb.] [6; 8; 12]. В Северном Прикаспии сар-сазан образует характерные сообщества – сарсазанники, приуроченные к мокрым и пухлым солончакам [6; 7], занимающие значительные по площади территории.
В этом регионе производится активная добыча углеводородного сырья, в ходе которой происходит интенсивное антропогенное воздействие на почвенно-растительный покров.
Материалы и методы
Нами была поставлена задача выяснить закономерности восстановления ценопопуля-ций сарсазана как вида-доминанта на солончаках в районах добычи нефти в Кумо-Ма-нычской впадине на территории Черноземель-ского района Республики Калмыкии. H. strobilaceum (Pall.) Bieb. представляет собой эугалофильный полукустарник (кустарничек) 15–25 см в высоту с распростертыми одревесневающими побегами и редуцированными листьями. Исследования проводились в полевые сезоны 2009–2011 гг. по классической методике изучения пространственной и демографической структуры популяций растений [3; 5; 11; 13; 14]. Для оценки горизонтальных проекций особей растений закладывались и картировались площадки 3 х 3 м в ценопопу-ляциях вида, отличающихся по возрастной и пространственной структуре.
Полученные результаты и их обсуждение
Особенности жизненного цикла Сар-сазана шишковатого (H. strobilaceum). Изучение онтоморфогенеза H. strobilaceum [4] позволило установить, что жизненный цикл отдельной особи сарсазана составляет около 8– 10 или более лет. Онтогенетический ряд
H. strobilaceum представлен следующими возрастными фазами (состояниями): проростки ( р ), ювенильные ( j ), имматурные ( im ), вирги-нильные ( v ), молодые генеративные ( g1 ), средневозрастные – старые генеративные ( g2 – g3 ), сенильно-виргинильные ( sv ), сенильно-вирги-нильно-генеративные ( svg ), сенильные ( s ).
Прегенеративные формы у сарсазана хорошо различимы, календарный возраст каждой составляет один год. В состоянии ( g1 ) растение находится в течение двух лет, затем переходит в средневозрастное – старое генеративное состояние ( g2 – g3 ), которое длится около трех лет. Длительность жизни сенильных особей представлена 1–2 годами.
В ходе исследований было обнаружено явление поливариантности онтоморфогенеза сарсазана, которое проявляется в переходе отдельных особей из прегенеративного (вирги-нильного) возрастного состояния в особое се-нильно-виргинильное ( sv ) [4]. Кроме того, на этапе ( g2 – g3 ), помимо закономерного перехода растения в сенильное состояние ( s ), наблюдается появление побегов, габитуально схожих с вегетативными, которые на следующий год могут зацветать (состояние svg ) [там же].
Поливариантность онтоморфогенеза наблюдается в жизненных циклах и других растений [1; 2]. Она рассматривается как важная адаптация ценопопуляций растений к воздействию факторов естественной или антропогенной природы [1; 2].
Переход генеративных особей ( g2 – g3 ) в сенильно-виргинильно-генеративное состояние ( svg ) схематично можно представить следующим образом (рис. 1).
Особи, находящиеся в генеративном состоянии ( g2 – g3 ), на завершающем этапе своего развития могут за счет распада разросшейся куртины на отдельные парцеллы переходить в особое возрастное состояние, которое можно определить как сенильно-виргинильно-генера-тивное ( svg ). При этом парцеллы распавшегося единого куста утрачивают связь между собой и существуют самостоятельно. Они сохраняют ограниченную способность к формированию зрелых семян, хотя процессы отмирания продолжаются. Одновременно такие особи способны формировать вегетативные побеги, характерные для виргинильного состояния ( v ).
Кроме того, в ходе исследований был выявлен механизм перехода виргинильных особей ( v ) сарсазана в особое квазистояние, которое можно охарактеризовать как сениль-но-виргинильное состояние ( sv ). Виргиниль-ные особи, минуя генеративную стадию развития, сразу переходят в своеобразное старческо-подростковое состояние, характеризующееся процессами отмирания части куста при сохранении характерных для виргиниль-ных особей вегетативных побегов. Этот процесс протекает на фоне разрастания и парти-куляции материнской особи.
Схема перехода виргинильных особей ( v ) в сенильно-виргинильное состояние показана на рисунке 2.
Пространственно-демографическая структура ценопопуляций H. strobilaceum . Анализ результатов изучения пространственно-демографической структуры ценопопуляций сарсазана в районе исследования позволил выявить три их условные группы: «молодые», «зрелые» и «стареющие».
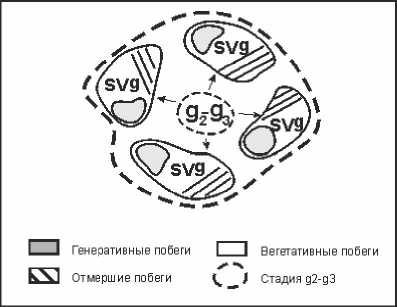
Рис. 1. Схема образования сенильно-виргинильно-генеративных ( svg ) особей H. strobilaceum
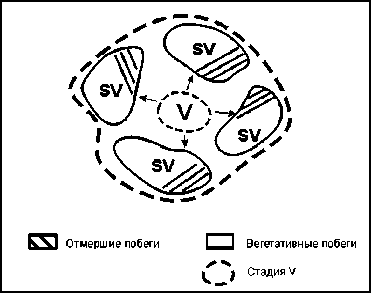
Рис. 2. Схема формирования сенильно-виргинильных ( sv ) особей H. strobilaceum
Горизонтальная проекция участка «молодой» ценопопуляции H. strobilaceum представлена на рисунке 3.
В такой ценопопуляции наблюдается численное господство особей в прегенеративном состоянии. Группировки сарсазана данного типа были зафиксированы по краям мокрых нированными участками рельефа, а также на участках солончаков, где был нарушен почвенно-растительный покров в ходе строительных или других производственных работ. Возрастной спектр такой ценопопуляции показан на рисунке 4. Он является левосторонним, с одним максимумом, приходящимся на стадию

S ■-----► N
g 1 g 2 –g 3 svg s
Сарзан шишковатый (Halocnemum strobilaceum)
Соперос европейский (Salicornia europaea)
Проростки
Молодое генеративное состояние
Средневозрастное – старое генеративное состояние
Сенильно-виргильно-генеративное состояние
Сенильное состояние
Рис. 3. Горизонтальная проекция участка «молодой» ценопопуляции H. strobilaceum
Следующим этапом развития ценопопу-ляции является ее «зрелое» состояние. Подобные ценопопуляции изредка встречаются на окраинах солончаков по соседству с «молодыми». В них представлены растения преимущественно в молодом генеративном
( g1 ) и средневозрастном – старом генеративном состоянии ( g2 – g3 ). Здесь также часто встречаются имматурные ( im ) и виргиниль-ные ( v ) особи. Горизонтальная проекция пространственной структуры такой ценопопуля-ции показана на рисунке 5.
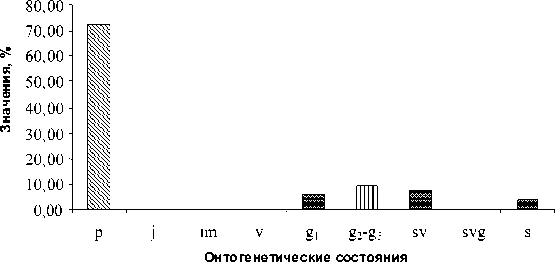
Рис. 4. Возрастной спектр «молодой» популяции H. strobilaceum

Сарсазан шишковатый ( Halocnemum strobilaceum ) Сведа ( Suaeda sp. )
Петросимония ( Petrosimonia sp .)
Бескильница ( Puccinellia sp .)
g1 Молодое генеративное состояние g2–g3 Средневозрастное – старое генеративное состояние
□о
v Виргинильное состояние im Имматурное состояние svg Сенильно-виргинильно-генеративное состояние
Рис. 5. Горизонтальная проекция участка «зрелой» ценопопуляции H. strobilaceum
Возрастной спектр, отражающий демографию зрелой популяции, характеризуется наличием двух максимумов, приходящихся на g1 и g2–g3 состояния (см. рис. 6). Популяции с такой демографической структурой свойственны преимущественно экотонным местообитаниям в верхней или средней части катены. Это связано, по-видимому, с мощным аллелопатическим эффектом как самого сарсазана, так и других галофитных видов – обитателей солончаков [9], а также с тем обстоятельством, что в средней и верхней части катены создаются наиболее благоприятные условия для прорастания семян H. strobilaceum.
Центральную пониженную часть солончаков и влажных депрессий занимают «стареющие» ценопопуляции H. strobilaceum . Особи сарсазана здесь находятся преимущественно в сенильно-виргинильном ( sv ) и се-нильно-виргинильно-генеративном ( svg ) состоянии. Горизонтальная проекция ценопопу-ляции показана на рисунке 7. Такие группировки практически лишены сопутствующих сарсазану на других участках катены галофитов и представлены монодоминантными сообществами, состоящими в основном из особей H. strobilaceum .
В таких ценопопуляциях могут встречаться особи, находящиеся в прегенератив-ной стадии развития ( p, j, im,v ), однако большинство из них отмирают и не достигают генеративной стадии развития по причине высокой плотности расположения особей сарса-зана и высокого уровня их аллепатической активности. Именно в таких ценопопуляциях наблюдается образование сенильно-вирги-нильных ( sv ) и сенильно-виргинильно-генера-тивных ( svg ) особей, описанное выше.
Возрастной спектр «стареющей» цено-популяции представлен на рисунке 8.
Базовый возрастной спектр всех типов це-нопопуляций сарсазана показан на рисунке 9. Он характеризуется сочетанием двух максимумов, приходящихся на проростки ( р ) и средневозрастное – старое генеративное состояние ( g2 – g3 ).
Кроме того, в процессе исследований были выполнены наблюдения за самовосстановлением сарсазанников, происходящим на участках солончаков, которые ранее подвергались уничтожению в процессе хозяйственной деятельности человека. На рисунке 10 представлена динамика изменения возрастной структуры цено-популяции за три года наблюдения (2009– 2011 гг.), прошедшая все три этапа – «молодой», «зрелой» и «стареющей» ценопопуляции.
На графике отражена своеобразная «волна жизни», возникающая в ходе восстановительной сукцессии. На территорию, где в процессе хозяйственной деятельности человека (прокладки нефтепровода в 2006 г.) были уничтожены заросли сарсазана, с соседних участков происходил занос диаспор (семян) и инициировался процесс самозарастания солончаков. На первом этапе наблюдений фиксировалось преобладание в демографическом спектре проростков ( р ) и структура ценопопуляций соответствовала их «молодому» состоянию. На второй год наблюдений ценопопуляции переходили в «зрелое» состояние и в них начинали доминировать имматурные ( im ), виргинильные ( v ) и генеративные ( g ) особи. В 2011 г. ценопопуляции достаточно быстро перешли в «зрело-старческое» состояние и в их возрастном спектре увеличилось число сенильно-виргинильных ( sv ) особей, хотя еще сохранялась значительная доля особей прегенеративной фракции ( j, im, v ).
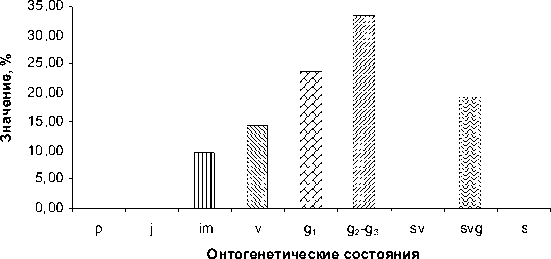
Рис. 6. Возрастной спектр «зрелой» популяции H. strobilaceum
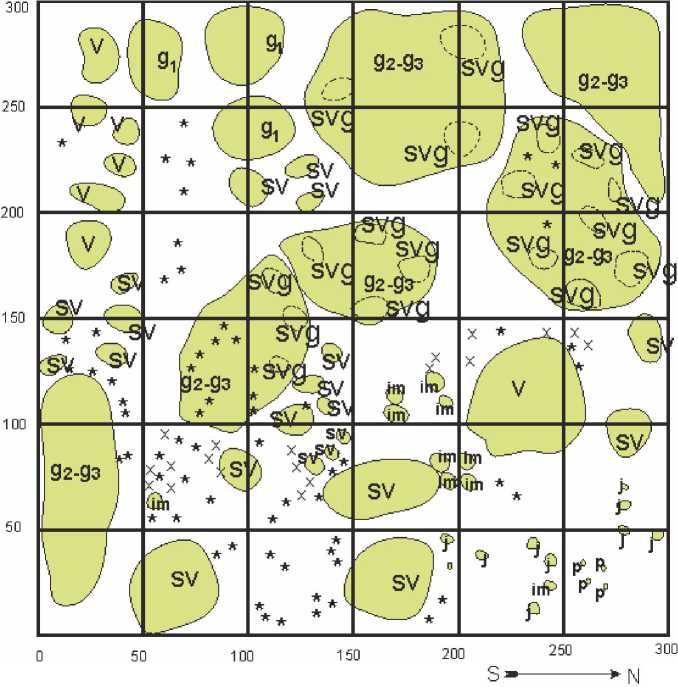
Сарсазан шишковатый (Halocnemum strobilaceum) Отмершие особи в имматурном и виргинильном состояниях х Отмершие проростки p Проростки j Ювенильное состояние im Имматурное состояние v Виргинильное состояние g1 Молодое генеративное состояние g2–g3 Средневозрастное – старое генеративное состояние sv Сенильно-виргинильное состояние svg Сенильно-виргинильно-генеративное состояние s Сенильное состояние
Рис. 7. Горизонтальная проекция участка «стареющей» ценопопуляции H. strobilaceum
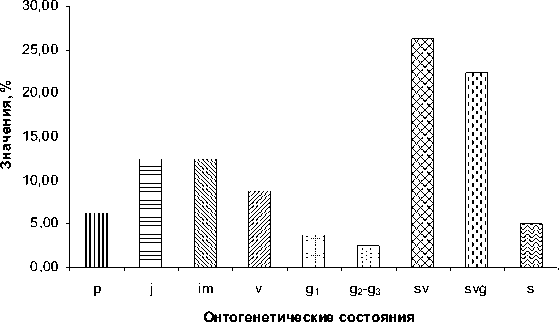
Рис. 8. Возрастной спектр «стареющей» популяции H. strobilaceum
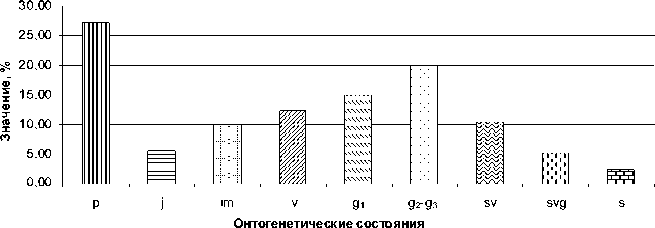
Рис. 9. Базовый онтогенетический спектр популяции H. strobilaceum
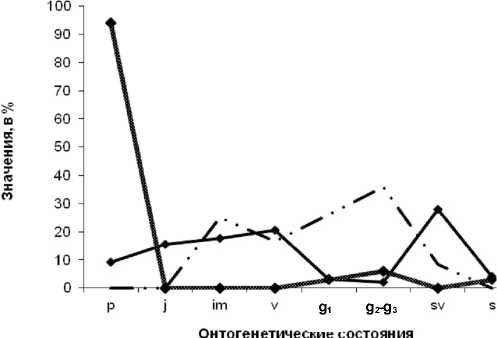
Рис. 10. График изменения возрастной структуры ценопопуляци сарсазана за 3 года наблюдений (2009 г. – сплошная жирная линия; 2010 г. – линия с пунктирами; 2011 г. – сплошная тонкая линия) в ходе восстановительной сукцессии после полного уничтожения сарсазанников
Заключение
Жизненный цикл сарсазана ( H. strobilaceum ), помимо латентной, включает 3 возрастных стадии (прегенеративную, генеративную и сенильную), представленные 9 возрастными состояниями: проростки ( р ), ювенильные ( j ), имматурные ( im ), виргиниль-ные ( v ), молодые генеративные ( g1 ), средневозрастные – старые генеративные ( g2 – g3 ), сенильно-виргинильные ( sv ), сенильно-вирги-нильно-генеративные ( svg ), сенильные ( s ). В онтоморфогенезе растения наблюдается поливариантность развития, связанная с нарушением нормального процесса роста и развития особей сарсазана в условиях монодоминант-ных сообществ. Общая продолжительность жизненного цикла длится 8–10 и более лет.
При изучении пространственно-демографической структуры популяций были выявлены три возрастных этапа – «молодой», «зрелой» и «стареющей» ценопопуляции, каж- дый из которых характеризуется специфической структурой и строением.
Наблюдения за процессами самозарас-тания нарушенных участков солончаков свидетельствуют о достаточно высоком уровне устойчивости сарсазановых сообществ к воздействию антропогенных факторов и их способности к саморегуляции и самоподдержа-нию за счет изменения пространственно-демографической структуры.
Список литературы Пространственная и демографическая структура ценопопуляций сарсазана шишковатого (Halocnemum strobilaceum (Pall.) Bieb., Chenopodiaceae) в Республике Калмыкии
- Жукова, Л. А. Поливариантность онтогенеза и динамика ценопопуляций растений/Л. А. Жукова, А. С. Комаров//Журнал общей биологии. -1990. -Т. 51, № 4. -С. 450-461.
- Жукова, Л. А. Популяционная жизнь луговых растений/Л. А. Жукова. -Йошкар-Ола: Ланар, 1995. -224 с.
- Заугольнова, Л. Б. Структура популяций семенных растений и проблемы их мониторинга: автореф. дис.... д-ра. биол. наук/Заугольнова Людмила Борисовна. -СПб., 1994. -70 с.
- Зенкина, Т. Е. Особенности онтогенеза Сар саза на шишковатого [Hal ocne mum strobilaceum (Pall.) Bieb., Chenopodiaceae] в условиях Нижнего Поволжья/Т. Е. Зенкина//Материалы междунар. науч. конф. (Волгоград, 22-24 апр. 2012). -Волгоград: Изд-во ВолГУ, 2012. -С. 56-58.
- Работнов, Т. А. Вопросы изучения состава популяций для целей фитоценологии/Т. А. Работнов//Проблемы ботаники. -М.; Л.: Изд-во АН СССР, 1950. -Вып. 1.-С. 465-483.
- Растительность европейской части СССР/под ред. С. А. Грибовой, Т. И. Исаченко, Е. М. Лавренко. -Л.: Наука, 1980. -429 с. 7.
- Рухленко, И. А. Конспект сообществ подкласса Kalidienea Golub et al. 2001 (кл. Salicornietea fruticosae) на территории Евразии/И. А. Рухленко//Изв. Самар. науч. центра РАН. -2011. -Т. 13, № 5. -С. 114-121.
- Симагина, Н. О. Фенольные соединения Artemisia santonica L., Halocnemum strobilaceum (Pall.) M. Bieb., проявляющие аллелопатическую активность/Н. О. Симагина, Н. В. Глумова//Уч. зап. Таврического нац. ун-та. Сер. «Биология, химия». -2008. -Т. 21(60), № 2. -С. 113-160.
- Смирнова, О. В. Структура травянистого покрова широколиственных лесов/О. В. Смирнова. -М.: Наука, 1987. -207 с.
- Уранов, А. А. Возрастной спектр фитоценопопуляций как функция времени и энергии волновых процессов/А. А. Уранов//Биологические науки. -1975. -№ 2. -С. 7-34. 12.
- Храмцов, В. Н. Растительность солончаков/В. Н. Храмцов//Ботаническая география Казахстана и Средней Азии (в пределах пустынной области). -СПб.: [б. и.], 2003. -С. 112-118.
- Ценопопуляция растений (основные понятия и структура). -М.: Наука, 1976. -217 с.
- Ценопопуляции растений (очерки популяционной биологии). -М.: Наука, 1988. -184 с.
- Растительный покров СССР: пояснительный текст к геоботанической карте СССР/под ред. Е. М. Лавренко, В. Б. Сочавы. -М.; Л.: Изд-во АН СССР, 1956. -Т. 2. -С. 461-971.
