Пространственное распределение почвенных нематод в ризосферах тополя и березы

Автор: Антропова Светлана Александровна, Карташев Александр Георгиевич
Журнал: Вестник Нижневартовского государственного университета @vestnik-nvsu
Рубрика: Экология животных
Статья в выпуске: 2, 2016 года.
Бесплатный доступ
Исследованы численность и родовой состав почвенных нематод в ризосферах тополя и березы. Пространственное распределение сообществ почвенных нематод изучалось в окрестностях г. Томска в ризосферах березы и тополя на расстоянии 20, 40, 60 и 80 см от корневой шейки. Нематод выделяли по модифицированному методу Бермана из навесок почвы в 15-30 г в 3-кратной повторности. Фиксатор - ТАФ (триэтаноламин + формалин + вода, в соотношении 2:7:91). Плотность популяций нематод рассчитывали на 100 г почвы. В ризосфере березы выявлены 11 родов нематод, относящихся к 4 трофическим группам, в ризосфере тополя - 9 родов нематод, относящихся к 5 трофическим группам. Доминирующей является группа нематод паразитов растений. Следующая трофическая группа нематод изучаемых деревьев представлена бактериотрофами. Высокая численность нематод-бактериотрофов, отмеченная в исследовании, - характерное явление для сообществ нематод лесных биотопов. Отмечена незначительная доля хищных нематод и нематод, ассоциированных с растениями. В ризосфере тополя отмечена группа нематод-политрофов. Изменения численности почвенных нематод в период исследования происходят за счет изменения степени увлажненности почвы. Максимальное количество почвенных нематод зарегистрировано на расстоянии 20 см от корневой шейки. Доминантный род нематод в изученных ризосферах - Meloidodera, субдоминантными являются рода Mononchus, Rhabditis, Plectus.
Почвенные нематоды, пространственное распределение, ризосфера
Короткий адрес: https://sciup.org/14116926
IDR: 14116926 | УДК: 592
Spatial distribution of soil nematodes in rhizosphere of poplar and birch
The following paper reflects the study of the number and generic composition of soil nematodes in poplar and birch root zones. The authors analyzed spatial distribution of soil nematode communities near in root zones of birches and poplars growing in forest areas near the city of Tomsk and located at a distance of 20, 40, 60 and 80 cm from the root collar. The researchers isolated nematodes applying the modified Berman technique to take 15-30 g of each soil sample three times and used triethanolamine + formalin + water in the ratio 2:7:91 as a fixative substance. The population density of nematodes was calculated per 100 g of soil. The studies revealed 11 nematode genera belonging to the 4 trophic groups in birch root zones and 9 nematode genera belonging to 5 trophic groups in poplar root zone. Plant parasites are the dominant group of nematodes. Another trophic group of nematodes is presented by bacteriotrophes. The study revealed an insignificant proportion of predatory nematodes and nematodes associated with plants, along with high levels bacteriotroph nematodes, which is a characteristic phenomenon of nematodes in forest habitats...
Текст научной статьи Пространственное распределение почвенных нематод в ризосферах тополя и березы
Нематоды — широко распространенная группа почвенных беспозвоночных животных с высокой численностью. Различные факторы среды оказывают влияние на распространение, численность и функциональную активность нематод (Randy, Anwar. Bilgrami 2004). Благодаря большому видовому разнообразию нематод влияние факторов на различные группы исследована недостаточна. Почвенные нематоды активно включаются в трофические цепи. Вследствие своей многочисленности, широкого распространения и приспособленности к различным источникам питания почвенные нематоды осуществляют значительные биогеоценотические функции. Совместно с почвенной микрофлорой они участвуют в разложении и преобразовании органического вещества, делая его доступным для растений, грибов и бактерий (Козловская 1976; Парамонов 1962). Нематоды регулируют численность грибов и бактерий, как подавляя, так и стимулируя их размножение и развитие. Наиболее заселена нематодами ризосфера деревьев, в которой обеспечивается контакт нематод с корневой системой и органами растения, погруженными в почву
(Парамонов 1962). Пространственное распределение сообществ нематод покрытосеменных деревьев исследовано недостаточно.
Целью наших исследований являлось исследование пространственного распределения сообществ нематод в ризосфере берёзы и тополя в летний период.
Методика исследований. Исследования проводились в окрестностях г. Томска, в подтаежной зоне Западной Сибири в весенне-летний период: май, июнь, июль. Образцы почвы для исследования численности и родового состава почвенных нематод отбирались в светло-серых лесных почвах. Для исследования распределения почвенных нематод выбран берёзово-зеленомошно-разнотравный лес. Древесный ярус образован преимущественно березой пушистой (Betula pubescensL) и тополем (Рopulus L.\ Для изучения особенностей биотопического распределения почвенных нематод заложены модельные площадки размером 10x10 м. На участках брались пробы из середины генетических горизонтов почвы. В ризосфере деревьев по поверхности закладывались площадки на расстоянии 20, 40, 60,
80 см от корневой шейки деревьев (Дюкарев и др. 1997; Соколов 1968).
Нематод выделяли по модифицированному методу Бермана из навесок почвы в 15—30 г в 3-кратной повторности. Фиксатор — ТАФ (триэтаноламин + формалин + вода, в соотношении 2:7:91). Плотность популяций нематод рассчитывали на 100 г почвы. Устанавливали систематическую принадлежность 100 особей из пробы. Идентификацию нематод проводили до рода. Каждый таксон нематод относили к одной из шести эколого-трофических групп: бактериотро-фы (Б), микотрофы (М), политрофы (П), хищники
Матвеева и др. 2008: 65; Yeates G.W. et al 1993). Для определения влияния факторов окружающей среды на сообщество нематод рассматривали: тип почвы, влажность, pH почвенной среды, как наиболее важные показатели, влияющие на жизнедеятельность почвенной микробиоты. Полученные результаты исследований обрабатывались в табличном процессоре Microsoft ® Excel 2010 и с использованием пакета Statistica 7.1.
Результаты и обсуждения. В результате проведенных исследований в ризосферах изученных деревьев выявлено 11 родов нематод. Результаты численности и родового состава почвенных не-
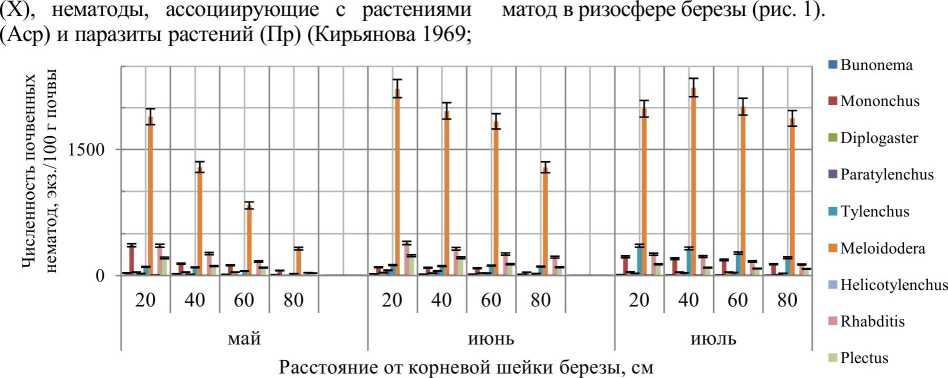
Рис. 1. Пространственное распределение родов сообществ почвенных нематод в зависимости от расстояний до корневой шейки берёзы
В ризосфере березы обнаружены 9 родов нематод: Випопета, Mononchus, Diplogaster, Paratylenchus, Tylenchus, Meloidodera, Helicotylenchus, Rhabditis, Plectus. Трофическая структура в ризосфере березы представлена 4 группами. Преобладающей является группа нематод паразитов растений, которая составляет 65% нематодного сообщества. Основная масса представлена родом нематод Meloidodera. Ме-лойдодеры регистрировались многими учеными как седентарные эндопаразиты корней древесной растительности. Численность данного рода на расстоянии 20 см составляет 1890 экз./100 г почвы, что свидетельствует о неблагоприятном влиянии нематод на рост и развитие деревьев. Следующая трофическая группа ризосферы представлена бактреотрофами, которые составляют 21%. Высокая численность нематод-бактериотрофов, отмеченная в исследовании, — характерное явление для сообществ нематод лесных биотопов (Матвеева и др. 2008; Armendariz, Arpin 1996; Tomar, Ahmad 2009). Доминирование бактериотрофов во всех типах сообществ северного региона — типичное явление, т.к. они наряду с прочими почвенными микроорганизмами являются в условиях севера основными участни ками процесса разложения растительных остатков и опада, способствуют их минерализации и гумификации (Кудрин 2011). Одной из причин высокого обилия бактериотроф может являться большее видовое разнообразие водорослей, бактерий и других микроорганизмов в ризосфере березы, по сравнению с еловыми, сосновыми и кедровыми насаждениями (Гарибова и др. 1978). Следует отметить незначительную долю хищных нематод (10%) и нематод, ассоциированных с растениями (4%). Отсутствие нематод-микотро-фоф можно объяснить тем, что береза является слабо микотрофной древесной породой.
Анализ данных, представленных на рис. 1, позволяет выявить изменение численности почвенных нематод в зависимости от расстояния от корневой шейки березы. У березы мочковатая корневая система, но корни залегают неглубоко. Корни дерева постоянно нуждаются в наличии влаги, поэтому вокруг растений находится небольшое число других представителей растений. Дерево вытягивает влагу и питательные вещества (Рахтеенко 1952). Возможно, данные факторы влияют на численный и родовой состав почвенных нематод. Наибольшая численность почвенных нематод наблюдается на рас- стоянии 20 см от корневой шейки. Максимальное значение численности характерно для рода Meloidodera, минимальное — для рода Helicotylenchus, присутствие которого отмечается на расстоянии 20 см от корневой шейки дерева. В июне происходит сокращение родов нематод в связи с выпадением рода Helicotylenchus. Доминирующий род Meloidodera увеличивает численность в пределах всей обследуемой зоны. Для других родов характерно увеличение численности простейших с максимальными значениями в области 20 см от корневой шейки дерева за счет изменения влажности. Данные по изменению показателей влажности представлены в таблице. По данным исследователей, отмечается изменение численности почвенных нематод в зависимости от градиента влажности. Выявлены рода нематод, которые реагируют на изменение влажности почв. Так, рода: Tylenchus, Rhabditis, Plectus, Paratylenchus снижают численность при увлажнении. Для рода Mononchus характерна прямая зависимость изменения численности с возрастанием влажности (Кудрин 2012).
Анализ данных пространственного распределения почвенных нематод в июле позволяет наблюдать смещение максимальной численности доминирующего рода к 40 см от корневой шейки. Частично восстанавливается род Helicotylenchus. Наблюдается активное распространение нематод на расстояния 60 и 80 см от корневой шейки дерева. Что, вероятно, связано с активными биологическими процессами в ризосфере берёзы.
Таблица 1
Физико-химические показатели почвы
|
Физико-химические показатели |
май |
|
|
Береза |
Тополь |
|
|
Влажность, % |
42,8 |
41,5 |
|
pH |
6,04 |
6,69 |
|
Гранулометрический состав |
суглинок легкий |
супесь |
|
июнь |
||
|
Влажность, % |
9,27 |
11,97 |
|
рн |
6,07 |
6,08 |
|
Гранулометрический состав |
суглинок легкий |
супесь |
|
июль |
||
|
Влажность, % |
19 |
19,5 |
|
pH |
5,74 |
6,15 |
|
Гранулометрический состав |
суглинок легкий |
супесь |
Meloidodera является наиболее многочисленным и составляет 62% нематодоценоза. Аналогично с ризосферой березы можно сделать вывод, что данный род нематод является доминирующим или свидетельствующим о болезни исследуемых деревьев. Второй группой нематодного сообщества являются бактериотрофы, составляющие 16%. Следует отметить присутствие хищных нематод (11%) и нематод, ассоциированных с растениями (7%). Выявленная структура сообществ аналогична трофической структуре ризосферы березы. Необходимо отметить наличие представителей еще одной группы нематод — политро-фов, которая присутствует в ризосфере тополя и составляет 4%. Для политрофов статистически значимым является влияние кислотности почвы, политрофы предпочитают менее кислые условия (Кудрин 2012).
Анализ данных (рис. 2) позволяет выявить изменение численности почвенных нематод в зависимости от расстояния до корневой шейки тополя. Известно, что корни тополя мощные, находятся на поверхности и распространяются на расстояние 3—4 м. Анализ данных по пространственному распределению почвенных нематод в ризосфере тополя позволяет выявить появление новых родов, не присутствовавших в ризосфере березы: Dorylaimus, Lelenchus. Доминирующим родом является Meloidodera. Максимальная численность нематод наблюдается на расстоянии 20 см от корневой шейки тополя.
В результате проведенных исследований можно сделать следующие выводы: в ризосфере берёзы выявлены 11 родов нематод, относящихся к 4 трофическим группам; к доминантным родам нематод относятся: Meloidodera, к субдоминант- ным: Mononchus, Rhabditis. Максимальная чис- течение летнего сезона на расстоянии от 20 до 60 ленность нематод пространственно смещается в см от корневой шейки дерева.
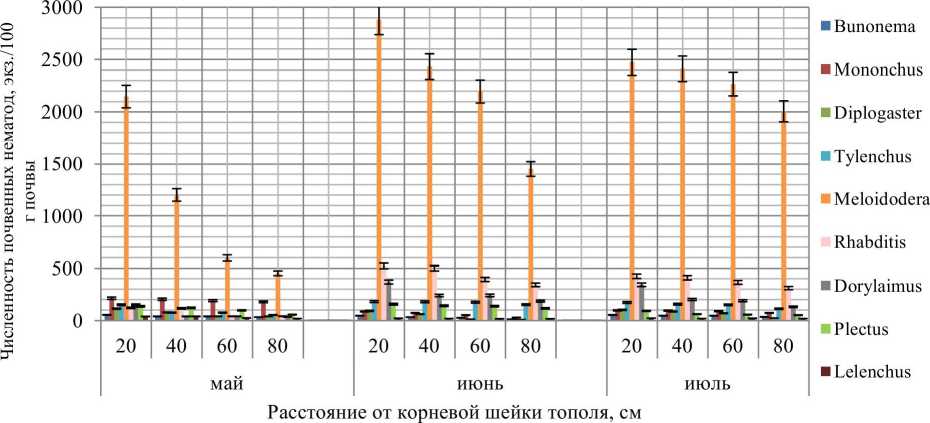
Рис. 2. Пространственное распределение родов сообществ почвенных нематод в зависимости от расстояний до корневой шейки тополя
В ризосфере тополя обнаружено 9 родов нематод, относящихся к 5 трофическим группам. К доминантным относятся: Meloidodera, к субдо минантным: Mononchus, Plectus. Максимальная численность нематод наблюдается в 20 см от корневой шейки дерева.
Список литературы Пространственное распределение почвенных нематод в ризосферах тополя и березы
- Гарибова Л. В., Дундин Ю. К., Коптяева Т. Ф., Филин В. Р. 1978. Водоросли, лишайники и мохообразные СССР. Москва: Мысль.
- Кирьянова Е. С., Кралль Э. Л. 1971. Паразитические нематоды растений и методы борьбы с ними. Москва: Наука, Т. 1.
- Козловская Л. С. 1976. Роль почвенных беспозвоночных и трансформации органического вещества болотных почв. Москва: Наука.
- Кудрин А. А. 2012. Разнообразие и экология почвенных нематод в пойменных экосистемах подзон средней и северной тайги Республики Коми: Автореф.. дис. кан. биол. наук. Сыктывкар.
- Кудрин А. А. 2011. Структура населения нематод в градиенте влажности пойменных осиново-березовых лесов республики Коми//Известия ПГПУ им. Белинского 25, 358-363.
- Матвеева Е. М., Груздева Л. И., Коваленко Т. Е., Сущук А. А. 2008. Почвенные нематоды как биоиндикаторы техногенного загрязнения таежных экосистем//Тр. КарНЦ РАН. Cерия Б. «Биогеография Карелии». Вып. 14, 63-75.
- Парамонов А. А. 1962. Основы фитогельминтологии. Москва: АН СССР. Т. 1.
- Дюкарев А. Г., Пологова Н. Н., Лапшина Е. Д. 1997. Природно-ресурсное районирование Томской области/Воробьев В. Н. (отв. ред.). Томск: Спектр.
- Рахтеенко И. Н. 1952. Корневые системы древесных и кустарничковых пород. Москва: Гослесбумиздат.
- Соколов А. В. 1968. Агрохимическая характеристика почв СССР: Районы Западной Сибири. Москва: АН СССР.
- Armendariz I., Arpin P.1996. Nematodes and their relationship to forest dynamics: I. species and trophic groups//Biol Fertil Soils. Vol. 23, 405-413.
- Gaugler R., Bilgrami A. L. 2004. Nematode Behaviour. Wallingford: CABI.
- Tomar V. V. S, Ahmad W. 2009. Food web diagnostics and function diversity of soil inhabiting nematodes in a natural woodland//Helminthologia. Vol. 23, 405-413.
- Yeates G. W., Bongers T., De Goede R. G. M., Freckman D. W., Georgieva S.S. 1993. Feeding habits in soil nematode families and genera -an outline for soil ecologists//J Nematol. Vol. 25, 315-331.
