Пространственно-временная динамика видов в сообществе горно-тундрового пояса Хибинских гор (Мурманская область)
")
Автор: Любезнова Н.В.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 2 т.17, 2023 года.
Бесплатный доступ
Нами исследованы пространственная динамика видов сообщества высокогорных тундр Хибинских гор. В сообществе наблюдается жестко структурированная система совместного произрастания видов. Частота встречаемости большинства из них связана положительными или отрицательными корреляциями. В соответствии с карусельной моделью и военной стратегией передвижения видов было выявлено, что виды имеют низкую или среднюю скорость мелкомасштабного круговорота. Видов с высокой скоростью мелкомасштабного круговорота не выявлено. Для большинства видов характерна стратегия «фаланги », причем массовое освоение новых микросайтов происходит раз в несколько лет и для разных видов эти годы не совпадают. Многие виды ограничены в своем передвижении и не могут занимать любые микросайты сообщества.
Мелкомасштабный круговорот, кумулятивная встречаемость, текущая встречаемость, сопряженная встречаемость, высокогорные тундры
Короткий адрес: https://sciup.org/148326704
IDR: 148326704 | УДК: 581.526.2 | DOI: 10.24412/2072-8816-2023-17-2-76-87
Spatio-temporal dynamics of species in the community of the mountain-tundra belt of the Khibiny mountains (Murmansk region)
The spatial dynamics of the species have been studied of the alpine tundra community of the Khibiny Mountains. In the community is observed a rigidly structured system of joint growth of species. The frequency of occurrence of most of them was associated positive or negative correlations. In accordance with the carousel model and the military strategy for the movement of species, it was found that the species have a low or medium speed of small-scale turnover. It have not been identified the species with a high rate of small-scale turnover. For most species, the « phalanx» strategy is typical. The mass development of new microsites occurs every few years and these years do not coincide for different species. Many species are restricted in their movement and cannot occupy each microsites of the community
Текст научной статьи Пространственно-временная динамика видов в сообществе горно-тундрового пояса Хибинских гор (Мурманская область)
Большинству фитоценозов свойственна неоднородность горизонтального сложения (мозаичность): часть видов конкурирует и вытесняет друг друга, другие вполне успешно сосуществуют вместе. Визуально в составе многих фитоценозов выделяются микрогруппировки (Yaroshenko, 1958; Rabotnov, 1983), но количественные подходы к изучению комплексной пространственно-временной динамики растительного покрова фитоценозов начали развиваться только с начала 90-х годов прошлого века.
Работы по исследованию постоянства и скорости передвижения видов на площадках, сравнимых с размером особи были одновременно начаты в Чехии и Швеции (Herben et al, 1993, 1994; van der Maarel, Sykes, 1993). Карусельная модель (van der Maarel, Sykes, 1993) предполагает, что виды, передвигаясь по сообществу, занимают и освобождают микросайты и со временем отмечаются во всех местах – проходят полный круг по сообществу. Эта модель хорошо работает в бедных сообществах, где преобладают однолетники, и наблюдается частая гибель особей. Однако авторы учитывали жизненную форму растений только на уровне однолетник-многолетник, игнорируя морфологическое строение. У них например не получалось адекватно объяснить закономерности передвижения длиннокорневищного Agrostis vinealis Schreb. (van der Maarel, Sykes, 1997). Для некоторых многолетних видов прерий Оклахомы создали модель M.W. Palmer, G.M. Rusch (2001). Модель, основанная на случайной колонизации видами микросайтов, показала, что колонизация немного отклонялось от случайной вероятности: виды имели тенденцию вновь занимать ранее освобожденное пространство чаще, чем ожидалось (Palmer, Rusch, 2001). Статистика мобильности хорошо согласовалась между площадками и годами, хотя время пребывания видов демонстрирует некоторые тонкие зависимости от масштаба площадок. В модели авторы просчитали время полного круговорота для выбранных многолетних видов, и это время составило сотни и даже тысячи лет (Palmer, Rusch, 2001). После этой работы интерес к карусельной модели резко упал. Однако, если не концентрироваться только на конечной цели определения времени полного круговорота видов и учитывать морфологическое строение видов растений, можно выявить интересные стратегии роста популяций.
В Чехии для богатых видами высокогорных лугов Карпат была выявлена взаимосвязь морфологического строения особей и стратегии передвижения видов вкупе с их способностью к удержанию занятых площадей (Herben et al, 1993, 1994). Сопоставление скорости передвижения с «военными» стратегиями роста растений, такими как «партизанская» – растения, способные передвигаться более чем на 3 см в год, и «фаланга» – растения, передвигающиеся медленно, но плотным «строем» (Lovett Doust L., Lovett Doust J., 1982), понимаемых в широком смысле, позволило конкретизировать особенности роста популяций отдельных видов (Herben et al, 1993, 1994).
Вопрос о возможной применимости карусельной модели с дополнением к ней результатов исследований постоянства видов горных лугов (Herben et al, 1993, 1994) и «военных» стратегий роста (Lovett Doust L., Lovett Doust J., 1982) к другим сообществам обсуждался в работе А.А. Маслова (2001). В сложно устроенном сообществе сосняка-черничника большинство видов были «фалангистами» и увеличивали свою кумулятивную встречаемость за счет медленного разрастания (Maslov, 2001). Кроме того, часть видов с высокой мобильностью была ограничена в своем движении и проходила круг только по части доступных микросайтов и среди них – группа видов со строго локальным размещением в специфичных микросайтах. Выделялась особая стратегия роста с высоким удержанием территории и очень низкой мобильностью, названная автором «сиднями» (Maslov, 2001).
Под мобильностью в данном случае понимается способность популяции растений перемещаться по площади сообщества как вегетативно, так с помощью семенного размножения (Maslov, 2001). Кумулятивная встречаемость определяется как общее число всех площадок, которые были заняты видом хотя бы в один год учета за весь период наблюдения. Мелкомасштабный круговорот – свойство популяции постоянно изменять положение особей в пространстве путем колонизации новых микросайтов, освобождения их и вновь колонизации через какое-то время (Maslov, 2001). Колонизация может проходить по «новым» квадратам, которые ни разу не были заняты за период наблюдений, а может и по «старым», которые были заняты ранее, а затем освобождены.
Своеобразные климатические условия высокогорных тундр: небольшой вегетационный сезон, пониженная температура и сильные ветра – приводят к появлению у растений ряда специфических признаков, помогающих переносить неблагоприятные условия (Shilova, 1988). В связи с этим всестороннее изучение эколого-биологических свойств популяций альпийских видов растений становится еще более актуальным. В суровых условиях высокогорий у растений уменьшаются размеры, и увеличивается срок жизни (Pavlov, 1987). Некоторому уменьшению должны подвергнуться и критерии, например, «партизанской» стратегии. В связи с небольшими размерами квадратики 5×5 см сравнимы с размерами многих растений.
М атериалы и методы
Для исследования было выбрано сообщество, расположенное близ вершины южного отрога горы Кукисвумчорр (N67°40'13,33" E33°41'23,51") на склоне юго-западной экспозиции, высота над уровнем моря составляла 850–855 м. Сообщество располагалось на выположенной части склона крутизной 10–15 градусов, прикрытой с севера небольшим гребнем; его площадь составляла примерно 50 на 20 м. В Хибинских горах субнивальный пояс начинается с высоты 1000 м над уровнем моря, поэтому выбранное сообщество можно отнести к верхней части субальпийского пояса. Сообщество представляет собой комплекс фрагментов ассоциаций Cassiopo-Salicetum herbaceae Nordh. 1936 с доминированием мхов в основном Kiaeria starkei (F. Weber & D. Mohr) I. Hagen и K. blyttii (Bruch & Schimp.) Broth. и Loiseleurio-Diapensietum Nordh. 1943 (Korolova, 2001) в которой большую роль играют кустовые лишайники Cetraria islandica (L.) Alh., Flavocetraria nivalis (L.) Kärnefelt et A. Thell., Stereocaulon Hoffm., Bryocaulon divergens (Ach.) Karnefelt, Cladonia uncialis (L.) Weber ex F. H. Wigg., Cl. graciles (L.) Willd., Cl. amaurocraea (Flörke) Schaer., а также имеет и оригинальные черты. В составе фитоценоза есть выходы скал и крупнокаменистые осыпи. Почвы горно-тундровые, сильнокаменистые, сообщество освобождается от снега в середине июня – начале июля в зависимости от толщины снежного покрова и температуры воздуха. Сообщество сильно мозаичное, относительно богатое видами: зафиксировано 38 видов сосудистых растений. Учетные площадки не закладывали на выходах скальных пород, на которых отмечены виды: Avenella flexuosa (L.) Drej., Oxyria digyna (L.) Hill., Cassiope tetragona (L.) D. Don, Cardamine bellidifolia L., Saxifraga oppositifolia L., Beckwithia glacialis (L.) A. & D. Löve, Salix phylicifolia L., Vaccinium myrtillus L. Было заложено 20 площадок 50×50 см, поделенные на квадраты 25×25 см, составленные из квадратиков 5×5 см, где расположение видов картографировалось в 2011–2019 годах. Учетной единицей являлся наземный побег. Для изучения подвижности популяций изучаемых видов и скорости их пространственного мелкомасштабного круговорота между моментами t 1 и t 2 для всех площадок за все годы наблюдений было посчитано: a – совокупность квадратов, занимаемых побегами данного вида на площадке в момент первого и второго учета, b – колонизация (совокупность новых занятых квадратов), c – освобожденные видом квадраты и d – совокупность квадратов, на которых вид отсутствовал (по методике описанной в статье А. А. Маслова (2001)). Также в колонизации отмечалась доля «старых» квадратов, которые были освобождены ранее за время наблюдений. Скорость мелкомасштабного круговорота особей отдельных видов (TR) оценивали по формуле: TR=1-V, где:
ad — be
-
V= , =
-
V (a + b)(c + d)(a + c)(b + d)
Скорость мелкомасштабного круговорота представляет величину от 0 до 1. Величина средней скорости мелкомасштабного круговорота вычислялась из 8 оценок. В анализ не включались единично распространенные на площадках виды Achillea milifolium L. (после двух лет исчезла с площадки), Betula pubescens Ehrh., Salix glauca L., C hamaenerion angustifolium (L.) Holub, который был занесен семенами, но после суровой зимы также исчез. Виды с низкой численностью: Hieracium alpinum L., Huperzia selago (L.) Schrank et Mart и Luzula spicata (L.) DC. также были удалены из анализа. Номенклатура сосудистых растений дана по Н. А. Секретаревой (2004).
Р езультаты
Сопряженная встречаемость видов
Более подробно результаты изложены в нашей статье (Lyubeznova, 2021), но поскольку анализ будет проведен по группам видов, выделенных ранее, мы кратко приведем получившиеся в работе результаты.
Для исследованного сообщества характерна жестко структурированная система совместного произрастания видов, а также полное отсутствие однолетников (Lyubeznova, 2021). Для выяснения этого были посчитаны корреляции Спирмена частоты встречаемости видов между собой.
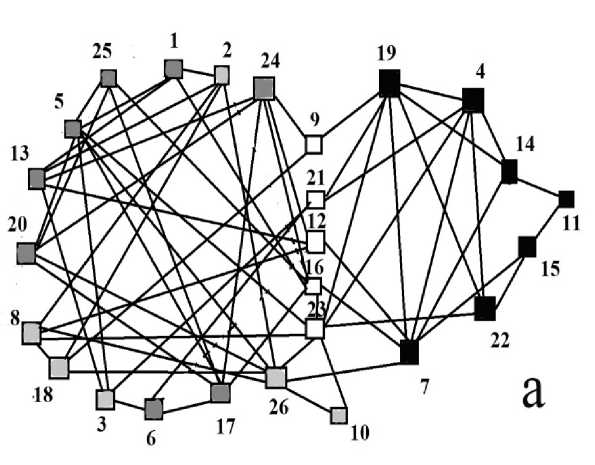
Рис. 1. a. Сопряженная встречаемость видов, положительные корреляции (Lyubeznova, 2021). b Отрицательные корреляции между видами 2 группы.
□ 1 группа, 1 подгруппа, п 2 подгруппа, ■ 2 группа, □ общие виды
Fig. 1. a. Conjugate frequency of species, positive correlations (Lyubeznova, 2021). b Negative correlations between Group 2 species.
□ Group 1, subgroup1, □ subgroup 2, ■ Group 2, □ common species.
1– Antennaria dioica (L.) Gaertn., 2 – Bartsia alpina L., 3 – Campanula rotundifolia L., 4 – Carex bigelovii Torr. ex Schwein., 5 – Dryas octopetala L., 6 – Empetrum hermaphroditum Hagerup, 7 – Festuca ovina L., 8 – Omalotheca supina (L.) DC., 9 – Harrimanella hyphoides (L.) Cov., 10 – Hieracium alpinum L., 11 – Huperzia selago (L.) Schrank et Mart, 12 – Jucus trifidus L., 13 – Loiseleuria procumbens L., 14 – Luzula arcuata (Wahlenb.) Sw., 15 – Luzula spicata (L.) DC, 16 – Lycopodium alpinum L., 17 – Phyllodoce coerulea (L.) Beb., 18 – Polygonum viviparum L., 19 – Salix polaris Wahl., 20 – Salix reticulata L., 21 – Sibbaldia procumbens L., 22 – Silene acaulis (L.) Jacq., 23 – Solidago lapponica Witker., 24 – Vaccinium uliginosum L., 25 – Vaccinium vitis-idaea L., 26 – Veronica alpina L.
Виды сообщества образуют две связанных между собой положительными корреляциями группы (рис. 1a). В первую, более крупную, входят 19 видов, и она частично соответствует составу ассоциации Loiseleurio-Diapensietum Nordh. 1943. Вторая, меньшая группа, включает 13 видов, и состав соответствует ассоциации Cassiopo - Salicetum herbaceae Nordh. 1936. Кроме того, существует 6 видов связанных корреляциями с обеими группами (рис. 1a).
В первой группе можно условно выделить две подгруппы – одна состоит из видов относительно нетронутых местообитаний (рис. 1a), во вторую подгруппу входят виды тяготеющих к немного нарушенным местообитаниям (рис. 1a) а также общие для двух групп виды. Между видами этих подгрупп наблюдается много отрицательных корреляций (табл. 1). Виды первой подгруппы принадлежат к жизненной форме кустарничков (кроме Antennaria dioica (L.) Gaertn.) и наибольшая конкуренция между ними происходит в наземной сфере. Между собой у них также наблюдаются отрицательные корреляции (табл. 1), особенно много у Vaccinium uliginosum L. и Loiseleuria procumbens L. так как имеют корневища и разветвленные надземные многолетние побеги. Phyllodoce coerulea (L.) Beb. однако хорошо сосуществует с Empetrum hermaphroditum Hagerup , а Salix reticulata L. – с видами Vaccinium, Phyllodoce coerulea и Dryas octopetala L.
Таблица 1. Распределение отрицательных корреляций в первой группе (Lyubeznova, 2021). Жирным шрифтом выделены виды первой подгруппы, курсивом – второй, отрицательные корреляции между видами в своей подгруппе выделены жирным шрифтом.
Table 1. Distribution of negative correlations in the first group (Lyubeznova, 2021). Species of the first subgroup are shown in bold and species of the second subgroup are shown in italics; negative correlations between species in their subgroup are shown in bold.
|
a .а а *а .а а а а Й а |
5 t а а 60 а ^ А |
S а «а ^ Cl В а •а |
•2 § ь. а а -а a s •2 8 |
а "а |
а |
а 60 а а а .а ‘а а |
1 *5 .5^ а а |
^2 to S5 ^ 1 '5 |
Со |
|
|
Phyllodoce coerulea (L.) Beb. |
-0,05 |
|||||||||
|
Salix reticulata L. |
-0,08 |
0,14 |
||||||||
|
Vaccinium uliginosum L. |
-0,09 |
-0,09 |
-0,10 |
|||||||
|
Vaccinium vitis-idaea L. |
||||||||||
|
Campanula rotundifolia L. |
-0,07 |
-0,05 |
-0,07 |
-0,06 |
||||||
|
Omalotheca supina (L.) DC. |
-0,05 |
-0,12 |
-0,10 |
-0,10 |
-0,11 |
|||||
|
Harrimanella hyphoides (L.) Cov. |
-0,10 |
-0,05 |
||||||||
|
Jucus trifidus L. |
-0,09 |
-0,08 |
-0,11 |
|||||||
|
Polygonum viviparum L. |
-0,05 |
-0,05 |
0,04 |
-0,06 |
||||||
|
Sibbaldia procumbens L. |
0,05 |
-0,10 |
0,09 |
-0,05 |
-0,09 |
-0,14 |
-0,07 |
|||
|
Solidago lapponica Witker. |
-0,05 |
-0,10 |
0,11 |
-0,05 |
-0,06 |
-0,05 |
||||
|
Veronica alpina L. |
-0,10 |
-0,08 |
-0,07 |
-0,06 |
Внутри второй группы отрицательные корреляции имеются только с общими с первой группой видами за исключением пары Festuca ovina L. и Salix polaris Wahl. (рис. 1b) .
Пространственно-временная динамика
Скорость мелкомасштабного круговорота, с которой вид передвигается по сообществу важная фитоценологическая характеристика. Она зависит от жизненной формы растения, конкурентной стратегии вида, эффективности семенного возобновления и может отличаться у одного вида в сообществах с разной напряженностью конкуренции (Lyubeznova, 2004). Мы оцениваем скорость больше 0.4 как среднюю. 0.3–0.4 как низкую, меньше 0.3 – очень низкую.
Очень низкая скорость мелкомасштабного круговорота характерна для всех видов 1 подгруппы 1 группы, а также для основных видов 2 группы, кроме злаков (рис. 2). Интересно, что все виды с очень низкой скоростью передвижения обладают либо горизонтально лежащими корневищами, либо являются полукустарничками со стелющимися или приподнимающимися наземными побегами. Обладая иногда значительным до 0.5 см годовым приростом, эти виды очень мало осваивают новые квадратики. За годы наблюдений большая часть новых занятых квадратиков была недавно освобождена (табл. 2). Особенно это характерно для Salix polaris. Эти виды обитают в ненарушенных участках сообществ с большими куртинами лишайников или сплошной подушкой мхов для второй группы.
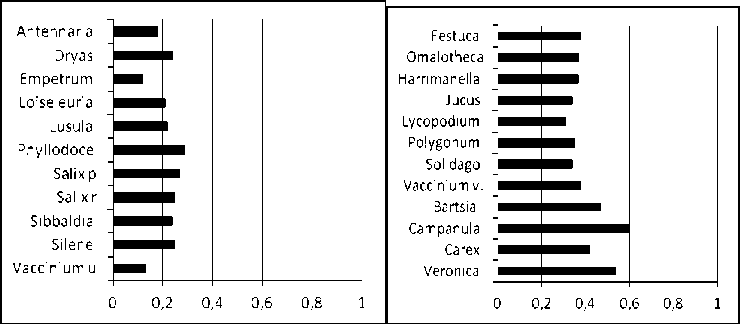
Рис. 2. Средняя скорость мелкомасштабного круговорота в сообществе горнотундрового пояса Хибин.
Fig. 2. Average rate of small-scale turnover in the community of the Khibin’s mountain tundra belt.
Виды с очень низкой скоростью мелкомасштабного круговорота / Species with a very low rate of small-scale turnover: Antennaria – Antennaria dioica (L.) Gaertn., Dryas – Dryas octopetala L., Empetrum – Empetrum hermaphroditum Hagerup, Loiseleuria – Loiseleuria procumbens L., Luzula – Luzula arcuata (Wahlenb.) Sw., Phyllodoce – Phyllodoce coerulea (L.) Beb., Salix p – Salix polaris Wahl., Salix r – Salix reticulata L., Sibbaldia – Sibbaldia procumbens L., Silene – Silene acaulis (L.) Jacq., Vaccinium u – Vaccinium uliginosum L.
Виды с низкой скоростью мелкомасштабного круговорота / Species with a low rate of small-scale turnover: Festuca – Festuca ovina L., Omalotheca – Omalotheca supina (L.) DC., Harrimanella – Harrimanella hyphoides (L.) Cov., Jucus – Jucus trifidus L., Lycopodium – Lycopodium alpinum L., Polygonum – Polygonum viviparum L., Solidago – Solidago lapponica Witker., Vaccinium v – Vaccinium vitis-idaea L.
Виды со средней скоростью мелкомасштабного круговорота / Species with an average rate of small-scale turnover: Bartsia – Bartsia alpina L., Campanula – Campanula rotundifolia L., Carex – Carex bigelovii Torr. ex Schwein., Veronica – Veronica alpina L.
Из видов 2 подгруппы 1 группы только Sibbaldia procumbens L. обладает особо низкой скоростью, остальные имеют низкую скорость мелкомасштабного круговорота, а Campanula rotundifolia L. и Veronica alpina L. среднюю (рис. 2). Большинство из них виды с регулярным семенным возобновлением, такие как Campanula rotundifolia, Festuca ovina, Omalotheca supina (L.) DC. , Jucus trifidus L. , Polygonum viviparum L. , Solidago lapponica Witker. и Veronica alpina , а Lycopodium alpinum L. и Vaccinium vitis-idaea L. длиннокорневищные виды.
Во второй группе Festuca ovina обладает низкой скоростью мелкомасштабного круговорота, за счет семенного возобновления, а длиннокорневищный Carex bigelovii Torr. ex Schwein. – средней скоростью мелкомасштабного круговорота.
Для большинства видов были характерны значительные колебания числа побегов по годам. Такие скачки отражались на текущей встречаемости – числе занятых видом квадратиков. Кумулятивная встречаемость зависит еще и от скорости мелкомасштабного круговорота. Так у видов с очень низкой скоростью мелкомасштабного круговорота на девятый год наблюдений прирост кумулятивной встречаемости составлял менее половины от текущей на тот момент (рис. 3), как у Salix polaris, менее трети у Loiseleuria procumbens или совсем ничтожный как у Empetrum hermaphroditum. У видов с низкой скоростью мелкомасштабного круговорота прирост кумулятивной встречаемости составлял примерно половину от текущей как у Festuca ovina и Vaccinium vitis-idaea (рис. 3), у видов со средней скоростью – превышал текущую встречаемость как у Carex bigelovii (рис. 3).
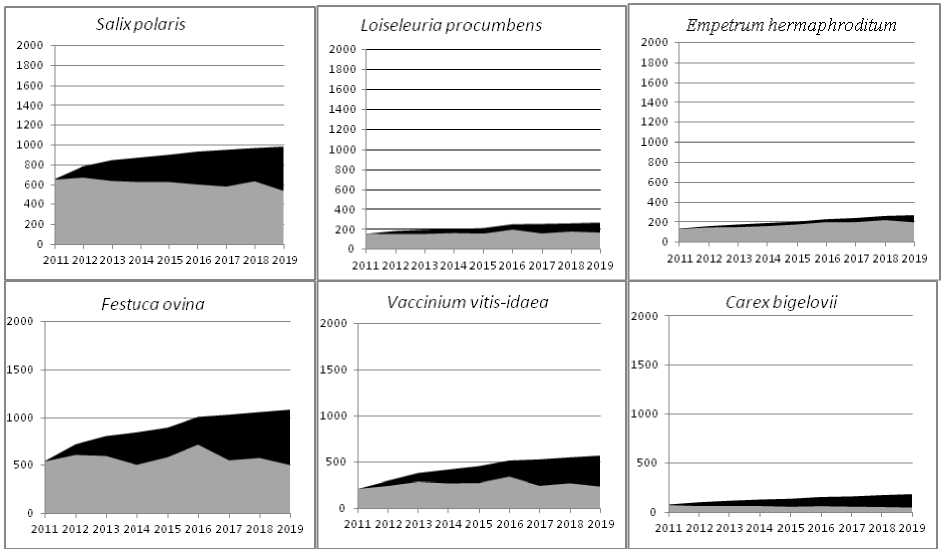
Рис. 3. Текущая и кумулятивная встречаемости для некоторых видов. Ось абсцисс – годы, ось ординат – число квадратиков. □ Текущая встречаемость, ■ кумулятивная встречаемость.
Fig. 3. Current and cumulative frequencies for several species. The abscissa axis is years, the ordinate axis is the number of squares.
□ Current frequency, ■ Cumulative frequency.
Колонизация растением может идти по новым квадратикам, а может по старым, которые были освобождены ранее год назад или более. Способность видов занимать освободившиеся микросайты чаще, чем случайно была отмечена и ранее (Palmer. Rusch. 2001). Для большинства видов высокогорных тундр доля колонизированных старых квадратов сильно выше случайной величины. Для корневищных видов это может быть распускание спящих почек на старых участках корневища, для других – разбрасывание семян недалеко от материнского растение и прорастание банка семян в течение долгого времени. По этому показателю у нас выделилось несколько категорий видов с разными стратегиями.
Первая, в которую входят виды 1–5 (табл. 2), у которых число старых квадратов за все время наблюдений меньше 60 %. то есть виды по большей части передвигаются по площади сообщества. Среди них длиннокорневищные Lycopodium alpinum и Carex bigelovii, хорошо размножающиеся семенами Omalotheca supina и Solidago lapponica. Empetrum hermaphroditum – вид с высоким постоянством и медленным приростом кумулятивной встречаемости (рис. 3).
У другой, в которую входят Jucus trifidus, Polygonum viviparum и Veronica alpina (табл. 2) – виды с хорошим семенным размножением и невысокой численностью в сообществе, доля колонизации старых квадратов более 70% единична на 4–5 год наблюдений. Такое уменьшение колонизации новых квадратов происходит у кого-то на спаде численности, у кого-то на подъеме.
Таблица 2. Доля старых квадратиков в колонизации
Table 2. Proportion of old squares in colonization
|
№ |
Species\ year |
2013 |
2014 |
2015 |
2016 |
2017 |
2018 |
2019 |
|
1 |
Empetrum hermaphroditum |
20,0 |
16,7 |
53,3 |
24,2 |
42,9 |
47,4 |
46,7 |
|
2 |
Omalotheca supina |
55,6 |
53,8 |
60,0 |
47,1 |
58,8 |
64,7 |
50,0 |
|
3 |
Lycopodium alpinum |
20,0 |
33,3 |
60,0 |
20,0 |
36,4 |
26,3 |
50,0 |
|
4 |
Solidago lapponica |
20,0 |
44,0 |
50,0 |
42,4 |
55,6 |
42,6 |
36,4 |
|
5 |
Carex bigelovii |
37,5 |
43,5 |
61,9 |
38,2 |
66,7 |
36,4 |
57,7 |
|
6 |
Jucus trifidus |
30,8 |
50,0 |
75,9 |
41,2 |
58,6 |
59,4 |
50,0 |
|
7 |
Polygonum viviparum |
33,3 |
83,3 |
40,0 |
40,0 |
66,7 |
50,0 |
50,0 |
|
8 |
Veronica alpina |
35,0 |
30,0 |
70,4 |
52,0 |
30,0 |
57,1 |
16,7 |
|
9 |
Dryas octopetala |
28,6 |
50,0 |
65,8 |
60,7 |
56,9 |
71,6 |
72,7 |
|
10 |
Vaccinium uliginosum |
25,0 |
55,6 |
50,0 |
63,6 |
50,0 |
73,7 |
85,7 |
|
11 |
Vaccinium vitis-idaea |
22,0 |
40,9 |
55,7 |
54,7 |
75,0 |
78,5 |
72,2 |
|
12 |
Bartsia alpina |
45,8 |
36,4 |
75,0 |
64,5 |
52,2 |
80,0 |
77,8 |
|
13 |
Antennaria dioica |
20,0 |
50,0 |
30,0 |
40,0 |
83,3 |
81,8 |
40,0 |
|
14 |
Loiseleuria procumbens |
62,5 |
76,7 |
65,5 |
38,1 |
81,8 |
82,4 |
76,7 |
|
15 |
Phyllodoce coerulea |
34,5 |
25,7 |
73,9 |
64,3 |
52,2 |
75,9 |
80,8 |
|
16 |
Salix reticulata |
38,9 |
46,9 |
72,4 |
69,8 |
39,2 |
80,0 |
72,4 |
|
17 |
Silene acaulis |
35,3 |
78,6 |
76,5 |
50,0 |
66,7 |
81,3 |
40,0 |
|
18 |
Campanula rotundifolia |
28,6 |
71,4 |
50,0 |
60,0 |
62,5 |
71,4 |
100,0 |
|
19 |
Festuca ovina |
32,6 |
59,3 |
72,2 |
54,4 |
77,6 |
83,1 |
76,4 |
|
20 |
Harrimanella hyphoides |
39,4 |
33,3 |
48,8 |
70,6 |
91,2 |
63,9 |
68,8 |
|
21 |
Sibbaldia procumbens |
72,2 |
44,4 |
90,0 |
77,8 |
42,9 |
68,8 |
69,2 |
|
22 |
Salix polaris |
37,8 |
72,6 |
74,4 |
73,0 |
83,8 |
85,6 |
76,1 |
В третьей, в которую входят Dryas octopetala, Vaccinium uliginosum, Vaccinium vitis-idaea и Bartsia alpina L. , доля занятых квадратов плавно повышается к 9 году наблюдений (табл. 2). По-видимому, эти виды уже освоили все возможные микросайты и передвигаются по ним.
Еще довольно многочисленная категория: виды № 13–21 (табл. 2), когда при высокой доле колонизации старых квадратиков почти во все годы за время наблюдений, вдруг на 5–7 год доля старых квадратиков падает, а потом опять возрастает. Таким образом, виды колонизируют новые площади раз в несколько лет и такая стратегия характерна для многих видов 1 подгруппы 1 группы.
И Salix polaris (табл. 2), у которой всегда высокая доля колонизации недавно освобожденных квадратиков (табл. 2). В связи с высокой численностью этого вида можно сказать, что он освоил все возможные для него участки.
О бсуждение
В результате наших исследований получилось, что виды высокогорных тундр обладают очень низкой мобильностью, нарушенные участки зарастают десятилетиями. В нарушенных пятнах селятся Omalotheca supina, Festuca ovina, Sibbaldia procumbens и Silene acaulis, но если нарушенный участок более 3 см диаметром то проростки погибают практически полностью, а кусочки лишайников сдуваются и смываются.
Многие вегетативно подвижные виды обладают стратегией «фаланги» дополненной регулярным распусканием спящих почек на старых участках корневищ, где вида уже нет год или два. Многие виды, особенно относящиеся к 1 подгруппе большей группы, предпочитают сохранять уже занятые участки не допуская туда виды со схожей жизненной формой, а Loiseleuria procumbens и Vaccinium uliginosum вытесняют постепенно все другие виды (Lyubeznova, 2021). Кроме того некоторые «фалангисты» активно колонизируют новые квадраты лишь раз в несколько лет (табл. 2).
К «сидням» в понимании А.А. Маслова (2001) можно отнести Empetrum hermaphroditum (рис. 3) и Vaccinium uliginosum – для них характерны высокое удержание территории и очень низкая мобильность. Оба вида также принадлежат к 1 подгруппе 1 группы (рис. 1a).
Среди видов сообщества есть три длиннокорневищных вида: Lycopodium alpinum , Vaccinium vitis-idaea и Carex bigelovii, относящихся к разным подгруппам . Первый, по-видимому, занесен недавно и активно осваивается на небольшом участке вегетативным разрастанием. Carex bigelovii в основном ограничена ассоциацией Cassiopo - Salicetum herbaceae Nordh. 1936, но была встречена и на относительно нарушенных участках. Vaccinium vitis-idaea за время наблюдений постепенно увеличивает число старых квадратов в колонизации, что говорит об ограничении в прохождении сайтов. Например, невозможность для вида с гипогеогенным корневищем и очень редким плодоношением преодолевать участки с высокой каменистостью. К этим видам условно можно применить «партизанскую» стратегию по меркам высокогорных тундр, где корневищ более 3 см в год никто не формирует.
Виды, размножающиеся в основном семенами: Omalotheca supina , Jucus trifidus , Polygonum viviparum , Solidago lapponica , Campanula rotundifolia , Veronica alpina принадлежат ко второй подгруппе 1 группы (рис. 1a) . Это виды с низкой и средней скоростью мелкомасштабного круговорота. Проростки у них возникают ежегодно или довольно часто, но их приживаемость низкая. Festuca ovina имеет низкую скорость мелкомасштабного круговорота тоже только благодаря проросткам с высокой смертностью. Вид бывает плотнокустовым на открытых участках и рыхлокустовым в зарослях лишайников. В некоторые зимы происходит отмирание всех побегов, но в последующие годы распускаются спящие почки и особь восстанавливается.
Bartsia alpina в своем передвижении зависит от распространения видов-хозяев, к которым относятся Antennaria dioica , Omalotheca supina , Loiseleuria procumbens , Polygonum viviparum , Veronica alpina (Lyubeznova, 2021).
З аключение
Большая часть видов ненарушенных местообитаний имеет очень низкую мобильность и сохраняет уже занятые местообитания. Для этой цели на освобожденных квадратиках остаются подземные корневища со спящими почками. Массовая колонизация новых квадратиков у них происходит раз в несколько лет.
Треть видов продолжает осваивать площадь сообщества. Текущая встречаемость всех видов зависит от погодных условий и сильно колеблется по годам. Кумулятивная встречаемость зависит еще от скорости мелкомасштабного круговорота.
Исследование выполнено в рамках научного проекта государственного задания МГУ «Изучение закономерностей морфогенеза и формирования элементов продуктивности под влиянием факторов внешней среды у высших растений» госбюджет, раздел 0110 (для тем по госзаданию), номер 12-1-21, номер ЦИТИС 121032500082-2.
Список литературы Пространственно-временная динамика видов в сообществе горно-тундрового пояса Хибинских гор (Мурманская область)
- Herben T., Krahulec F., Hadincova V., Kovarova M. 1993. Small-scale spatial dynamics of plant species in a grassland community over six years — Journal of Vegetation Science. 4(2): 171-179.
- Herben T.. Krahulec F., Hadincová V., Pechácková S. 1994. Is a grassland community composed of coexisting species with low and high spatial mobility? — Folia Geobotanica et Phytotaxonomica. 29: 459-4б8.
- [Korolova] Королева H. E. 2001. Синтаксономический обзор горнотундровой растительности Хибин. — Бюл. МОИП. Отд. биол. 10б (4): 50-57.
- Lovett Doust L., Lovett Doust J. 1982. The battle strategies of plants — New Scientist. 95(1313): 81-84.
- [Lyubeznova] Любезнова H3. 2004. Пространственная структура и динамика численности побегов Agrostis vinealis ssp. planifolia (C. Koch.) Tzvel. и Anthoxanthum odoratum L. в высокогорьях северо-западного Кавказа. — В кн.: Комплексные исследования альпийских экосистем Тебердинского заповедника. Труды Тебердинского государственного биосферного заповедника. М. С. 3-45.
- [Lyubeznova]Любезнова H3. 2021 Сопряженная встречаемость видов в сообществе горно-тундрового пояса Хибинских гор (Мурманская область). — Труды Кольского научного центра PAH. 9(12): 45-50.
- [Maslov] Маслов А.А. 2001. Пространственно-временная динамика популяций лесных растений в проверке «карусельной модели» на примере сосняка-черничника — Бюл. МОИП. 106(5): 59-б5.
- [Pavlov] Павлов В.И 1987. Особенности растений в экстремальных условиях высокогорий Средней Азии. — Бюл. МОИП. Отд. биол. 92(5): 89-9б.
- [Rabotnov] Работнов Т.А. 1983. Фитоценология. 292 с.
- [Secretareva] Секретарева H.A. 2004. Сосудистые растения Российской Арктики и сопредельных территорий. 131 с.
- [Shilova] Шилова H^. 1988. Ритмы роста и пути структурной адаптации тундровых растений. 212 с.
- van der Maarel E., Sykes M.T. 1993. Small-scale plant species turnover in limestone grassland: the carousel model and some comments on the niche concept. — Journal of Vegetation Science. 4(2): 179-188.
- van der Maarel E., Sykes M.T. 1997. Rates of small-scale species mobility in alvar limestone grassland. — Journal of Vegetation Science. 8(2): 199-208.
- [Yaroshenko] Ярошенко П.Д. 1958. К изучению горизонтального расчленения растительного покрова. — Ботанический журнал. 43(3): 381-387.
