Пространственная организация ценопопуляций Artemisia keiskeana в дубовых лесах юга Приморского края

Автор: Бисикалова Е.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агролесомелиорация и лесное хозяйство
Статья в выпуске: 10, 2014 года.
Бесплатный доступ
В статье описаны основные изученные нами закономерности пространственной организации це-нопопуляций A. keiskeana в дубовых лесах юга Приморского края. Установлено, что в результате активного вегетативного размножения растения растут взаимосвязанно, образуя куртины, состоящие из молодых побегов. Произрастает в окнах древостоев.
Пространственная организация, ценопопуляция, куртина, побег, онтогенетический период
Короткий адрес: https://sciup.org/14083354
IDR: 14083354 | УДК: 581.55
Spatial organization of Artemisia keiskeana coenopopulations in the oak forests of the Primorsky Krai south
The basic laws of the A. keiskeana coenopopulation spatial organization in the oak forests of the Primorsky Krai south studied by the authors are described in the article. It is established that as a result of the active vegetative reproduction the plants grow interconnected, forming clumps, consisting of young shoots. It grows in the forest stand windows.
Текст научной статьи Пространственная организация ценопопуляций Artemisia keiskeana в дубовых лесах юга Приморского края
Материалы и методы исследования. Для изучения пространственной организации ценопопуляций A. keiskeana была заложена геоботаническая пробная площадь размером 30×30 м в дубовом лесу на юге Приморского края (Горнотаёжная станция ДВО РАН). Первым этапом работы был поиск и ознакомление с пробной площадью. Обозначение пробной площади производилось по периметру маркировочными лентами. Далее разбивали площадь на полосы шириной по 2 метра (всего 15 полос). Каждая полоса разбивалась на квадраты 2×2 метра, которые в свою очередь разбивались на квадраты 0,5×0,5 метров (в сумме 3600 учётных площадок) для более точного сбора данных. Травянистые виды дубравной свиты учитывались на учётных площадках по всей пробной площади. Древесный и кустарниковый ярус учитывался на квадратах 1×1 метр. Учёт травянистых растений проводился по онтогенетическим периодам: первая и вторая фаза прегенеративно-го онтогенетического периода (V1, V2), генеративный онтогенетический период (G) и сенильный онтогенетический период (S) (онтоморфологическая характеристика [Бисикалова, 2013]). Счётная единица – побег.
Впервые при исследовании пространственной структуры ценопопуляций применена методика анализа точечных мозаик [Ухваткина, Омелько, 2011; Омелько, Ухваткина, 2013], основанная на использовании парной корреляционной функции [Wiegand, Moloney, 2004; Wiegand et al., 2007; Грабарник, 2010]. Метод анализа точечных мозаик был привлечён для выявления особенностей размещения растений видов дубравной свиты относительно друг друга и относительно деревьев. Анализы проводились с помощью программного обеспечения Programita . Для тестирования эффектов в данной работе использовались следующие нуль-модели: процесс кластеризации (cluster process); гетерогенный случайный процесс (heterogeneous Possion process); процесс независимого размещения элементов двух мозаик (torroidal shift). При использовании каждая нуль-модель имеет свое значение. Так, с помощью cluster process (CL) было протестировано распределение элементов в кластере (проверено, как расположены точки – случайным образом, разрежено или сгруппировано). Модель heterogeneous Possion process (HP) используется в случае однородности элементов одной мозаики. При использовании toroidal shift (TSH) мы смотрим, совпадает ли размещение групп разных видов или они расположены в разных местах. При этом не сравнивается структура этих мозаик, но учитывается при анализе [Ухваткина, Омелько, 2011]. Анализы пространственной организации проводились в трёх направлениях: 1) однородности размещения растений; 2) однородности размещения растений в онтогенетических периодах; 3) независимости расположения растений относительно деревьев.
Результаты и их обсуждение. Пробная площадь расположена в леспедецевом типе дубового леса. Экспозиция склона южная, уклон ≈ 15º. Травяной покров образует 22 вида, из них 10 относятся к дубравной свите. Общее проективное покрытие травяного яруса 21,95 %. На пробной площади отмечено 11 видов кустарников с общим проективным покрытием 46,6 %. Древостой составляет Quercus mongolica , с проективным покрытием 80 % от общей площади (табл. 1).
Видовой состав растений на пробной площади
Таблица 1
|
Вид |
Проективное покрытие, % |
Вид |
Проективное покрытие, % |
Общее проективное покрытие, % |
|
Травяной ярус |
||||
|
Artemisia keiskeana |
4 |
Carex reventa |
6 |
21.95 |
|
Atractylodes ovata |
1.5 |
Dictamnus dasicarpus |
0.05 |
|
|
Adenophora pereskiifolia |
0.6 |
Viola orientalis |
0.05 |
|
|
Artemisia stolonifera |
1 |
Potentylla fragarioides |
0.1 |
|
|
Convallaria keiskei |
0.5 |
Carex siderosticta |
0.1 |
|
|
Plagiorhegma dubia |
2 |
Moehringia lateriflora |
0.05 |
|
|
Vincetoxicum acuminatum |
0.4 |
Sedum aizoon |
0.05 |
|
|
Vicia unijuga |
4 |
Campanula punctata |
0.6 |
|
|
Geranium maximowiczii |
0.1 |
Pteridium aquilinum |
0.5 |
|
|
Doellingeria scabra |
0.1 |
Lathyrus humilis |
0.1 |
|
|
Melampyrum roseum |
0.1 |
Sanguisorba officinalis |
0.05 |
|
|
К |
старниковый ярус |
|||
|
Quercus mongolica |
22 |
Tilia amurensis |
0.8 |
46.6 |
|
Lespedeza bicolor |
20 |
Pinus koraiensis |
1 |
|
|
Fraxinus rhinhophylla |
0.7 |
Betula davurica |
0.2 |
|
|
Acer mono |
0.7 |
Corylus heterophylla |
0.2 |
|
|
Euonymus pauciflora |
0.5 |
Micromeles alnifolia |
0.2 |
|
|
Maakia amurensis |
0.3 |
|||
|
Древесный ярус |
||||
|
Quercus mongolica \ |
80 1 |
80 |
||
Результаты анализа однородности размещения растений показали, что растения растут, образуя взаимосвязанные группы (куртины или группы куртин) до 6 м в диаметре (табл. 2; рис.). A. keiskeana – это длиннокорневищное растение, преимущественно размножающееся вегетативно, поэтому групповой характер распределения растений свойственен для вида.
Результаты анализов пространственной организации ценопопуляций A. keiskeana
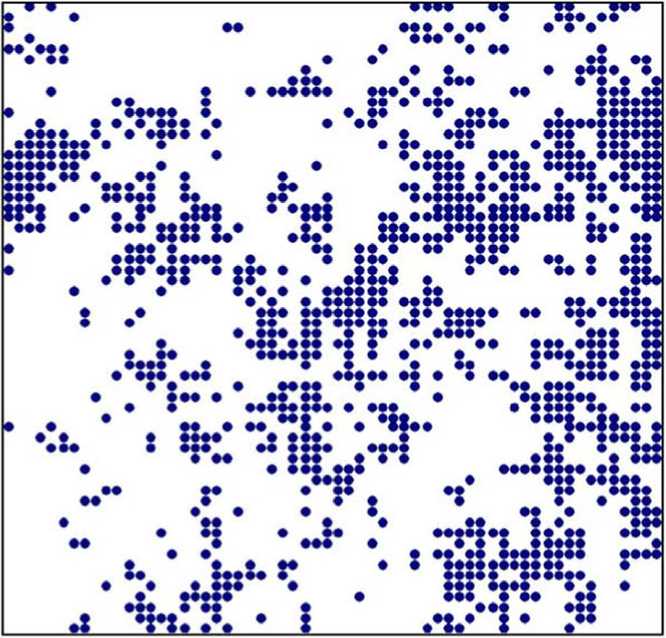
Таблица 2
|
Вид |
Число, шт. |
Расстояние (радиус), м |
||||||||||||||
|
0.5 |
11 |
1.5 1 |
21 |
2.5 1 |
31 |
3.5 1 |
4п |
4.5 1 |
51 |
5.5 1 |
61 |
6.5 1 |
7п |
7.5 |
||
|
Анализ однородности размещения растений |
||||||||||||||||
|
Ak |
4488 |
> |
> |
> |
> |
> |
> |
< |
< |
< |
< |
< |
< |
< |
< |
< |
|
Анализ однородности размещения растений в онтогенетических периодах |
||||||||||||||||
|
Ak (V2) |
3724 |
> |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
|
Ak (G) |
728 |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
~ |
|
Анализ независимости расположения |
растений относительно деревьев |
|||||||||||||||
|
Qm-Ak |
104-4488 |
< |
<п |
< i |
~ п |
<п |
< |
<п |
< п |
<п |
<п |
~ п |
~ п |
~ п |
~ п |
~ |
Примечание: Qm – Quercus mongolica, Ak – Artemisia keiskeana; онтогенетические периоды: V2 – вторая фаза прегенеративного онтогенетического периода; G – генеративный онтогенетический период; ~ – случайное размещение; > – «сближение» анализируемых элементов; < – «отталкивание» анализируемых элементов; цветом обозначена сила притяжения или отталкивания.
Расположение растений A. keiskeana относительно друг друга на пробной площади (точками обозначены побеги растений)
В анализ однородности размещения растений в онтогенетических периодах не были включены растения V1 и S онтогенетических периодов, так как число побегов недостаточно для проведения анализа (табл. 3). Результаты показали, что растения в онтогенетическом периоде G растут внутри куртин независимо друг от друга. Растения в онтогенетическом периоде V2 образуют небольшие слабосвязанные группы до 1 м в диаметре (табл. 2). Вегетативное размножение приводит к большому численному преимуществу молодых побегов онтогенетического периода V2 над другими, поэтому ценопопуляции вида в основном слагают молодые растения (побеги). Взаимосвязь побегов в куртинах до 1 м в диаметре подтверждается биоморфологически-ми особенностями A. keiskeana (длинным корневищем).
Таблица 3 Числовое соотношение побегов A. keiskeana в различных онтогенетических периодах
|
Число побегов, шт. |
|||
|
V1 |
V2 |
G |
S |
|
36 |
3724 |
728 |
– |
Примечание: V1 – первая фаза прегенеративного онтогенетического периода; V2 – вторая фаза прегенера-тивного онтогенетического периода; G – генеративный онтогенетический период; S – сенильный онтогенетический период.
Результаты анализа независимости расположения растений относительно деревьев показали, что A. keiskeana избегает влияния древостоя до 5 м (табл. 2), произрастая в окнах. Это характеризует её как умеренно светолюбивый вид.
В результате проведенных анализов пространственной организации ценопопуляций A. keiskeana выявлен групповой характер распределения растений. Образование взаимосвязанных групп растений или куртин объясняется биологическими особенностями вида. Пазушные почки на корневище развивают ежегодные, моноподиально нарастающие в течение нескольких лет, прямостоячие верхушечные полурозеточные побеги [Бисикалова, 2013], относящиеся к онтогенетическому периоду V2, поэтому ценопопуляции вида в основном слагают молодые побеги.
По жизненной форме и экологическим особенностям [Бисикалова, 2013; Бисикалова, Крестов, 2013] A. keiskeana близка к луговому разнотравью, но на лугах конкуренция со стороны мезофильных луговых видов высокая. В дубовых лесах небольшое затенение кронами ослабляет конкуренцию со стороны потенциальных луговых видов, при этом освещение и уровень влажности удовлетворяют потребностям A. keiskeana . В сильно затенённых кронами деревьев участках A. keiskeana теряет способность нормального развития, поэтому избегает подкроновых пространств, произрастая в окнах древостоев.
