Протеолитическая регуляция экспрессии ростовых факторов и HIF-1 при раке эндометрия

Автор: Спирина Л.В., Юнусова Н.В., Кондакова И.В., Коломиец Л.А., Коваль В.Д., Чернышова А.Л., Шпилева О.В.
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 2 (50), 2012 года.
Бесплатный доступ
Проведено исследование связи химотрипсинподобной активности протеасом и содержания ИФР-I, ИФР-II, PAPP-A, VEGF и HIF-1 в тканях рака эндометрия. Показано, что тотальная активность протеасом и активность 20S и 26S пулов протеасом в злокачественных опухолях значительно выше, чем в неизмененном эндометрии. Обнаружены корреляционные связи между ак- тивностью пулов протеасом и экспрессией ИФР-I и HIF-1 в опухолевой ткани, которые свидетельствуют о возможной регуляции содержания ростового и транскрипционного факторов протеасомами. Поскольку считается, что основной пул ИФР-I находится во внеклеточном пространстве, то, вероятно, в регуляции содержания ИФР-I также принимают участие внеклеточные протеасомы. Положительные корреляционные связи между содержанием ИФР- I, HIF-1, VEGF и металлопротеиназой PAPP-A дают основание считать, что этот протеолитический фермент является другим важным регулятором содержания ростовых и транскрипционного факторов. Представленные данные показывают возможность протеолитической регуляции ростовых и ядерных факторов, что может играть важную роль в патогенезе злокачественных новообразований.
Инсулиноподобные факторы роста, фактор роста эндотелия сосудов, протеасома, рак эндометрия
Короткий адрес: https://sciup.org/14056202
IDR: 14056202
Protelytic regulation of growth factors and HIF-1 in endometrial cancer
We studied the correlation between chymotrypsin-like activity of proteasomes and IGF-I, IGF-II, PAPP-A, VEGF, HIF-1 levels in endometrial cancer tissues. The total activity of proteasomes and the activity of the 20S and 26S proteasomes were shown to be significantly higher in the malignant tumors than in the normal endometrium. The relationships between the proteasome pool activity and IFG-I and HIF-1 expressions were found. The data obtained indicate the possible proteasome regulation of growth and transcription factors. The major pool of IGF-I is located in the extracellular space, and it is likely that extracellular proteasomes also take part in the regulation of the IGF-I content. The positive correlation between the IGF-I, HIF-1, VEGF levels and PAPP-A metalloproteinase give evidence that this proteolytic enzyme is one more important regulator of growth and transcription factors. The present data show the evidence of proteolytic regulation of growth and nuclear factors that can play an important role in cancer pathogenesis.
Текст научной статьи Протеолитическая регуляция экспрессии ростовых факторов и HIF-1 при раке эндометрия
Особенности роста, развития и метастазирования злокачественных опухолей определяются ростовыми и транскрипционными факторами, среди которых одними из наиболее значимых являются активируемый гипоксией транскрипционный фактор (HIF-1), HIF-индуцируемые инсулиноподобные факторы роста (ИФР) и фактор роста эндотелия сосудов (VEGF). В настоящее время сформировано понятие об ИФР-системе, в которую входят ИФР-I и ИФР-II, рецептор ИФР первого типа, а также 6 белков, связывающих инсулиноподобные факторы роста (IGFBPs). Кроме того, функционально с этой системой связаны протеиназы, расщепляющие IGFBPs, среди которых важное значение отводят белку, ассоциированному с беременностью – PAPP-A (pregnancy-associateds plasma protein) [2].
Все компоненты ИФР-системы продуцируются стромальными клетками нормального, гиперплазированного и малигнизированного эндометрия, а также рядом опухолей эпителиального происхождения [2, 8, 14, 16]. Другим важнейшим ростовым фактором, участвующим в развитии и прогрессии рака эндометрия является VEGF. Семейство сосудистых эндотелиальных факторов роста представлено 5 членами: VEGF-A, VEGF-B, VEGF-C, VEGF-D и плацентарным ростовым фактором. Процесс неоангиогенеза обеспечивает фактор VEGF-A, который и называют VEGF [17].
Ростовые факторы, в том числе и ИФР, индуцируют активируемый гипоксией транскрипционный фактор – HIF-1, так называемый независимый от гипоксии путь активации HIF-1 [7, 11]. На примере клеточной линии глиобластомы показана возможность инактивации сигнального пути ИФР-STAT3-HIF1 при использовании ингибиторов рецептора ИФР [12]. C другой стороны, ИФР-II, IGFBP-2, -3 и VEGF являются HIF-индуцируемыми генами [11, 22]. HIF-1 является гетеродимером, состоящим из α- и β-субъединиц. β-cубъединицы фактора являются конститутивными, активность фактора зависит от экспрессии и посттрансляционных модификаций α-субъединиц. При гидроксилировании остатков пролина и аспарагина наблюдается связывание HIF-1α с белком фон Хиппель-Линдау (pVHL), что сопровождается прикреплением цепочки убиквитина к белку и его последующей деградацией в протеасоме [22, 26]. В условиях гипоксии происходит стабилизация HIF-1α и формирование функционально активных гетеродимеров с HIF-1β с повышением ДНК-связывающей активности транскрипционного фактора.
Необходимо отметить, что регуляторным этапом многих физиологических и патологических процессов является внутриклеточная деградация белков в протеасомах. Основным компонентом убиквитин-протеасомной системы являются про-теасомы – мультисубъединичные комплексы, включающие каталитическое ядро 20S, к которому присоединена одна или две регуляторные частицы. Если по крайней мере одной из этих частиц является РА700 (19S регуляторная частица), то образуется 26S протеасома, осуществляющая, главным образом, АТФ- и убиквитин-зависимый протеолиз большинства клеточных белков. Если же в роли регуляторной частицы выступает другой белковый комплекс (РА28, РА200), то такая ассоциация представляет собой активированную 20S протеасому, которая деградирует малые, аномальные и коротко живущие белки [5]. 20S протеасомы представлены иммунными и конститутивными формами, образующимися при сочетании конститутивных (α1α2α3α4α5α6α7) или иммунных (LMP 2, LMP 7, MECL-1) протеолитических субъединиц. Замена конститутивных субъединиц на иммунные приводит к образованию модифицированных форм протеасом и сопровождается изменением их специфичности, вследствие чего они продуцируют иммуногенные пептиды для их презентации главным комплексом гистосовместимости. В то же время известно, что иммунные протеасомы могут выполнять неиммунные функции. Встроенные иммунные субъединицы приводят к изменению активности протеасом [3]. Относительное содержание протеасом в клетке, их состав и активность изменяются в соответствии с потребностями клетки и условиями, в которых она находится, что становится возможным благодаря взаимодействию протеасом с большим количеством белков, а также благодаря механизмам регуляции на уровне транскрипции, которые на данный момент остаются мало изученными [5].
Другой аспект протеолитической регуляции экспрессии ряда белков связан с металлопротеиназами, действующими в основном экстраклеточно. Металлопротеиназа PAPP-A относится к подсемейству паппализинов, и ее основной функцией является ИФР-зависимый протеолиз, главным образом IGFBP-4, но также IGFBP-5 и IGFBP-2. Таким образом, PAPP-A участвует в регуляции биодоступности ИФР. Единственным ингибитором PAPP-A является проформа большого основного белка эозинофилов (pro-MBP) [14]. В настоящее время очень мало известно о молекулярной регуляции IGFBP-4 протеазной системы, однако есть данные о сочетанном повышении экспрессии PAPP-A и снижении pro-MBP в клеточных линиях при инкубации с трансформирующим фактором роста бета. Другим регулятором этой системы является ИФР-II, действующий на посттрансляционном уровне [19].
Имеется определенная взаимосвязь между HIF-1, ростовыми факторами и активностью протеасом в клетках. Передача сигнала с активированного рецептора ИФР осуществляется через вну- триклеточные субстраты инсулинового рецептора (IRS), а уровень протеасомной деградации IRS-2 влияет на степень активации ИФР-зависимого сигнального пути [21]. Также было выявлено, что разрушение самого рецептора ИФР происходит в протеасомах [13]. Показано, что в опухолях достаточно много факторов регулируют стабилизацию HIF-1α, однако роль протеасом в этом процессе также существенна [22]. Так, снижение деградации HIF-1α при использовании ингибиторов протеа-сом или при гипоксии приводит к значительному повышению экспрессии как мРНК VEGF, так и самого белка в опухолевых клетках [17, 25]. Таким образом, возможно опосредованное участие про-теасом в механизмах развития опухоли через изменение активности транскрипционных факторов и соответствующее изменение синтеза ростовых факторов. В настоящее время протеолитическая регуляция экспрессии ростовых факторов и HIF, их взаимное влияние в злокачественных опухолях человека изучены недостаточно. Проведение таких исследований особенно актуально в отношении рака эндометрия, так как ИФР и VEGF играют в развитии и прогнозе данной патологии ведущую роль [2, 8, 17].
Целью исследования явился сравнительный анализ содержания компонентов системы ИФР, VEGF, HIF-1α, тотальной активности протеасом, активности их пулов и определение возможного регуляторного влияния отдельных компонентов системы вне- и внутриклеточного протеолиза на экспрессию ростовых и транскрипционного факторов.
Материал и методы
В исследование вошли больные с морфологически верифицированным раком эндометрия (РЭ) I–II стадии (n=49, средний возраст 56,8 ± 1,5 года), из них Iа стадию заболевания имели 13 больных, Ib – 24 , Iс – 3 пациентки, II стадию – 9 больных. Объемы диагностики и лечения больных раком эндометрия соответствовали рекомендуемым алгоритмам объемов диагностики и лечения злокачественных новообразований, утвержденным Министерством здравоохранения и социального развития РФ. Проведение данной работы одобрено этическим комитетом НИИ онкологии СО РАМН. При оценке тканевого уровня ИФР, IGFBPs и PAPP-A была сформирована группа сравнения из 20 больных с гиперпластическими процессами эн- дометрия (ГПЭ). У 5 больных гистологически была подтверждена атипичная гиперплазия эндометрия, у 15 – сложная типичная гиперплазия эндометрия. Средний возраст этих больных составил 52,8 ± 4,5 года. Образцы гиперплазированного эндометрия получали при выполнении гистероскопии с биопсией или радикальной операции. Материалом для исследования протеасом явились образцы опухолевой и гистологически не измененной ткани, находящиеся на расстоянии не менее 1 см от границы опухолей, полученные при выполнении радикального оперативного вмешательства, которые после взятия замораживались и хранились при –80 оС.
Получение осветленных гомогенатов. Замороженную ткань (100 мг) гомогенизировали в жидком азоте, затем ресуспендировали в 300 мкл 50 мМ трис-HCl буфера (pH=7,5), содержащего 2 мМ АТФ, 5 мМ хлорид магния, 1 мМ дитиотреитол, 1мМ ЭДТА и 100 мМ хлорид натрия. Гомогенат центрифугировали 60 мин при 10000g и 4оС.
Фракционирование протеасом. Все процедуры проводили при 4°С. Белки осветленных гомогенатов фракционировали с помощью сульфата аммония в два этапа. Фракцию, обогащенную 26S-протеасомами, получали добавлением сульфата аммония до 40 % насыщения, фракцию 20S-протеасом – добавлением сульфата аммония до 70 % насыщения [1]. В полученных фракциях определяли активность протеасом.
Определение активности протеасом. Химо-трипсинподобную активность тотального пула протеасом, 26S и 20S пулов протеасом определяли в осветленных гомогенатах опухолевых и неизмененных тканей, а также во фракциях протеасом, по гидролизу флуорогенного олигопептида N-Succi-nyl-Leu-Leu-Val-Tyr-7-Amido-4-Methylcoumarin, утилизирующегося химотрипсинподобными центрами протеасом [6], на флуориметре «Hitachi-850» (Япония) при длине волны возбуждения 380 нм и эмиссии 440 нм. Реакционная смесь для определения активности 20S протеасом содержала 20 мМ трис-HCl (pH=7,5), 1 мМ дитиотреитол, 30 мкМ N-Succinyl-Leu-Leu-Val-Tyr-7-Amido-4-Methylcou-marin. Для определения активности 26S протеасом в реакционную смесь дополнительно вводили 5 мМ хлорида магния и 1 мМ АТФ. Реакцию проводили при 37°С в течение 20 мин и останавливали 1 % додецил сульфатом натрия. Для оценки активности примесных протеаз в образцах применяли специфический ингибитор протеасом – MG132. Удельную активность протеасом выражали в единицах активности на 1 мг белка. Содержание белка определяли по методу Лоури.
Определение содержания ИФР, связывающих их белков, PAPP-A, VEGF и HIF-1α. Образцы осветленных гомогенатов опухолей использовались для определения тотального (внутриклеточного и внеклеточного) пула свободных ИФР-I, ИФР-II, а также IGFBP-3, IGFBP-4, VEGF и металлопротеиназы PAPP-A с использованием наборов для твердофазного иммуноферментного анализа (R&D Systems, DSL, США) на ИФА-анализаторе «Anthos 2020». Экспрессию HIF-1α субъединиц определяли с помощь наборов Caymanchem (США), ELISA. Приготовление и очистка ядерных экстрактов тканевого гомогената проводились в соответствии с рекомендациями фирмы-производителя. Уровень белка в гомогенатах и ядерных экстрактах определялся по Лоури. Результаты определения содержания ростовых факторов, связывающих их белков и PAPP-A выражали в нг/мг белка, а HIF-1α – в условных единицах на мг белка в лунке.
Статистическая обработка. Статистическую обработку результатов проводили с применением пакета статистических программ Statistica 6.0. В зависимости от вида распределения результаты представлены как M±m, где М-среднее выборочное, m – ошибка среднего или как медиана с интерквартильным размахом (25-й и 75-й процентили). Значимость различий исследовали с помощью t-критерия Стьюдента или критерия Манна-Уитни. Корреляционный анализ был проведен с использованием непараметрического критерия Спирмена.
Результаты и обсуждение
При изучении уровня ИФР, IGFBPs и PAPP-A в опухолях эндометрия и в гиперплазированном эндометрии выявлено, что уровень обоих ростовых факторов и IGFBP-4 был статистически значимо ниже в опухолевой ткани по сравнению с гиперплазированной (таблица). Анализируя полученные результаты, можно отметить, что в литературе отсутствуют сравнительные данные об уровне параметров ИФР-системы, за исключением экспрессии рецептора ИФР первого типа, при гиперпластических процессах и раке эндометрия. Имеются лишь данные о сходном уровне ИФР-I и его рецептора в опухолях эндометрия и соответствующей нетранс-формированной ткани [2]. Снижение уровня обоих ИФР и IGFBP-4 в опухолях эндометрия может быть обусловлено либо снижением синтеза этих факторов, повышением их деградации или увеличением фракций, связанных с IGFBPs ИФР.
Содержание другого ростового фактора (VEGF) коррелировало с глубиной инвазии опухоли в миометрий. У больных с инвазией в пределах слизистой и с инвазией опухоли до середины миометрия уровень ростового фактора составил 153,9 ± 40,1 пг/мг белка, а у больных с Iс стадией заболевания – 321,0 ± 60,1 пг/мг белка. При этом уровень экспрессии HIF-1α не зависел от стадии процесса и глубины инвазии опухоли в миометрий и составил в среднем 17,2 ± 3,3 УЕ/мг белка в лунке.
Выявлено, что в ткани рака эндометрия происходит увеличение тотальной активности про-теасом в 2,7 раза по сравнению с неизмененной тканью (164,1 ± 25,6 × 103 МЕ/мг белка и 59,5 ± 5,0 х 103 МЕ/мг белка соответственно). Активность пула 26S протеасом повышалась в 1,9 раза по сравнению с неизмененной тканью, активность пула 20S протеасом увеличивалась в 2,6 раза (рис. 1). Вероятно, увеличение активности про-теасом в опухоли по сравнению с неизмененными тканями связано с тем, что при опухолевом росте все процессы, происходящие в клетке, очень интенсивны. Опухолевый рост сопровождается
Уровень инсулиноподобных факторов роста, связывающих их белков и РАРР-А в гиперплазированном и малигнизированном эндометрии, Me (25–75%)
Таблица
|
Группы больных |
ИФР-I, нг/мг белка |
ИФР-II, нг/мг белка |
IGFBP-3, нг/мг белка |
IGFBP-4, нг/мг белка |
PAPP-A, нг/мг белка |
|
ГПЭ |
0,53 (0,29–1,25) |
8,49 (4,92–17,74) |
3,14 (1,02–11,1) |
1,11 (0,39–1,46) |
5,93 (2,00–10,9) |
|
РЭ |
0,32 (0,16–0,55)* |
5,05 (3,66–7,36)* |
3,80 (1,90–6,98) |
0,51 (0,22–0,83)* |
6,42 (1,38–11,3) |
Примечание: * – различия статически значимы по сравнению с группой больных с ГПЭ, p<0,05. СИБИРСКИЙ ОНКОЛОГИЧЕСКИЙ ЖУРНАЛ. 2012. № 2 (50)

□ опухолевая ткань ■ неизмененная ткань
Рис. 1. Активность протеасом в ткани рака эндометрия и неизмененной ткани. Примечание: * – различия статистически значимы по сравнению с неизмененной тканью (р<0,05)
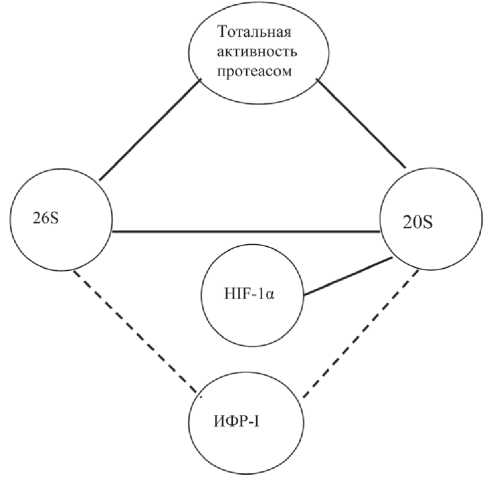
Рис. 2. Предполагаемая схема регуляции экспрессии HIF-1 и ИФР протеасомами при раке эндометрия. Примечание: 20S и 26S – пулы протеасом; ИФР- I – инсулиноподобные факторы роста-I;
HIF-1 – активируемый гипоксией транскрипционный фактор нарушением клеточного цикла, усилением пролиферации, ингибированием апоптоза. Несомненно, чтобы клетке продолжать свое существование, необходимо усилить систему, занимающуюся деградацией выполнившего свою функцию белкового и пептидного материала.
При проведении корреляционного анализа в опухолях эндометрия были выявлены взаимосвязи между активностью всех основных пулов про-теасом: между 20S и 26S пула протеасом (r=0,60; p<0,05), между тотальной протеасомной активностью и 20S пула протеасом (r=0,77; p<0,05), между тотальной протеасомной активностью и 26S пула протеасом (r=0,59; p<0,05). Активность 20S пула протеасом коррелировала с экспрессией HIF-1α (r=0,39; p<0,05). Таким образом, в опухолях эндометрия увеличение активности протеасом приводит к повышению экспрессии HIF-1α. Также получены отрицательные корреляционные взаимосвязи между экспрессией ИФР-I и активностью 20S и 26S пула протеасом (r1=–0,47; r2=–0,49; p<0,05) соответственно (рис. 2).
Выявленная прямая корреляционная зависимость между активностью пулов 26S и 20S протеа-сом при раке эндометрия может быть объяснена как через активацию транскрипции, так и с помощью внегеномных механизмов регуляции протеолиза. Влияние белков, участвующих в сборке протеасом и в контроле протеасом-зависимого протеолиза, обусловлено воздействием многих факторов и распространяется на оба пула протеасом [24]. Известно, что протеасомы принимают участие в pVHL-зависимой деградации HIF-1α. В условиях гипоксии происходит стабилизация HIF-1α, снижение степени деградации фактора и формирование функционально активных гетеродимеров с HIF-1β с повышением ДНК-связывающей активности транскрипционного фактора, с чем часто связывают повышение экспрессии HIF-1-индуцированных генов в опухолях. Однако при раке эндометрия имеет место сочетанное повышение тотальной протеасомной активности и ее пулов и повышение экспрессии HIF-1α. При этом необходимо отметить существование так называемого независимого от гипоксии пути активации HIF-1. Стабилизация HIF-1 и, соответственно, увеличение количества HIF-1α в ядерных экстрактах могут происходить при активации рецепторных тирозинкиназ при связывании с соответствующими ростовыми факторами, прежде всего с эпидермальным фактором роста и ИФР [22]. Также есть литературные данные о наличии альтернативных протеасом-независимых путей деградации HIF-1α другими протеолитическими системами, прежде всего кальпаин-опосредованной [26]. Вероятно, при раке эндометрия активация HIF-1α происходит преимущественно альтернативным путем.
Экспрессия ИФР является как HIF-1, так и NF-kappaB-зависимой. С учетом полученных собственных результатов и данных литературы об отсутствии повышенной экспрессии IGFBPs
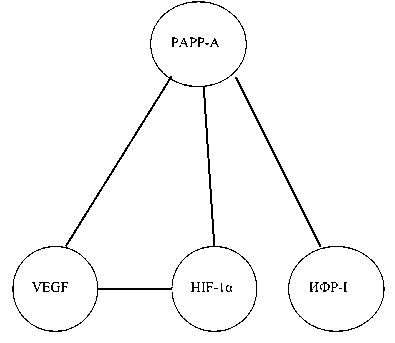
Рис. 3. Предполагаемая схема регуляции экспрессии ростовых факторов и HIF-1 металлопротеиназой PAPP-A при раке эндометрия. Примечание: ИФР- I – инсулиноподобный фактор роста-I; PAPP-A (pregnancy-associateds plasma protein-A) – протеиназа, расщепляющая IGFBP-2, -4 и -5; HIF-1α – α-субъединица активируемого гипоксией транскрипционного фактора; VEGF – фактор роста эндотелия сосудов в опухолях эндометрия, о повышении в опухолях эндометрия тотальной протеасомной активности и ее пулов с параллельным повышением экспрессии HIF-1 и снижении экспрессии NF-kappaB (p65) [4], об активации NF-kappaB при использовании ингибиторов протеасом в клеточных линиях рака эндометрия [10] можно сделать предположение о NF-kappaB-зависимом снижении синтеза обоих ИФР в опухолях эндометрия.
Выявленные отрицательные корреляционные взаимосвязи между экспрессией ИФР-I и активностью 20S и 26S пула протеасом, по-видимому, не могут считаться прямыми, поскольку действие протеасом на ростовые факторы и связывающие их белки, по данным большинства авторов, опосредуется через транскрипционные факторы, в том числе и через NF-kappaB и HIF-1 [9, 20]. В то же время существуют данные о присутствии протеасом, представленных, главным образом, 20S пулом, во внеклеточных средах организма. Поскольку остается открытым вопрос о наличии таких протеасом в различных тканях, их строении и функциях за пределами клетки, нельзя исключать участие внеклеточного пула протеасом в деградации экстраклеточного пула ИФР-I [15, 23].
Экспрессия HIF-1α в опухолях эндометрия была взаимосвязана не только с активностью пула протеасом, но и с экспрессией металлопротеиназы
PAPP-A (r=0,72; p<0,05). HIF-1α-зависимая экспрессия VEGF также была связана с экспрессией PAPP-A. Кроме того, выявлена корреляция между экспрессией PAPP-A и ИФР-I, r=0,52; p<0,05 (рис. 3).
Положительная корреляционная связь между содержанием ИФР-I и металлопротеиназой PAPP-A дает основание считать, что этот протеолитический фермент является другим важным регулятором содержания ростового фактора, осуществляя расщепление белков, связывающих ИФР- I, и тем самым повышая его содержание в тканях. Предполагаемая регуляция экспрессии HIF-1 и VEGF металлопротеиназой PAPP-A практически не изучена. Однако появились работы, в которых обсуждается ИФР-независимый антиангиоген-ный и противоопухолевый эффект IGFBP-4 [18]. C учетом того, что доминирующим субстратом PAPP-A является IGFBP-4 [14], становится понятной взаимосвязь PAPP-A с уровнем экспрессии VEGF в опухолях эндометрия. Кроме того, в исследованиях M.S. Moreno et al. (2006) показано, что IGFBP-4 является ингибитором ангиогенных эффектов VEGF и других ростовых факторов [18]. Интересно, что в образцах опухолей эндометрия не выявлено статистически значимых связей между экспрессией PAPP-A и ИФР-II. В то же время в культуре остеобластов, продуцирующих значительные количества ИФР, IGFBP-4 и PAPP-A, выявлено регулирующее влияние ИФР-II на протеолитическую активность PAPP-A [19].
Заключение. Таким образом, впервые проанализированы взаимосвязи содержания ИФР, VEGF и HIF-1α с химотрипсинподобной активностью про-теасом и экспрессией металлопротеиназы PAPP-A. Установлено, что протеасомы могут играть важную роль в регуляции содержания HIF-1α и ИФР-I в тканях рака эндометрия. Следует отметить, что если на фактор транскрипции протеасомы оказывают непосредственное регулирующее влияние, то на ИФР- I оно, вероятно, опосредовано через NF-kappaB и HIF-1. Нельзя исключить также расщепление ИФР-I во внеклеточных протеасомах. Другим важным протеолитическим регулятором содержания ИФР-I, VEGF, а возможно и HIF-1, в клетках является металлопротеиназа PAPP-A. Поскольку HIF-1 и ИФР-I являются патогенетически значимыми факторами рака эндометрия, то изучение регуляции их содержания необходимо для четкого понимания механизмов развития заболевания и поиска эффективных противоопухолевых средств при молекулярно-направленной терапии. Полученные данные свидетельствуют о возможности использования ингибиторов протеасом в таргетной терапии рака эндометрия. Необходимы дальнейшие исследования протеасомной системы в контексте прогрессии и клинического течения рака эндометрия для выяснения ее возможной роли в метастазировании злокачественных опухолей и использовании для поиска новых маркеров прогноза течения заболевания.
Работа выполнена при финансовой поддержке Федеральной целевой программы (ФЦП) «Научные и научно-педагогические кадры инновационной России» на 2009 – 2013 годы (Гос. контракт № П-320).
