Пространственно-временная структура гнездовых поселений пеночки-зарнички на западном макросклоне Баргузинского хребта

Автор: Мещерягина Светлана Галимзяновна, Бачурин Геннадий Николаевич, Ананин Александр Афанасьевич
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Зоология
Статья в выпуске: 4, 2015 года.
Бесплатный доступ
Пеночки-зарнички Phylloscopus inornatus в центральной части западного макросклона Баргузинского хребта формируют пространственные группировки гнездовых поселений. На территории Баргузинского государственного природного биосферного заповедника осуществляется долговременный мониторинг населения данного вида пеночек на постоянном ключевом участке. В июне - июле 2012-2013 гг. на указанной территории были исследованы гнездовые поселения пеночек-зарничек в долинах рек Давше и Правый Таркулик. Проводились поиск гнезд, это- логические наблюдения и анализ местообитаний. Обсуждаются новые данные по структуре заселения местообитаний, биологии гнездования и поведению у этого вида.
Гнездовые поселения, многолетняя динамика, пеночка-зарничка phylloscopus inornatus, пространственное распределение
Короткий адрес: https://sciup.org/148183863
IDR: 148183863 | УДК: 598.9(571.54)
Spatio-temporal structure of the breeding settlements of yellow-browed warbler in the western slopes of the Barguzinsky range
Yellow-browed Warbler Phylloscopus inornatus in the central part of the western slopes of the Barguzinsky Range form spatial grouping of breeding populations. In the territory of the Barguzinsky State Nature Biosphere reserve a long-term monitoring of warblers population of this species is carried out at the constant key site. In June-July 2012-2013 in the mentioned above territory the Yellow-browed Warbler breeding settlements were studied in the river valleys of the Davsha and Pravy Tarkulik. The search of nests, ethological observations and analysis of habitat were conducted. The new data on the structure of the settlement habitat, breeding biology and behavior of this species are discussed.
Текст научной статьи Пространственно-временная структура гнездовых поселений пеночки-зарнички на западном макросклоне Баргузинского хребта
Пеночка-зарничка Phylloscopus inornatus (Blyth, 1842) — гнездящийся вид на всей территории Байкальской Сибири. В пределах области встречаются две формы Ph. (inornatus) inornatus (практически вся территория Байкальской Сибири) и Ph. (inornatus) humei (хр. Хамар-Дабан) [7]. Исследуемая территория — западный макросклон Баргузинского хребта — находится вне зоны пространственного контакта форм humei и inornatus [20]. Для определения подвидовой принадлежности птиц, формирующих изучаемые гнездовые поселения, в 2012 г. нами добыты на высоте 1480–1550 м над ур. м. 3 самца ad., которые переданы на хранение в фонды Зоологического музея Московского университета. Данные экземпляры определены Я. А. Редькиным как Ph. (inornatus) inornatus .
Характерной особенностью размножения пеночки-зарнички является склонность к гнездованию изолированными группами. Данная черта гнездового поведения у этого вида пеночки подмечена сравнительно недавно [13, 23] и весьма мало изучена [9, 10, 25, 26].
Пеночки-зарнички Phylloscopus inornatus в центральной части западного макросклона Баргузин-ского хребта формируют пространственные группировки гнездовых поселений. На указанной территории осуществляется долговременный мониторинг населения данного вида пеночек на постоянном ключевом участке (с 1984 г.). На основании полученных результатов выявлены оптимальные и субоптимальные местообитания для пеночки-зарнички, оценена возможность межгодовых перераспределений видового обилия на высотном экологическом профиле и между соседними речными долинами, а также оценена связь долговременных изменений численности с метеорологическими параметрами конкретного года [4].
Механизмы, поддерживающие устойчивую связь вида с определенным ландшафтно-биотопическим окружением и конкретной территорией, рассмотрены К. Е. Михайловым и Н. Н. Балацким [14]. P. H. W. Biedermann [25] рассматривает гипотезу скрытого пения как главное объяснение образования скоплений, показывая, что самцы с территориями в центре территории-конгрегации являются сексуально самыми привлекательными для самок.
Однако эти исследования осуществлялись в поселениях зарничек, характеризующихся авторами как обособленные гнездования за пределами основного ареала. Поэтому в задачу работы входило охарактеризовать пространственно-временную структуру многочисленных группировок гнездования, найденных в основной области распространения вида.
Исследователи структуры группировок пеночек-зарничек, причин и механизмов их возникновения используют для определения такой формы гнездования различную терминологию.
В. К. Рябицев, анализируя территориальные отношения сообщества птиц, называет пеночку-зарничку самым склонным к групповому гнездованию видом среди изученных неколониальных птиц Субарктики [21]. Для характеристики гнездования этого вида применяется понятие « парцелла », введенное Н. П. Наумовым, то есть группа животных («большая семья»), живущих в непосредственной близости друг к другу и потому тесно контактирующих между собой [18].
И. М. Марова, описывая биологию размножения тусклых зарничек в Западном Танну-Ола, придерживается понятия одновидовые поселения , характеризуя их так группы из гнездящихся по соседству птиц, отделенных от других птиц своего вида большими расстояниями [13].
К. Е. Михайлов и Н. Н. Балацкий, выясняя статус пребывания и характер распределения пеночки-зарнички на южной границе ареала в северо-восточном Приморье, применяют термин микропоселение — крошечное изолированное поселение, состоящее из нескольких пар на небольшой территории, вокруг которой в радиусе многих километров вид не обнаружен [14].
P. H. W. Biedermann, исследуя процесс формирования скоплений пеночек-зарничек в северовосточной части Монголии, употребляет понятие гнездовая группировка — гнездящиеся пары распределены по подходящей среде обитания не беспорядочно, а объединены в группы. В отличие от колониального размножения каждая пара еще имеет свою собственную защищаемую территорию [25].
O. V. Bourski & W. Forstmeier, основываясь на многолетние данные по гнездованию пеночек-зарничек в Центральной Сибири, исследовали функции агрегации мест гнездования этого вида и выдвинули гипотезы, объясняющие причины группирования [26]. При этом берется за основу понятие территориальная группировка .
-
А. А. Ананин, выявляя особенности долговременной динамики населения пеночки-зарнички на Западном макросклоне Баргузинского хребта, использует в качестве структурной единицы пространственную группировку [4]. Под этими последними двумя понятиями рассматривалось население птиц определенного местообитания (биотопа).
В данной работе для характеристики социальной организации мест размножения применяется понятие гнездовое поселение . Под этим понимается гнездовая группировка, неоднократно возникающая на определенной территории и образованная одной или несколькими парцеллами. Регулярность возникновения группировки определяет долгосрочность гнездового поселения. Таким образом, ежегодное гнездование характеризует поселение как долгосрочное (регулярное); не ежегодное, но периодичное — как краткосрочное (нерегулярное); однократное гнездование на территории, ранее не используемой данным видом, не рассматривалось как гнездовое поселение.
Материалы и методы
Долговременный мониторинг населения модельного вида на постоянном ключевом участке выполнен в гнездовой период 1984–2007 гг. на постоянных учетных маршрутах в долинах трех рек, охватывающих высотный профиль от побережья оз. Байкал до высокогорий (460–1 700 м над ур. м.). На этих профилях, разбитых на 11 участков, представлена большая часть разнообразия местообитаний нижней и верхней части горнолесного и подгольцово-субальпийского поясов [1, 5]. Общая протяженность летних пеших маршрутных учетов, положенных в основу настоящей работы, — 7 400 км. Обилие птиц рассчитано по методу Ю. С. Равкина [19].
Сведения по гнездовой экологии были собраны в Северо-Восточном Прибайкалье на территории Баргузинского государственного природного биосферного заповедника. Исследования осуществлялись на участках с долговременно существующими регулярными гнездовыми поселениями пеночек-зарничек, охватывающих высотный профиль от побережья оз. Байкал до высокогорий (табл. 1): три — в долине р. Давше и одно — в долине р. Правый Таркулик.
Таблица 1
Размещение мест гнездования пеночек-зарничек в центральной части Баргузинского заповедника
|
Высотный пояс растительности |
Локализация в долине реки |
Статус места гнездования |
№ участка |
Абс. высоты, м над ур. м. |
Координаты размещения |
Площадь, га |
|
Прибрежноравнинный |
Давше, 3–4 км |
многократное нерегулярное |
1 |
480 |
54 º22 'N, 109 º33 'E |
3,0 |
|
Давше, 5 км |
однократное |
- |
500 |
54 º22 'N, 109 º34 'E |
2,0 |
|
|
Предгорный |
Давше, 9–11 км |
многократное нерегулярное |
2 |
500–525 |
54 º23 'N, 109 º37 'E |
32,0 |
|
Давше, 12 км |
однократное |
- |
530 |
54 º24 'N, 109 º39 'E |
2,0 |
|
|
Давше, 12,5–13,5 км |
регулярное |
3 |
510–530 |
54 º23 'N, 109 º42 'E |
33,0 |
|
|
Горнолесной |
Давше, 16,5 км |
однократное |
- |
570 |
54 º21 'N, 109 º47 'E |
1,5 |
|
Давше, 26,5 км |
регулярное |
4 |
1220–1310 |
54 º21 'N, 109 º47 'E |
31,0 |
|
|
Подгольцовосубальпийский |
Правый Таркулик, истоки |
регулярное |
5 |
1300–1420 |
54 º20 'N, 109 º50 'E |
48,0 |
|
Давше, 27,5–28,5 км |
регулярное |
6 |
1380–1590 |
54 º20 'N, 109 º48 'E |
45,0 |
Кроме того, в 2013 г. при увеличении численности вида были исследованы еще два участка с нерегулярно возникающими гнездовыми поселениями в долине р. Давше и зафиксированы три места однократного гнездования 1–2 пар пеночек-зарничек. Обследуемые местообитания классифицированы соответственно выделам высотных поясов растительности, используемых в работах А. А. Ананина [5].
Основные исследования выполнены в гнездовой период 2012–2013 гг., однако в качестве дополнения использовали данные за 2005–2007 гг. (табл. 2).
Таблица 2
Сроки проведения орнитологических исследований на территории Баргузинского заповедника
|
Вид исследования |
Предшествующие |
Основные |
|||
|
Год |
2005 1, 2 |
2006 2 |
2007 2, 3 |
2012 1, 3 |
2013 1, 3 |
|
Первый обход |
5–20.VII |
5–9.VII |
6–10. VII |
30.VI–21.VII |
22.VI–17.VII |
|
Второй обход |
19.VII |
15–16. VII |
|||
|
Всего дней работ |
16 |
6 |
7 |
22 |
26 |
Примечание: 1 — исследования осуществлены Г. Н. Бачуриным, 2 — И. Ф. Вурдовой, 3 — С. Г. Мещерягиной.
В перечисленные периоды исследований наиболее изучено гнездовое поселение на участке 4 в верхней части горнолесного пояса долины р. Давше. В период 2005–2007 гг. поселения на участках 5 и 6 обследовали частично. Поиск гнезд в поселениях на участках 1, 2 и 3 осуществляли лишь в 2013 г. (в предыдущие сезоны только фиксировали поющих самцов).
Ежегодно на каждом обследуемом участке при первичном обходе учитывали число поющих самцов, демонстрирующих беспокойство пар птиц, таким образом, определяя примерное количество гнездовых участков. В дальнейшем при систематических обходах данный показатель корректировали. По итогам пятилетних исследований, учитывая местоположение участков с найденными гнездами и территорий с поющими самцами, выявили для регулярно изучаемых участков (4, 5 и 6) потенциально-возможную площадь заселения в гнездовом поселении.
На каждом отдельном участке выискивали максимально возможное число гнезд, но в 2013 г. при высокой численности вида на отдельных участках осуществляли поиск только периферийных и некоторых центральных гнезд (участки 3, 5 и 6).
Поиск гнезд осуществляли 3 методами: визуальным обнаружением, вспугиванием с гнезда ("кошением") и выслеживанием птиц на гнездовых участках. При обнаружении гнезда определяли его месторасположение с помощью DPRS-навигатора Garmin GPSMAP 62. Определение расстояния между гнездами осуществлялось в программе Google earth.2013 с помощью инструмента «линейка– линия». Расчет площадей участков осуществляли в этой же программе, очерчивая при помощи инструмента «линейка — многоугольник» общий контур вокруг зафиксированных точек (обнаруженных гнезд и учтенных гнездовых участков без находки гнезд) на расстоянии 50–60 м от краевых точек.
В основу построения пространственно-временной последовательности возникновения гнезд на участках использовали дату откладки первого яйца в каждом зафиксированном гнезде. Данный показатель рассчитывали из схемы «12 суток насиживания с четвертого яйца», основанной на собственных наблюдениях и литературных данных по пеночке-зарничке [22]. Насиженность яиц в кладке определяли в момент обнаружения гнезда. При этом просвеченные с помощью овоскопа яйца сравнивались с фотографиями яиц точно известного срока инкубации (собственные данные). В дальнейшем достоверность определения срока насиживания контролировали по дате вылупления птенцов. В случае обнаружения гнезда с птенцами определяли их возраст, пользуясь методикой Мальчевского [12] для мелких воробьиных птиц, гнездящихся открыто, и сравнивая с фотографиями птенцов точно известного возраста (собственные данные).
Статистические расчеты реализованы с применением пакетов программ Statistica 6.0 и Excel.
Результаты
В результате анализа долговременных изменений численности пеночки-зарнички на постоянных участках наблюдений к оптимальным местообитаниям с высоким уровнем среднемноголетней численности и минимальной изменчивостью обилия вида были отнесены предгорный, прибрежноравнинный и подгольцово-субальпийский выделы долины р. Давше, а также горнолесной участок долины р. Езовка и прибрежно-равнинный выдел долины р. Большая. На всех из них зарегистрированы групповые поселения пеночек-зарничек, которые предпочитают березовые и ивовые кустарники по болотам, заболоченные берега рек, увлажненные высокогорные редколесья и кустарниковые заросли [2].
Таблица 3
Локальная плотность гнездящихся пар в поселениях пеночки-зарнички на фоне изменений численности вида в целом по территории Баргузинского заповедника в исследуемые периоды
|
Год |
Количество гнездовых пар (из них найденных гнезд) / площадь, эксплуатируемая в гнездовом поселении в текущем году, га |
Локальная плотность, пар/га |
Численность, особ/км2 |
|||||
|
№ 1 |
№ 2 |
№ 3 |
№ 4 |
№ 5 |
№ 6 |
|||
|
2005 |
- |
- |
+ |
12(7) 9.0 |
3(2) 2.4 |
3(1) 2.3 |
1.3 |
2.59 |
|
2006 |
- |
- |
+ |
13(2) 10.8 |
7(4) 6.7 |
3(1) 2.7 |
1.1 |
1,85 |
|
2007 |
- |
- |
+ |
25(13) 23.8 |
11(8) 10.0 |
4(3) 4.2 |
1.0 |
1,46 |
|
2012 |
- |
- |
2(1) 1.7 |
39(35) 27.4 |
19(18) 17.6 |
28(19) 30.4 |
1.1 |
2,79 |
|
2013 |
3(1) 2.8 |
17(14) 15.0 |
13(10) 11.1 |
30(19) 23.4 |
33(21) 29.1 |
25(15) 30.3 |
1.1 |
11,79 |
|
lim, га/пару |
0.9 |
0.9 |
0.9 |
0.7–1.0 |
0.8–1.0 |
0.8–1.2 |
||
Изменения численности вида в подгольцово-субальпийском поясе р. Давше статистически не связаны с динамикой во всех обследованных прибрежно-равнинных, предгорных и горнолесных местообитаниях, что может подтверждать наличие отличающихся популяционных группировок, населяющих лесную зону и субвысокогорье западного макросклона Баргузинского хребта. Ранее нами было отмечено, что изменение численности вида в высокогорной части Баргузинского хребта (гольцовоальпийский и подгольцово-субальпийский пояса) происходит синхронно [3]. Также было отмечено, что при более раннем прилете пеночки-зарнички регистрируется повышение ее численности в долине р. Давше и для всего ключевого участка в целом [3].
За период исследований (2012–2013 гг.) в целом на всех участках с обследованными гнездовыми поселениями зарничек обнаружено 153 гнезда из 209 зафиксированных гнездящихся пар, а также выявлено в трех местах однократного гнездования 5 гнездящихся пар. В 2005–2007 гг. на трех участках (4, 5 и 6) всего зафиксировано 81 гнездовая пара, из них у 41 выявлены гнезда (табл. 3).
Площадь использования территорий для гнездования пеночками-зарничками в отдельные годы отследили на трех соседствующих участках (рис. 1), где между гнездовым поселением 4 и 6 минимальное расстояние составляет 650 м, а между 6 и 5 — 850 м.
Динамика количественного соотношения птиц в гнездовых поселениях от общего числа зафиксированных в них гнездящихся пар в исследуемом году представлена для тех же участков (рис. 2).
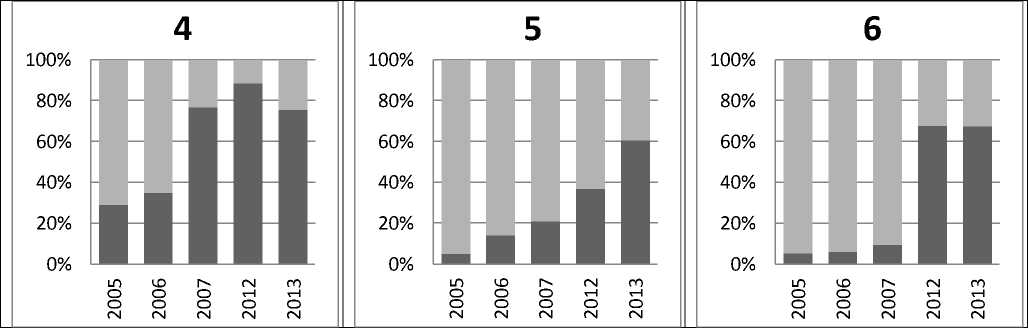
Рис. 1. Доля используемой в различные годы для гнездования площади (темно-серые столбцы) от потенциально-возможной площади заселения в гнездовом поселении (светло-серые столбцы)
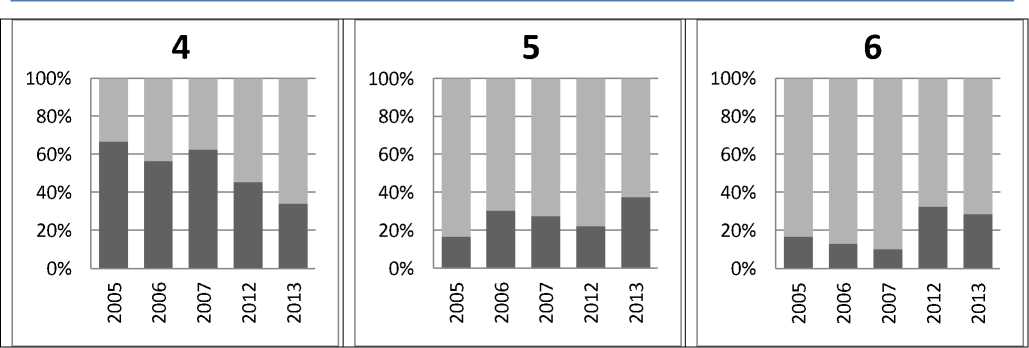
Рис. 2. Доля птиц в гнездовых поселениях (темно-серые столбцы) от общего числа зафиксированных гнездящихся пар в исследуемом году (светло-серые)
Всего удалось отследить сроки откладки яиц в 183 гнездах: в 2013 г. — 80, в 2012 — 68, в 2007 — 21, в 2006 — 6 и в 2005 — 8 гнездах (рис. 3).
|
S |
я |
я |
я |
||||||||||||||||||
|
У |
част |
ки 1 |
, 2 и |
3 |
|||||||||||||||||
|
— |
= |
—■ |
— |
— |
— |
||||||||||||||||
|
Участок 4 |
|||||||||||||||||||||
|
— |
|||||||||||||||||||||
|
Участок 5 |
|||||||||||||||||||||
|
5 |
о |
о |
о |
о |
5 |
5 |
5 |
о |
5 |
5 |
|||||||||||
|
480–530 м над ур. м. |
|||||||||||||||||||||
|
1220–1310 м над ур. м. |
|||||||||||||||||||||
|
— |
|||||||||||||||||||||
|
= |
|||||||||||||||||||||
|
— |
|||||||||||||||||||||
|
— |
|||||||||||||||||||||
|
— |
|||||||||||||||||||||
|
LLL |
|||||||||||||||||||||
|
1300–1420 м над ур. м. |
|||||||||||||||||||||
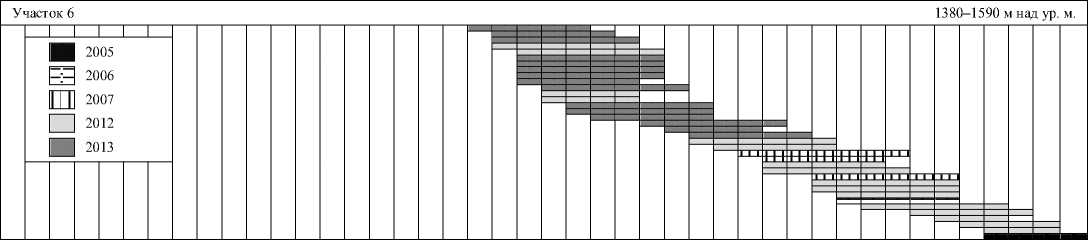
Рис. 3. Обобщенные данные о сроках откладки яиц пеночками-зарничками на отдельных участках
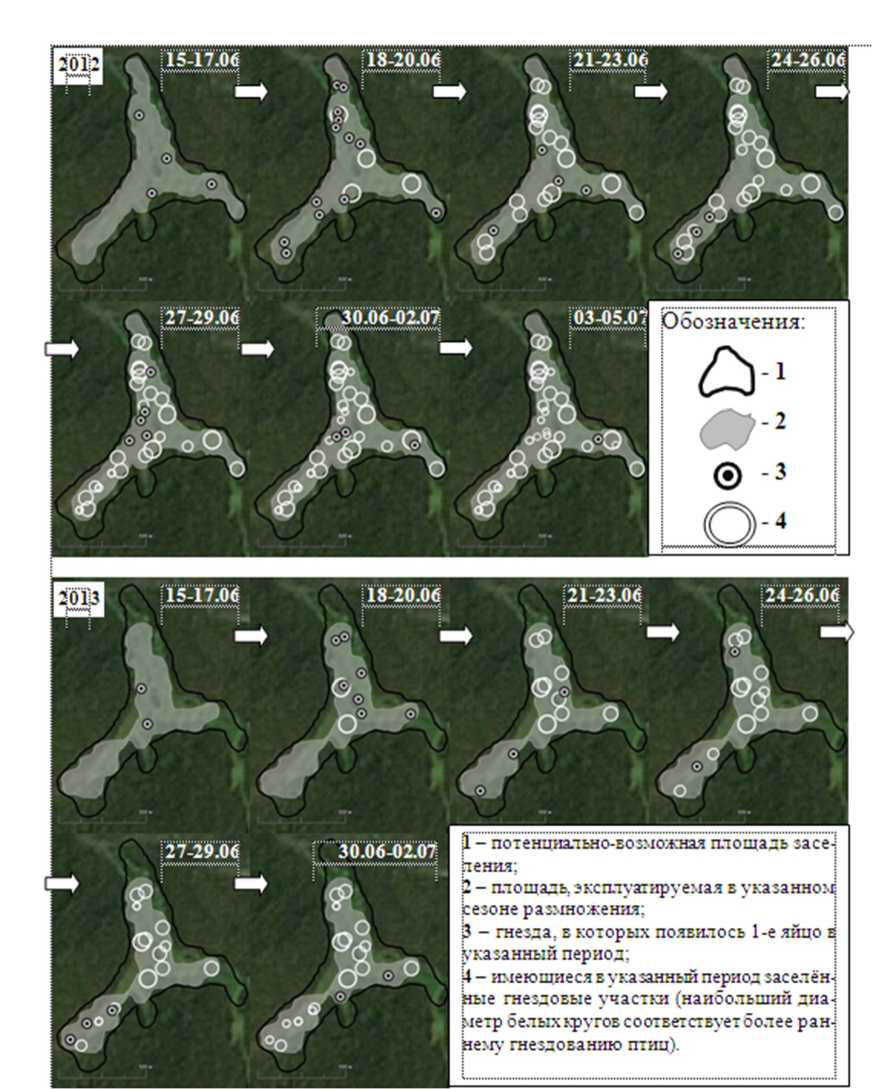
Рис. 4. Пространственно-временная структура формирования гнездового поселения пеночек-зарничек на примере 4 участка в 2012 и 2013 гг.
На основе данных о сроке откладки первого яйца и GPS-координат размещения гнезда реконструировали пространственно-временную структуру формирования поселений в 2012 и 2013 гг. для трех гнездовых поселений пеночек-зарничек на участках 4, 5 и 6 (рис. 4). Для тех же гнездовых поселений на основе данных о сроке откладки первого яйца и расстоянии до уже имеющегося на участке ближайшего гнезда с кладкой выявили пределы линейных параметров уплотнения поселений пеночек-зарничек (табл. 4).
Таблица 4
Минимальные расстояния (R ± SE , в м) между вновь возникающими и имеющимися гнездами пеночек-зарничек в трех гнездовых поселениях (4, 5 и 6)
|
Период гнездования |
Характер размещения вновь возникающих гнезд относительно имеющихся заселенных гнездовых участков |
||
|
«за пределами» |
«между» |
«в пределах» |
|
|
2012 |
266±57 (n=22) |
119±21 (n=21) |
53±8 (n=23) |
|
[125–571] |
[72–238] |
[21–79] |
|
|
2013 |
301±62 (n=17) |
135±22 (n=18) |
55±11 (n=17) |
|
[100–490] |
[86–244] |
[15–86] |
|
|
обобщенный |
281±39 (n=39) |
126±14 (n=39) |
54±6 (n=40) |
|
за 2 года |
[100–571] |
[72–244] |
[15–86] |
Обсуждение
В целом опубликованных данных о существовании долгосрочных гнездовых поселений пеночек-зарничек в конкретных местах весьма ограниченное количество. В. В. Морозов описывает два случая наблюдений долгосрочных гнездовых поселений пеночек-зарничек на западном макросклоне Полярного Урала: в еловом редколесье на г. Южной в период с 1986 по 1993 г. [15], в лиственничной ерни-ковой редине в устье р. Хороташор в 2000 [16] и 2007 гг. [17].
P. H. W. Biedermann [25], описывая два участка скоплений зарничек в северо-восточной части Монголии (научно-исследовательская станция «Khonin Nuga»), указывает, что один из них был заселен как минимум два года (2002–2003 гг.).
На территории Баргузинского заповедника выявлено три варианта социальной организации мест размножения пеночки-зарнички: регулярное (долгосрочное) гнездовое поселение, многократное нерегулярное (краткосрочное) гнездовое поселение и однократное гнездование. Одновременно все указанные варианты зарегистрированы нами в сезон размножения 2013 г., характеризующийся высоким уровнем среднемноголетней численности вида (табл. 3). В основном на исследуемой территории пеночки-зарнички образуют регулярные (долгосрочные) гнездовые поселения. Так, изученное нами гнездовое поселение № 4 на указанном месте (табл. 1) ежегодно регистрировали в течение 35 лет: А. В. Федоров с 1979 по 1987 г. и А. А. Ананин с 1984 по 2013 г. (архивные материалы Баргузинского заповедника).
Площадь, эксплуатируемая пеночками-зарничками в каждом отдельном гнездовом поселении, по годам различается (табл. 3). Однако не зависимо как от варианта социальной организации мест размножения, так и от уровня среднемноголетней численности вида одна пара птиц в среднем использовала 0,7–1,2 га (табл. 3). Для тусклой зарнички приводится более широкий диапазон указанного параметра. Так, в Заилийском Алатау на участке 2,5 га обнаружено 8 жилых гнезд, на участке около 5 га — 20 гнезд [9, 10]; в южных отрогах хр. Западный Танну-Ола на территории около 12 га — 10 гнезд [13].
Таким образом, в годы повышенной численности вида при заселении в каждом отдельном гнездовом поселении потенциально-возможной площади для размножения возможно возникновение мест однократного гнездования. Так, в 2013 г. нами зафиксированы в прибрежно-равнинном, предгорном и горнолесном высотных поясах (табл. 1) по одному случаю однократного гнездования пеночек-зарничек в количестве 1–2 пары. В подгольцово-субальпийском поясе (гнездовое поселение 5 и 6), где доля используемой площади составляла в 2013 г. примерно 60% потенциально-возможной площади заселения в гнездовом поселении (рис. 1), случаев однократного гнездования не зафиксировано.
По мнению В. К. Рябицева [21], свойственные зарничкам поселения из нескольких гнездящихся по соседству пар могут еще в предгнездовое время распадаться и образовываться в другом месте. Так, в Хан-Хэнтэйском заповеднике после регистрации на участке двух первых поющих самцов было отмечено четыре–пять стай (от 8 до 22 самцов), устанавливающих во время миграции краткосрочные территории в течение 1–10 дней на этом участке [25]. Нами такие наблюдения на территории Баргузин-ского заповедника не отмечались, что, возможно, связано с более поздними сроками проведения полевых работ. В целом ежегодно в гнездовых поселениях отмечались самцы, поющие в течение всего гнездового сезона и не проявляющие в поведении практически беспокойства на присутствие наблюдателя в пределах эксплуатируемой самцом территории. По нашему мнению, эти самцы составляют популяционный резерв и в размножении в текущем году не участвуют. В случае же разорения гнезд, исходя из неоднократных наблюдений, самцы перестают беспокоиться, зато начинают активно петь по периметру эксплуатируемого участка, а через 1–2 сутки исчезают с участка.
Численность пеночек-зарничек в гнездовых поселениях ежегодно варьирует (табл. 3). Так, например, В. В. Морозовым [15] была отражена нестабильность численности зарнички на западном макросклоне Полярного Урала, на участке одного гнездового поселения в течение нескольких лет: в июле 1986 г. найдено 2 поющих самца, в 1987 — 6 пар, в июне 1989 — 11 территориальных самцов, а в июне 1993 — 4 пары и 1 территориальный самец. В наших наблюдениях получился подобный результат, когда на фоне изменения общей численности зарничек на территории заповедника (табл. 3) одни поселения укрупнялись (№ 5), другие уменьшались (№ 4), а в некоторых (№ 6) доля от общей численности оставалась без изменений (рис. 2). Аналогичные несинхронные изменения численности вида регистрировались и на отдельных участках постоянных учетных маршрутов на всей ключевой территории в Баргузинском заповеднике в 1984–2014 гг. [4].
На западном макросклоне Баргузинского хребта гнездовые поселения пеночек-зарничек встречены на высотах от 480 до 1590 м над ур. м.
На Полярном Урале максимальная высота встречи пеночек-зарничек составляла 320 м над ур. м. у верхней границы леса в ерниковом травяно-моховом лиственничном редколесье [6]. В Вишерском заповеднике (Пермский край) на западной границе ареала гнездовые поселения обнаружены на высоте 580–740 м над ур. м. в редколесьях и криволесьях по границам горных лугов и верховых болот (Мещерягина С. Г., неопубликованные данные). В Хан-Хэнтэйском заповеднике описаны попытки гнездования пеночек-зарничек на высоте 1000 м над ур. м. в области лесостепей в северной части Монголии, представляющей южную границу ареала гнездования вида [25].
Находки гнезд тусклой зарнички Phylloscopus inornatus humei зафиксированы в северо-западной части Хамар-Дабана (Южное Прибайкалье) на высотах 1750–1800 м над ур. м. [8], а в горно-таежной части Западного Алтая — на высотах 1 000–2 100 м над ур. м. [24].
В зоне пространственного контакта форм humei и inornatus в центральной части Западного Саяна зарнички формы inornatus отмечены на высоте 1 300–1 600 м над ур. м. в пойменных ольхово-ивовых лесах и на открытых участках в таежном поясе [11].
Широкий диапазон высотного распределения гнездовых поселений пеночек-зарничек на территории Баргузинского заповедника определяет различия в сроках начала откладки яиц (рис. 3). Запаздывание сроков размножения в один и тот же сезон у зарничек, размножающихся в высокогорье по сравнению с прибрежно-равнинной и предгорной частью лесного пояса, отмечалось ранее А. А. Ананиным [2]. В 2013 г. гнездование в горнолесном и подгольцово-субальпийском поясе началось тогда, когда в прибрежно-равнинном и предгорном заканчивалось (разница в 15 суток).
-
В. К. Рябицев [23], рассматривая процесс формирования территориальной структуры одновидовых групповых поселений в условиях различной плотности, приходит к заключению, что у изученных на контрольных площадках в разные годы видов птиц схема формирования поселений очень сходна: сначала образовывалось рыхлое поселение, которое затем уплотнялось путем вселения новых особей.
Пространственно-временная последовательность возникновения гнезд во всех гнездовых поселениях на изучаемой территории в указанные периоды была подобна схеме, проиллюстрированной на примере участка 4 (рис. 4). Первые гнезда появлялись в центре и в периферийных частях потенциально-возможной площади заселения в гнездовом поселении, а затем происходило вселение на территории между и/или внутрь имеющихся гнездовых участков.
Таким образом, минимальные расстояния между вновь возникающими и имеющимися гнездами пеночек-зарничек в гнездовых поселениях варьировали в пределах от 15 до 571 м и зависели от характера вселения (табл. 4). Интересно, что в случае обособленного гнездования пеночек-зарничек за пределами основного ареала была зарегистрирована находка двух гнезд, сделанных в 60 м одно от другого. При этом в случае повторного гнездования (после разорения) одна пара отстроила гнездо в 40 м от предыдущего [14]. Для тусклой зарнички приводятся минимальные расстояния между двумя гнездами в Западном Танну-Ола — 11 м [13] и в Кунгей-Алатау — 15 м [10].
Выводы
На западном макросклоне Баргузинского хребта пеночки-зарнички Phylloscopus inornatus формируют пространственные группировки гнездовых поселений, различающихся социальной организацией (регулярные долгосрочные или многократные нерегулярные краткосрочные), но схожих в процессе формирования территориальной структуры. Кроме того, возможны случаи однократного гнездования. Долговременные изменения плотности населения вида отдельных пространственных выделов на постоянных учетных маршрутах и количества пар в постоянных гнездовых поселениях носят асинхронный характер, что определяется возможностью пространственного перераспределения в пределах высотных поясов и различных речных долин территории Баргузинского заповедника.
Список литературы Пространственно-временная структура гнездовых поселений пеночки-зарнички на западном макросклоне Баргузинского хребта
- Ананин А. А. Долговременные исследования динамики численности птиц Баргузинского хребта//Развитие современной орнитологии в Северной Евразии: тр. XII Междунар. орнитол. конф. Северной Евразии. -Ставрополь: Изд-во Ставропол. гос. ун-та, 2006а. -С. 280-297.
- Ананин А. А. Птицы Баргузинского заповедника. -Улан-Удэ: Изд-во Бурят. гос. ун-та, 2006б. -276 с.
- Ананин А. А. Многолетняя динамика летнего населения птиц высокогорий Баргузинского хребта (Северо-Восточное Прибайкалье)//Известия Самарского научного центра РАН. -2009. -Т. 11, № 1/1. -С. 9-12.
- Ананин А. А. Особенности долговременной динамики населения пеночки-зарнички на западном макросклоне Баргузинского хребта//Актуальные вопросы изучения птиц Сибири: материалы Сибирской орнит. конф., посвящ. памяти и 75-летию Э. А. Ирисова. -Барнаул: Азбука, 2010а. -С. 54-57.
- Ананин А. А. Птицы Северного Прибайкалья: динамика и особенности формирования населения. -Улан-Удэ: Изд-во Бурят. гос. ун-та, 2010б. -296 с.
- Головатин М. Г., Пасхальный С. П. Птицы Полярного Урала. -Екатеринбург: Изд-во Урал. ун-та, 2005. -С. 386-388.
- Доржиев Ц. З. Птицы Байкальской Сибири: систематический состав, характер пребывания и территориальное размещение//Байкальский зоол. журн. -2011. -Вып. 1(6). -С. 30-54.
- Дурнев Ю. А., Сонин В. Д., Сирохин И. Н. Орнитологические находки в северо-западной части Хамар-Дабана (южное Прибайкалье)//Русский орнит. журн. -2012. -Т. 21, экспресс-вып. 767. -С. 1416-1418.
- Ковшарь А. Ф. Певчие птицы в субвысокогорье Тянь-Шаня. -Алма-Ата, 1979. -312 с.
- Ковшарь А. Ф., Гаврилов Э. И., Родионов Э. Ф. О биологии тусклой зарнички Phylloscopus inornatus humei//Орнитология. -1974. -Вып. 11. -С. 253-267.
- Лисовский А., Рубцов А. Взаимоотношения обыкновенной и тусклой зарничек в зоне симпатрии на Западном Саяне//Актуальные проблемы изучения и охраны птиц восточной Европы и северной Азии: материалы междунар. конф. (XI орнитол. конф.). -Казань: Матбугат Йорты, 2001. -С. 378-379.
- Мальчевский А. С. Орнитологические экскурсии. Сер. Жизнь наших птиц и зверей. -Л.: Изд-во Ленингр. ун-та, 1981. -Вып. 4. -296 с.
- Марова И. М. Биология размножения и пространственная структура поселений тусклых зарничек в Западном Танну-Ола//Орнитология. -1990. -Вып. 24. -С. 128-130.
- Михайлов К. Е., Балацкий Н. Н. Гнездование пеночки-зарнички Phylloscopus inornatus на южной границе ареала в северо-восточном Приморье//Русский орнит. журн. -1997. -Т. 6, экспресс-вып. 19. -С. 8-13.
- Морозов В. В. Фаунистические находки на западном макросклоне Полярного Урала//Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. -Екатеринбург: Изд-во Урал. отд-я РАН, 1995. -С. 56-59.
- Морозов В. В. К орнитофауне Полярного Урала//Русский орнит. журн. -2003. -Т. 12, эспресс-вып. 212. -С. 3-5.
- Морозов В. В. Материалы по фауне птиц западного макросклона Полярного Урала//Орнитология. -2013. -Т. 38. -С. 85-105.
- Наумов Н. П. Этологическая структура популяций наземных позвоночных//Поведение животных: Экологические и эволюционные аспекты. -М.: Наука, 1972. -С. 37-39.
- Равкин Ю. С. К методике учета птиц в лесных ландшафтах//Природа очагов клещевого энцефалита на Алтае. -Новосибирск: Наука, 1967. -С. 66-75.
- Редькин Я. А., Коновалова М. В. К вопросу о репродуктивных отношениях двух форм зарнички Phylloscopus inornatus (Blyth, 1842) на основе анализа внешних морфологических признаков и деталей распространения//Русский орнит. журн. -2003. -Т. 12, Экспресс-вып. 247. -С. 1407-1430.
- Рябицев В. К. Территориальные отношения и динамика сообщества птиц в Субарктике -Екатеринбург: Наука, 1993. -296 с.
- Рябицев В. К. Птицы Урала, Приуралья и Западной Сибири: справ.-определитель. -Екатеринбург: Изд-во Урал. ун-та, 2008. -634 с.
- Рябицев В. К., Бачурин Г. Н., Шутов С. В. К распространению птиц на западном склоне Приполярного Урала//Фауна Урала и Европейского Севера. -Свердловск, 1980. -С. 54-59.
- Щербаков Б. В. Тусклая зарничка Phylloscopus humei на Западном Алтае//Русский орнит. журн. -2010. -Т. 19, экспресс-вып. 578. -С. 1067-1070.
- Biedermann P. H. W. Hidden leks in the Yellow-browed Warbler Phylloscopus inornatus? -Investigations from the Khan Khentey reserve (Mongolia)//Acrocephalus. -2006. -V. 27 (128-129). -С. 21-35.
- Bourski O. V. and Forstmeier, W. Does interspecific competition affect territorial distribution of birds? A longterm study on Siberian Phylloscopus warblers//Oikos. -2000. -N 88. -С. 341-350.
