Птицы России и сопредельных стран: малая конюга Aethia pygmaea (Gmelin, 1789). 1. Внешний вид, распространение и численность
. 1. Внешний вид, распространение и численность")
Автор: Н.Б. Конюхов
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 2573 т.34, 2025 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140312752
IDR: 140312752
Birds of Russia and neighboring countries: the whiskered auklet Aethia pygmaea (Gmelin, 1789). 1. Appearance, distribution and abundance
Текст статьи Птицы России и сопредельных стран: малая конюга Aethia pygmaea (Gmelin, 1789). 1. Внешний вид, распространение и численность
Данная публикация представляет собой видовой очерк, аналогичный тем, что были опубликованы в сводке «Птицы СССР. Чистиковые». С тех пор прошло уже 35 лет, и наши знания значительно обогатились за счёт применения новых методов исследований; получены новые данные о распространении, экологии, поведению, генетике практически всех видов чистиковых. Планируется собрать всё известное на данный момент и опубликовать в серии статей, в которую войдут, по крайней мере, все виды чистиковых, отмеченные на территории России. Каждый видовой очерк будет состоять из трёх частей. Это – первая часть видового очерка, посвящённого малой конюге Aethia pygmaea (Gmelin, 1789).
Общая характеристика и полевые признаки
Мелкая чистиковая птица. Верхняя сторона тела чёрная с голубоватым серебристым оттенком, который виден только с близкого расстояния; серая окраска груди постепенно переходит грязно-белое брюшко и далее в чисто-белое подхвостье. На голове хохол из загибающихся вперёд узких пёрышек. Единственная из конюг, у которой на голове 3 пары белых косиц. Клюв красный с более светлой желтоватой вершиной (рис. 1).
На воде кажется чёрной, включая и клюв. В полёте хохол прижат к голове и не виден. Белое пятно в районе уздечки заметнее косиц, которые различимы лишь с небольшого расстояния и при хорошем освещении. Светлое брюшко заметно только в полёте и только когда птицы делают виражи (рис. 2.).
На воде малая конюга отличается от конюги-крошки Aethia pusilla тёмной грудью, от большой конюги Aethia cristatella – беловатым под-хвостьем и меньшими размерами, от белобрюшки Aethia psittacula – меньшими размерами и наличием хохолка.
Систематика
Анализ географической изменчивости малых конюг (Feinstein 1959) выявил различия в размерах и окраске птиц, населяющих Алеутские и
Курильские острова, и было высказано предположение, что алеутская и курильская популяции могут иметь статус подвидов: A. p. pygmaea и A. p. camtschatica , соответственно (рис. 3). Это разделение получило признание не всеми (Головкин 1990).

Рис. 1 (слева). Взрослая малая конюга Aethia pygmaea . Остров Булдырь. Фото Н.Конюхова.
Рис. 2 (справа). Малая конюга Aethia pygmaea в полёте. Пролив Малая Танага.
Фото Steven Mlodinow,

Рис. 3. Взрослые малые конюги в летнем оперении. А – Aethia pygmaea pygmaea , остров Булдырь, фото Н.Конюхова; Б – Aethia pygmaea camtschatica , остров Ионы, фото Ю.Артюхина
Aethia pygmaea pygmaea (Gmelin, 1789)
Распространение . Алеутские острова от острова Акун до Ближних и Командорских островов.
Описание внешнего вида . Такое описание алеутских птиц дано в оригинале статьи (Feinstein 1959). Голова и шея чёрные, переходящие в шиферный цвет на надхвостье, спине и верхних кроющих хвоста. Верхняя сторона рулевых перьев иссиня-чёрная. Бока головы (кроме уздечки) иссиня-чёрные; уздечка белая. Горло и верх груди иссиня-чёр-ные. В нижней части груди и на боках иссиня-чёрный цвет постепенно переходит в тёмный мышино-серый. Брюшная область бледного дымчато-серого цвета, переходящего в белый на нижних кроющих хвоста. Маховые перья темно-буро-чёрные. Клюв (сухие тушки) красный, с желтоватым кончиком. Лапы, перепонки и цевка (сухие тушки) в среднем коричневатые. Украшающие пучки перьев белые, за исключением чёрного лобного хохолка. Зимний наряд взрослых особей аналогичен описанному выше, но клюв коричневый.
Aethia pygmaea camtschatica (Lepechin, 1801)
Распространение . Курильские острова и острова Охотского моря.
Описание внешнего вида . Вот перевод описания подвида, которое сделал Бернард Фейнштейн (1959). Отличается от A. p. pygmaea более коричневатым (менее чёрным) оттенком и в целом крупнее по всем параметрам; крыло значительно длиннее; белая брюшная область резко контрастирует с коричневой грудью * ; голова и шея черновато-коричневые, переходящие в тёмный нейтрально-серый на спине, надхвостье и верхних кроющих хвоста; верхняя сторона рулевых перьев иссиня-чёр-ная; бока головы (кроме уздечки) черновато-коричневые; уздечка белая; горло и верх груди темно-буро-чёрные; нижняя часть груди и бока каштановые (hair brown); подхвостье и брюхо белые; маховые перья тёмнобурые; клюв (сухие тушки) красный с желтоватым кончиком; лапы, перепонки и цевка (сухие тушки) в среднем коричневатые; украшающие косицы белые, за исключением черноватого хохолка.
Замечания по систематике
Исследования О.С.Пшеничниковой (Пшеничникова 2017; Pshenichni-kova et al . 2017), посвящённые внутривидовой структуре малой конюги, выявили различия между популяциями вида, населяющими разные части ареала. У малой конюги дифференциация митохондриальных линий, вероятно, произошла в позднем плейстоцене. Дивергенции митохондриальных линий охотоморских и командорских конюг от алеутских птиц произошла около 126 тыс. лет назад. Образовались, видимо, два изолированных рефугиума. Охотоморские и командорские популяции разделились около 36 тыс. лет назад. Сейчас существует поток генов с Охотского моря на Командорские острова (1.57 самки за поколение; 95%
CI: 0.426-25.410) и в обратном направлении (1.28 самок за поколение; 95% CI: 0.455-26.110). Птицы из колоний Командорских и Алеутских островов с одинаковой суточной активностью на суше сильнее различаются между собой, нежели малые конюги с Охотоморья и Командор, у которых разная активность в колонии. Статистически значимой дифференциации между выборками малой конюги с разной суточной активностью в колонии выявлено не было (U’st = 0.039, Р >0.05) (Пшеничникова 2017).
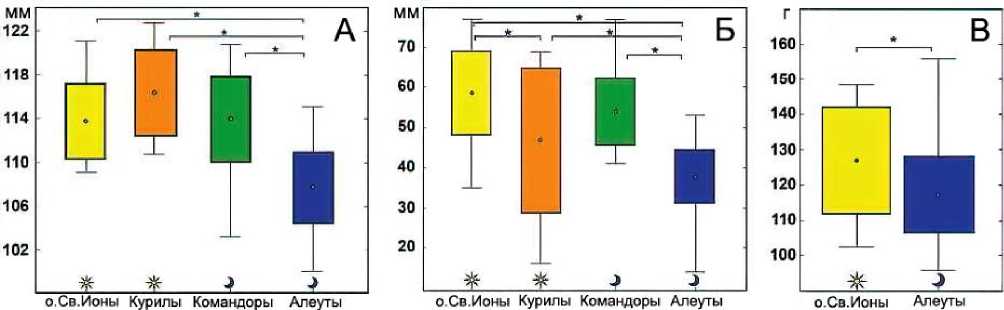
Рис. 4. Различия в длине крыла (А), длине хохла (Б) и массе (В) у малых конюг из разных колоний. Звёздочка (*) означает значимые различия (ANOVA, Tukey HSD post hoc test, P <0.05).
Точки – среднее значение; прямоугольник – SD , «усики» – lim. Символ солнца – колонии с дневной активностью птиц, луна – с ночной (Pshenichnikova et al . 2017)
Малые конюги с острова Ионы по сравнению с другими популяциями были самыми крупными. Их масса (141±12 г; SD , n = 20) не перекрывалась с массой птиц с Алеутских островов (111±7 г; SD , n = 110). У малых конюг алеутской популяции были значительно меньше крылья, хохол, цевка, высота и длина клюва, чем у других птиц (ANOVA, P <0.05 для всех переменных). Морфометрические показатели птиц с Курильских и Командорских островов занимали промежуточное положение между аналогичными параметрами конюг с острова Ионы и Алеутских островов, однако по ряду характеристик статистически не отличались от показателей птиц с острова Ионы (one-way ANOVA, P >0.05, рис. 4). Некоторые малые конюги с Командорских и Алеутских островов, у которых одинаковая суточная активность, значительно отличались друг от друга (ANOVA, тест Tukey HSD post hoc, P <0.05), в то время как некоторые птицы с острова Ионы и Командорских островов, имеющих разную суточную активность, не различались (ANOVA, Tukey HSD post hoc test, P >0.05) (Пшеничникова 2017; Pshenichnikova et al . 2017).
Результаты, полученные по генетическим маркерам и фенотипическим признакам, хорошо согласуются друг с другом. Малые конюги с островов Охотского моря значительно отличались от птиц с Алеутских островов как по фенотипическим, так и по генетическим маркерам. Полученные результаты противоречат гипотезе изоляции расстоянием, так как генетические дистанции не коррелируют с географическими. Ко-нюги с Командорских островов оказались генетически ближе к птицам Охотского моря, чем к птицам, гнездящимся на Алеутских островах, хотя расстояние между Командорскими и Алеутскими островами меньше (около 340 км) расстояния от Командорских островов до острова Ионы (около 1500 км) или до Курильских островов (около 1100 км). Результаты молекулярно-генетического анализа свидетельствуют о том, что в настоящее время поток генов между всеми группировками поддерживается, поэтому статус подвидов для данных групп весьма сомнителен (Пшеничникова 2017).
Результаты анализа фенотипических признаков свидетельствуют о клинальной изменчивости – малые конюги с острова Ионы и Алеутских островов чётко различаются между собой по морфометрическим и акустическим параметрам, а птицы с Курильских и Командорских островов занимают промежуточное положение (Пшеничникова 2017). Однако оказалось, что птицы с острова Эгг крупнее птиц с острова Булдыря по всем морфометрическим параметрам, включая украшающие образования: длина крыла больше на 3%, клюва – на 1%, цевки – на 1%, а также хохолка – на 7%, аурикулярной косицы – на 3%, риктальной косицы – на 3% и фронтальной косицы – на 3%. Птицы с острова Эгг также, по-ви-димому, крупнее птиц, измеренных на острове Улак в 2005 году (Jones, Seneviratne 2005) (табл. 1).
Таблица 1. Масса и размеры взрослых малых конюг (среднее ± SD*/SE**, в скобках n , затем lim)
|
Возраст |
Пол |
Масса, г |
Крыло, мм |
Цевка, мм |
Клюв, мм |
Год |
Источник |
|
|
Длина |
Высота |
|||||||
|
Алеутские острова |
||||||||
|
ad |
♂+♀ |
– |
103.76±3.0* (16) 99.4–108.1 |
21.14±0.91* (16) |
9.21±0.42* (16) |
– |
– |
Feinstein 1959 |
|
♂ |
– |
102.80±2.6* (9) 99.4–108.1 |
21.3 (9) 19.2–23.1 |
9.2 (9) 8.4–10.0 |
– |
– |
Feinstein 1959 |
|
|
♀ |
– |
101.90±3.6* (5) 97.3–106.0 |
20.7 (5) 20.1–22.0 |
9.3 (5) 9.0–9.7 |
– |
– |
Feinstein 1959 |
|
|
ad |
♂+♀ |
111±7* (110) |
107.7±3.2* (160) |
– |
9.4±0.5* (110) |
7.4±0.4* (110) |
– |
Пшеничникова 2017 |
|
Восточные Алеутские острова |
||||||||
|
ad |
♂+♀ |
116.7±10.8* (43) |
109.1±0.7* (48) |
21.6±1.1* (10) |
10.1±0.5* (10) |
– |
– |
Gaston, Jones 1998 |
|
Остров Егг |
||||||||
|
ad |
♂+♀ |
112.1±7.9* (54) 91–131 |
110.3±2.1* (54) 105–115 |
23.2±0.8* (54) 21.6–24.7 |
9.9±0.4* (54) 8.9–10.7 |
– |
2005 |
Jones, Seneviratne 2005 |
|
ad |
♂ |
117.35±9.12* (20) |
108.56±4.9* (16) |
21.84±1.45* (16) |
10.16±0.39* (14) |
7.35±0.22* (12) |
19921993 |
Pitocchelli et al . 2003 |
Продолжение таблицы 1
|
Возраст |
Пол |
Масса, г |
Крыло, мм |
Цевка, мм |
Клюв, мм |
Год |
Источник |
|
|
Длина |
Высота |
|||||||
|
ad |
♀ |
114.72±12.29* |
109.83±3.4* |
21.62±0.64* |
10.14±0.47* |
7.02±0.37* |
1992- |
Pitocchelli et al . |
|
(18) |
(18) |
(18) |
(18) |
(18) |
1993 |
2003 |
||
|
Западные алеутские острова |
||||||||
|
ad |
♂+♀ |
123.8±14.0* (11) |
109.2±2.6* (10) |
21.3±1.1* (9) |
10.0±0.5* (10) |
– |
– |
Gaston, Jones 1998 |
|
Остров Булдырь |
||||||||
|
ad |
♂+♀ |
110.8±0.44** |
106.5±0.19** |
20.9±0.05** |
9.3±0.03** |
7.2±0.02** |
1993 |
Конюхов и др. |
|
(155) |
(156) |
(110) |
(108) |
(108) |
2000 |
|||
|
109.±7.3* |
109.3±2.96* |
20.7±0.9* |
9.9±0.5* |
7.3±0.4* |
Н.Б.Конюхов, неопубл. данные |
|||
|
ad |
♂+♀ |
(70) 96–130 |
(62) 104–117 |
(62) 18.4–22.4 |
(62) 8.5–11.2 |
(62) 6.6–8.4 |
1999 |
|
|
106.2±6.2* |
106.6±3.0* |
20.8±0.8* |
9.9±0.6* |
7.01±0.3* |
Н.Б.Конюхов, неопубл. данные |
|||
|
sad |
♂+♀ |
(39) 90–117 |
(36) 100–111 |
(36) 18.9–22.4 |
(36) 8.4–10.8 |
(36) 6.6–7.9 |
1999 |
|
|
127.0±9.8* |
||||||||
|
ad |
♂ |
(27) 110–150 |
107.5±2.5* (28) |
21.0±0.8* (27) |
9.3±0.3* (28) |
– |
– |
Byrd, Williams 1993 |
|
118.1±7.0* |
||||||||
|
ad |
♀ |
(8) 105–126 |
105.6±3.3* (9) |
20.6±0.7* (9) |
8.9±0.4* (9) |
– |
– |
Byrd, Williams 1993 |
|
ad |
♂+♀ |
115.6±8.5* (94) |
– |
– |
– |
– |
– |
Byrd, Williams 1993 |
|
99–136 |
||||||||
|
ad |
♂+♀ |
111.9±7.3* (230) |
109.3±2.4* (10) |
21.8±0.7* (229) |
– |
– |
19941997 |
Hunter et a l. 2002 |
|
92–132 |
104.2–113.2 |
20.0–24.0 |
||||||
|
ad |
♂ |
– |
107.5±2.5* (28) |
21.0±0.8* (27) |
9.3±0.3* (28) |
– |
– |
Gaston, Jones 1998 |
|
ad |
♀ |
– |
105.6±3.3* (9) |
20.6±0.7* (9) |
8.9±0.4* (9) |
– |
– |
Gaston, Jones 1998 |
|
ad |
♂+♀ |
116.6±8.4* (228) 99–141 |
– |
21.9±0.8* (169) 20.3–24.2 |
9.5±0.5* (129) |
7.4±0.4* (168) 6.4–8.4 |
– |
Gaston, Jones 1998 |
|
sad |
♂+♀ |
107.0±6.3* (56) |
– |
– |
– |
– |
– |
Gaston, Jones 1998 |
|
94–120 |
||||||||
|
116±8.2* |
107.6±2.6* |
22.0±0.8* |
9.4±0.5* |
19922005 |
Jones, |
|||
|
ad |
♂+♀ |
(303) |
(45) |
(303) |
(262) |
– |
Seneviratne |
|
|
90–143 121±7* |
101–113 |
17.9–24.2 |
8.1–11.0 |
2005 Day, Byrd 1989 |
||||
|
ad |
♂+♀ |
(60) 102–138 |
– |
– |
– |
– |
1976 |
|
|
Командорские острова |
||||||||
|
ad |
♂ |
– |
111.1±0.7* (21) |
21.6±0.4* (17) |
9.5±0.1* (20) |
– |
– |
Byrd, Williams 1993 |
|
ad |
♀ |
– |
110.9±0.2* (10) |
21.5±0.2* (8) |
9.5±0.1* (9) |
– |
– |
Byrd, Williams 1993 |
|
108.3±3.2* |
22.0±0.5* |
9.9±0.3* |
||||||
|
ad |
♂+♀ |
– |
(18) |
(18) |
(17) |
– |
– |
Stejneger 1885 |
|
114–104 |
21–23 |
9–10 |
||||||
|
ad |
♂ |
– |
111.1±0.7* (21) |
21.6±0.4* (17) |
9.5±0.1* (20) |
– |
– |
Gaston, Jones 1998 |
|
ad |
♀ |
– |
110.9±0.2* (10) |
21.5±0.2* (8) |
9.5±0.1* (9) |
– |
– |
Gaston, Jones 1998 |
|
107.30±2.71* |
||||||||
|
ad |
♂+♀ |
– |
(31) 102.0–112.0 |
21.37±0.80* (31) |
9.28±0.48* (31) |
– |
– |
Feinstein 1959 |
Окончание таблицы 1
|
Возраст |
Пол |
Масса, г |
Крыло, мм |
Цевка, мм |
Клюв, мм |
Год |
Источник |
|
|
Длина |
Высота |
|||||||
|
111.60±1.4* |
23.05 |
9.4 |
||||||
|
♂ |
– |
(2) |
(2) |
(2) |
– |
– |
Feinstein 1959 |
|
|
110.2–113.0 107.07±3.1* |
23.0–23.1 |
9.1–9.7 |
||||||
|
♀ |
– |
(14) |
– |
– |
– |
– |
Feinstein 1959 |
|
|
102.0–112.0 |
||||||||
|
114.5 |
||||||||
|
ad |
♀ |
(2) 104–125 |
– |
– |
– |
– |
– |
Карташев 1979 |
|
107 |
||||||||
|
ad |
♂ |
(7) 93–115 |
– |
– |
– |
– |
– |
Карташев 1979 |
|
♂+♀ |
– |
113.8±3.9* (48) |
– |
9.8±0.4* (19) |
8.0±0.5* (17) |
– |
Пшеничникова 2017 |
|
|
Курильские острова |
||||||||
|
ad |
♂ |
– |
114.8±0.5* (24) |
22.6±0.2* (25) |
9.9±0.1* (25) |
– |
– |
Byrd, Williams 1993 |
|
ad |
♀ |
– |
110.6±1.3* (7) |
21.7±0.2* (9) |
9.7±0.2* (9) |
– |
– |
Byrd, Williams 1993 |
|
ad |
♂+♀ |
– |
111.98±2.16* (9) |
22.15±0.53* (9) |
9.60±0.33* (9) |
– |
– |
Feinstein 1959 |
|
ad |
♂ |
– |
114.8±0.5* (24) |
22.6±0.2* (25) |
9.9±0.1* (25) |
– |
– |
Gaston, Jones 1998 |
|
ad |
♀ |
– |
110.6±1.3* (7) |
21.7±0.2* (9) |
9.7±0.2* (9) |
– |
– |
Gaston, Jones 1998 |
|
112.43±2.7* |
21.7 |
9.8 |
||||||
|
♀ |
– |
(3) |
(3) |
(3) |
– |
– |
Feinstein 1959 |
|
|
110.0–115.4 |
21.3–22.0 |
9.5–10.2 |
||||||
|
♂+♀ |
116.3±3.9* |
9.8±0.8* |
7.7±0.6* |
Пшеничникова |
||||
|
– |
(12) |
– |
(12) |
(12) |
– |
2017 |
||
|
Остров Ионы |
||||||||
|
ad |
♂+♀ |
141±12* |
113.7±3.4* |
9.8±0.4* |
8.0±0.3* |
Пшеничникова |
||
|
(20) |
(20) |
– |
(20) |
(5) |
– |
2017 |
||
|
145.1±6.4** |
Андреев и др. 2012 |
|||||||
|
ad |
♂+♀ |
(13) 133–155 |
– |
– |
– |
– |
2009 |
|
Статистических различий как морфометрических промеров, так и размеров скелетных элементов между самцами и самками в разных возрастных группах на острове Егг обнаружено не было (Pitocchelli et al . 2003).
Считалось, что у малых конюг существует клинальное увеличение размеров тела с востока на запад, причём самые мелкие птицы населяют восточные Алеуты, а самые крупные – острова азиатской части ареала (Feinstein 1959; Byrd, Williams 1993; Pitocchelli et al. 2003; Пшеничникова 2017). Однако результаты исследований на острове Егг противоречат этому (Jones, Seneviratne 2005). Вполне вероятно, что морфологические различия могут быть результатом ограниченного осёдло-стью особей потока генов (Schacter, Jones 2018), и у малых конюг на разных островах или группах близкорасположенных островов существуют отдельные дифференцированные популяции, а не одна свободно смешивающаяся популяция Алеутских островов (Jones, Seneviratne 2005). И для выяснения как морфометрических, так и генетических различий между локальными популяциями необходимы данные из большего числа колоний по всему ареалу вида.
Внешний вид
Малые конюги мономорфны практически по всем промерам, но высота клюва и ширина черепа могут быть использованы для определения пола птиц. Самцы несколько крупнее самок (табл. 1). Полового диморфизма в окраске нет (Pitocchelli et al . 2003). Цветовых морф не отмечено.
Взрослый летний (брачный) наряд (Definitive Alternate)
Оперение . Верх и бока головы, а также вся верхняя сторона тела аспидно-чёрные с серебристо-голубым оттенком, который более выражен на спине, пояснице и надхвостье. Подбородок, горло и шея спереди темно-серые, область зоба, верхняя часть груди и бока тела серые, светлее, чем шея Середина груди, брюшко и подхвостье беловатые, грязно -белые или белые. В начале сезона размножения рулевые, маховые и верхние кроющие крыльев чёрные с маслянистым блеском, к концу гнездового сезона, по мере снашивания, перья утрачивают блеск и становятся черно-бурыми, а иногда и коричневыми. Крыло снизу тёмное и разделено по цвету на две части: серебристо-серые маховые и их большие кроющие и тёмные остальные кроющие и перья пропатагиума. Первостепенные и второстепенные маховые снизу серебристо-серые; внешнее опахало менее серебристое, более бурое. Большие кроющие первостепенных и второстепенных маховых целиком серебристо-серые. Средние кроющие первого-четвёртого первостепенного махового — серебристо-серые с бурыми вершинами, но граница между цветами размыта; пятого-десятого — буровато-темно-серые. Средние кроющие второстепенных маховых серебристо-серые с узкой достаточно чёткой светло-бурой вершинной каёмкой. Малые кроющие первостепенных, как и средние кроющие, серебристо-серые, но с широкой тёмной серо-бурой каймой, образующей заметный струйчатый рисунок. Малые кроющие второстепенных маховых и перья пропатагиума — коричнево-буроватые с более тёмной каймой, образующей заметный струйчатый рисунок. Первостепенных маховых 11, одиннадцатое рудиментарно, а вершину крыла образует десятое первостепенное. Рулевых 14.
Украшающие образования представлены хохолком или султаном и тремя парами косиц — удлинённых нитевидных перьев (рис. 5). Хохолок состоит из 10-12 загибающихся вперёд полукольцом чёрных перьев. По структуре это обычные контурные перья. Перья косиц представляют собой filoplumulae (Конюхов 2001). Фронтальная и риктальная косицы начинаются от общего белого пятна, расположенного в районе уздечки и отделённого от надклювья узкой полоской чёрных перьев. Фронтальные косицы проходят над глазом, загибаясь кверху, к затылку, и выступают над теменем в виде двух небольших хохолков. Риктальная косица начинается на уровне угла клюва и проходит под глазом через щёку. Аурикулярная косица начинается от глаза и спускается книзу на всю длину шеи. Данное выше описание относится к наибольшей степени их развития у номинативного подвида; обычно косицы короче. У подвида A. p. camtschatica украшающие образования длиннее.
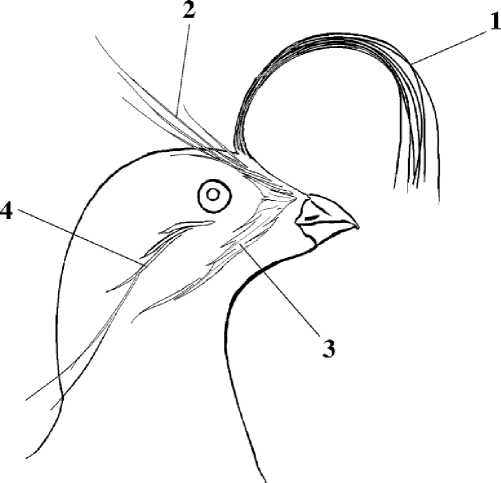
Рис. 5. Украшающие образования малой конюги. 1 – султан (хохолок); косицы: 2 – фронтальная, 3 – риктальная, 4 – аурикулярная (Конюхов 2001)
Размеры украшающих образований представлены в таблице 2.
К filoplumulae относятся и нитевидные перья, которые растут по всему телу птиц, но наиболее многочисленны на шее и голове. Собственно нитевидные перья самые короткие. Они не выступают над поверхностью контурного оперения и, вероятно, являются, скорее всего, сенсорами давления при нырянии. Опахала нитевидных украшающих перьев более развиты, а сами перья длиннее и выступают над поверхностью оперения. Нитевидные украшающие перья есть только на голове и шее. Фактически нитевидные, украшающие перья и перья косиц отличаются друг от друга лишь длиной ствола и степенью развития на нём опахала. Перья, формирующие косицы – самые длинные. Их опахала хорошо развиты. Различия между этими тремя перьевыми структурами условны, и у любой птицы можно проследить все переходы от крошечных, всего лишь в несколько миллиметров, нитевидных перьев к перьям косиц, достигающих длины десятков миллиметров. Идентичность этих перьевых структур подтверждается как их положением относительно контурных перьев, так и сходством строения (Конюхов 2001). У контурных перьев чистиковых, как и у остальных ржанкообразных, есть побочный стержень (Козлова 1957, 1961). Однако у filoplumulae его нет. Отсутствие побочного стержня у перьев косиц не может быть вызвано незначительным их размерами, поскольку у малой конюги они по длине в несколько раз превосходят мелкие контурные перья, а диаметр очина у тех и других одинаков. Однако у маленьких контурных перьев есть хотя бы незначительный как по длине, так и по толщине побочный стержень, а у украшающих перьев он отсутствует. Кроме того, как и нитевидные перья, они всегда ассоциированы с контурными перьями. Это показывает, что перья косиц – это filoplumulae, а не контурные перья (Конюхов 2001).
Таблица 2. Размеры украшающих образований (мм) у малой конюги (среднее ± SD *, в скобках n , затем lim)
|
Возраст |
Пол |
Хохолок |
Косицы |
Год |
Источник |
||
|
Фронтальная |
Аурикулярная |
Риктальная |
|||||
|
Остров Егг |
|||||||
|
ad |
♂+♀ |
43.9±4.6* (54) |
35.0±6.8* (54) |
35.4±3.8* (54) |
33.2±4.0* (54) |
2005 |
Jones, Seneviratne |
|
30.9–52.1 |
15.9–49.3 |
23.8–43.8 |
25.6–42.0 |
2005 |
|||
|
Остров Булдырь |
|||||||
|
ad |
♂+♀ |
37.3±6.1* (299) |
31.8±6.2* (252) |
32.7±4.0* (253) |
30.3±3.2* (252) |
1992- |
Jones, Seneviratne |
|
13.6–57.4 |
9.6–46.6 |
19.1–44.0 |
18.3–39.3 |
2005 |
2005 |
||
|
ad |
♂+♀ |
36.8±6.7* (225) 8.5–57.0 |
29.2±8.5* (158) 0–46.6 |
32.6±4.1* (124) 20.5–43.9 |
29.8±3.6* (123) 16.1–37.8 |
– |
Gaston, Jones 1998 |
|
sad |
♂+♀ |
19.8±8.8* (55) |
22.0±7.6* (32) |
28.2±3.1* (32) |
26.9±3.2* (32) |
Gaston, Jones |
|
|
0–38.0 |
0–38.0 |
21.6–34.8 |
19.1–32.9 |
1998 |
|||
|
ad |
♂+♀ |
38.72±7.32* (33) |
32.75±5.85* (33) |
– |
– |
2006 |
Seneviratne, Jones 2008 |
|
ad |
♂+♀ |
33.8±6.5* (94) |
30.3±6.7* (87) |
31.9±4.0* (87) |
29.0±3.7* (88) |
Byrd, Williams 1993 |
|
|
8.5–44.4 |
9.6–46.6 |
20.5–43.1 |
16.1–37.8 |
||||
|
Алеутские острова |
|||||||
|
ad |
♂+♀ |
37.8±6.5* (96) |
– |
– |
– |
– |
Пшеничникова 2017 |
|
ad |
♂ |
39.3 (9) 34.6–47.2 |
– |
– |
– |
– |
Feinstein 1959 |
|
ad |
♀ |
33.9 (5) 28.2–40.0 |
– |
– |
– |
– |
Feinstein 1959 |
|
Командорские острова |
|||||||
|
ad |
♂+♀ |
53.9±8.3* (19) |
– |
– |
– |
– |
Пшеничникова 2017 |
|
ad |
♂ |
49.1 (2) 46.2–52.0 |
– |
– |
– |
– |
Feinstein 1959 |
|
ad |
♀ |
49.8 (3) 48.2–53.1 |
– |
– |
– |
– |
Feinstein 1959 |
|
Курильские острова |
|||||||
|
ad |
♂+♀ |
46.9±18.0* (11) |
– |
– |
– |
– |
Пшеничникова 2017 |
|
Остров Ионы |
|||||||
|
ad |
♂+♀ |
58.6±10.5* (15) |
– |
– |
– |
– |
Пшеничникова 2017 |
Исследования, направленные на выявление асимметрии косиц, обнаружили отсутствие свидетельств того, что у неполовозрелых птиц они более асимметричны, чем у взрослых птиц; не было найдено связи между асимметрией украшающих перьев и возрастом (некоторые птицы отлавливались в течение нескольких лет), не найдено корреляции между асимметрией и физическим состоянием птицы, а также не было доказательств того, что у неразмножающихся птиц украшающие образования были более асимметричными, чем у размножающиеся (Jones et al. 2023). В то же время длина хохолка и фронтальной косицы положительно коррелируют как между собой, так и с размером птиц (Seneviratne, Jones 2008).
Украшающие образования на головах малых конюг, кроме визуальных маркеров физического состояния птиц, имеют ещё утилитарную функцию. Экспериментальные исследования позволяют предположить, что хохолок и фронтальные косицы малые конюги используют как сенсорные структуры, позволяющие птицам лучше ориентироваться «на ощупь» и не натыкаться на препятствия, когда они ночью пробираются в свои гнездовые камеры по длинным, извилистым и узким трещинам в скалах. В эксперименте, в лабиринте, испытуемые птицы, хохолок и фронтальные косицы которых были приклеены к оперению липкой лентой, на 275% чаще касались головой неровностей лабиринта, чем птицы контрольной группы без клейкой ленты на оперении или птицы, у которых клейкая лента была приклеена к оперению, но хохолок и косицы оставались свободными (Seneviratne, Jones 2008, 2010).
Малые конюги, как и большие, обладают характерным цитрусовым запахом (Jones 1993). Установлено, что этот специфический аромат обусловлен наличием альдегидов с чётным числом углеродных атомов в цепях (Douglas et al . 2001). У птиц обнаружены небольшие уникальные специализированные «фитильные» перья, характеризующиеся выраженной пористой волосовидной структурой. Каждое такое перо развивается из специализированного шаровидного основания, содержащего, вероятно, секреторные клетки, продуцирующие пахучие соединения (одорант) (Douglas 2011). Предполагается, что этот одорант отпугивает эктопаразитов от взрослых особей и «окуривает» место гнездования. Существует гипотеза, согласно которой продукция специфического цитрусового запаха и развитие украшающих структур у малых конюг представляют собой взаимосвязанные вторичные половые признаки. Важно отметить, что запах выполняет двойную функцию: является индикатором качества потенциального партнёра и обеспечивает устойчивость к эктопаразитам, что является предполагаемой основой для выбора партнёра у малых конюг (Douglas et al . 2001).
Неоперённые части тела. Клюв короткий, выпуклый по коньку надклювья. Ноздри узкие, щелевидные. Рамфотека состоит из 4 парных и 2 непарных щитков (рис. 6) (Конюхов 2001), но границы между ними становятся заметными только во время смены рамфотеки. Клюв в основ- ном ярко-красный. Вершина надклювья светлая, беловатая или светло -серая. Кульменальный, гониальный, нижний томиальный и субнариаль-ные щитки — темно-красные, оперкулярный — бледно-красный, верхний томиальный — светло-серый.
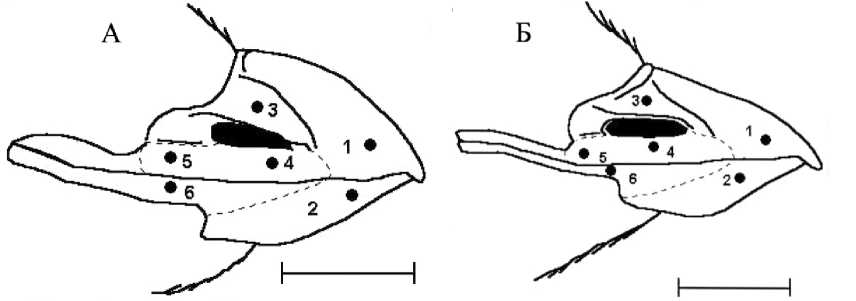
Рис. 6. Строение рамфотеки малой конюги: А – взрослая птица, Б – слёток. Щитки: 1 – кульменальный, 2 – гониальный, 3 – оперкулярный, 4 – субнариальный, 5 – верхний томиальный, 6 – нижний томиальный. Пунктиром показаны примерные границы между щитками (масштабный отрезок равен 5 мм) (Конюхов 1994, 2001)
Зев и язык очень светлые, бледно-розовые или почти белые с незначительным светло-телесным оттенком.
Орбитальное кольцо не выражено. Радужина чисто-белая либо белая с голубоватым оттенком, как правило, очень слабым.
Цевка и пальцы сверху светлые, голубовато-серые или бледно-голубые; на суставах более тёмные за счёт большего количества прикрывающих их щитков, кожа между которыми почти чёрная, нижняя сторона пальцев и цевки — серовато-чёрные, черноватые или чёрные, но цвет не насыщенный, тусклый. Перепонки сверху и снизу чёрные, но на верхней стороне в большей или меньшей степени может быть выражен голубой оттенок. Когти чёрные.
Взрослый зимний наряд (Definitive Basic) *
Оперение . По окончании полной линьки, когда украшающие образования достигают нормальных размеров, зимний наряд взрослых птиц, видимо, ничем не отличается от летнего, за исключением того, что он более яркий, так как перья ещё не обношены. Не упоминая источник, А.Н.Головкин (1990) пишет, что в зимнем наряде окраска горла птиц становится беловатой, а зоба и груди — серой. Поскольку Л.Штейнегер (Stejneger 1885) отмечает, что у птиц, добытых весной, идёт линька контурного оперения, то можно предположить, что зимняя окраска горла, зоба и груди может отличаться от летней.
Украшающие перья малых конюг во взрослом зимнем оперении короче (Byrd, Williams 1993).
Неоперённые части . Структура рамфотеки сходна с таковой в летнее время. Зимой клюв не такой яркий, как летом. Наиболее верное описание, видимо, дано Л.Штейнегером (Stejneger 1885), описывавшего све-жедобытых птиц у Командорских островов зимой. Клюв молочно-белый на вершине и у основания подклювья. Остальная поверхность клюва, включая кульменальный, верхний томиальный щитки и среднюю часть гониального щитка – ярко-красные, оперкулярный щиток и часть куль-менального щитка, примыкающая к ноздре – черновато-коричневые.
Окраска зева – как и летом; Л.Штейнегер (Stejneger 1885) его описывал как очень светлый, почти белый с незначительным светло-телесным оттенком. Орбитальное кольцо не выражено. Окраска радужины – как и летом (Stejneger 1885). Цевка, пальцы, перепонки и когти окрашены так же, как и летом (Stejneger 1885).
Пуховой птенец (Downy) (+ к описанию взрослых)
Пуховой наряд малых конюг представлен эмбриональным, постэмбриональным и мезоптильным пухом (рис. 7). Как ни кажется это странным, но пуховой наряд малых конюг, как и всех чистиковых, гнездящихся в укрытиях, камуфляжного типа. Длинный пух чёрно-серых тонов с постепенными переходами окраски полностью размывает очертание птенца в тёмной гнездовой камере, так что его порой трудно разглядеть даже при свете фонаря.

Рис. 7 (слева). Пуховой птенец малой конюги Aethia pygmaea . Остров Булдырь. Фото Н.Конюхова.
Рис. 8 (справа). Слёток малой конюги Aethia pygmaea . Остров Булдырь. Фото Н.Конюхова
Эмбриональный наряд (neossoptile). В эмбриональном наряде птенец покрыт густым типичным эмбриональным пухом neoptile plu-mulae. На спинной стороне пух чёрный; на голове он кажется чернее, чем на спине, однако его оттенки зависят от направления падающего света. Грудь серовато-чёрная, брюхо серое. На кисти пух серый, как на брюхе, контрастирующий с чёрным пухом на боках. Длина пуха на вен- тральных птерилиях составляет 14-19 мм, на дорсальных – 20-22 мм. Наиболее короткий пух растёт вокруг клюва, и чем ближе к клюву, тем он короче и светлее. По характеру вытеснения эмбриональный пух является предшественником контурного пера praepennae I типа (Конюхов и др. 2000).
Неоперённые части . При вылуплении аптерии голые, хорошо выраженные; цвет кожи, который можно на них видеть – красновато-телесный. Клюв: границы между щитками не выражены. Надклювье чернороговое, более тёмное на коньке и по режущему краю, кончик светлее, чем остальной клюв. Оперкулярный щиток серовато-роговый с чёрным вентральным краем. Подклювье телесного цвета – беловато-телесное с чёрной вершиной. Верхний томиальный щиток черноватый (светлее клюва). Яйцевой зуб белый, 1-2 мм. Полость рта светло-розовая. Язык чуть светлее полости рта, его концевая (ростральная) часть черноватая, причём, чем ближе к краю, тем интенсивнее чёрный цвет.
Орбитальное кольцо не выражено. Радужина при вылуплении темно-серая, очень тёмная (Конюхов и др. 2000).
Цевка и пальцы сверху серовато-телесные с синеватым оттенком. Цевка снизу черноватая, пальцы, как и сверху, но темнее. Перепонки телесного цвета как сверху, так и снизу. Когти на лапе темно-серые или роговые. На крыле коготь есть только на втором пальце. Он, как правило, светлый – почти белый, светло-серый или серый с черноватым кончиком.
Цветовые морфы не отмечены.
Оперение при слёте . Слётки покидают гнездовую камеру в полностью сформированном оперении, которое представлено контурным пером I типа и пуховидным пером I и II типа (рис. 8). Количество контурных перьев на переднем крае крыла заметно больше, чем суммарное количество эмбриональных и постэмбриональных пушин. Это указывает на развитие части контурных перьев из нефункционирующих сосочков, то есть они относятся к контурному перу II типа (Конюхов и др. 2000).
Верх головы, шея сзади, спина, поясница, крылья и надхвостье – чёрные с более или менее выраженным серебристо-голубоватым оттенком, который не так интенсивен, как у взрослых птиц. Маховые и рулевые – чёрные. Подбородок и горло – светло-серые или бежевые. За счёт белых вершин перьев может присутствовать незначительный белый струйчатый рисунок. Шея с боков темно-серая. Шея спереди и верх груди – серые. Серый цвет на груди постепенно переходит в белый за счёт увеличения белого сектора в вершинной части пера; нижняя часть груди, брюхо и подхвостье – белые, причём чем ближе к подхвостью, тем оперение, как правило, становится белее и часто подхвостье бывает чисто-белым. Бока серые, иногда светлее груди; здесь белые каёмки перьев создают лёгкий струйчатый рисунок.
Крыло снизу в основном серое. Все маховые и их большие кроющие серебристо-серые. Остальные нижние кроющие — серые с более светлой вершинной каёмкой, создающей неоднородность окраски. Внутренние первостепенные маховые уже полностью выросли, хотя внешние ещё продолжают расти.
Хохолка у слётков нет. Косицы есть, но степень их развития у разных птиц неодинакова. Аурикулярная косица представлена только пятном под глазом, окрашенным в бежевый цвет разной интенсивности. Задняя граница этого пятна размыта. Аурикулярная и риктальная косицы окрашены в светло-серый или грязно-белый цвет с желтоватым оттенком. Последняя из них обычно выражена в меньшей степени, а порой и отсутствует вовсе. Обе эти косицы состоят из перьев, сходных по структуре с filoplumulae (Конюхов и др. 2000). У слётков могут сохраняться редкие пушинки на крестце и шее (Карташев 1979; Н.Б.Конюхов, неопубл. данные.). Молодые особи осенью, после слёта, не линяют и сохраняют своё оперение до следующего лета.
Неоперённые части . Клюв: рамфотека без видимых границ между щитками. Деление рамфотеки на щитки такое же, как у взрослых птиц (рис. 3). Надклювье как правило чёрное, но у небольшого числа слётков некоторые участки кульменального и субнариального щитков могут быть черновато-телесного цвета. Подклювье обычно светлее надклювья; в основании гониального щитка имеется более светлое пятно, размер которого у разных особей варьирует (Конюхов и др. 2000). Яйцевой зуб отпадает в возрасте 2-3 недель (Конюхов и др. 2000). Полость рта и язык бледно-розовые.
Орбитальное кольцо не выражено. Радужина у птенцов по мере их роста постепенно светлеет, проходя через разные оттенки серого цвета. К слёту она становится голубовато-серой или голубовато-белой (Конюхов и др. 2000).
Цевка и пальцы сверху светлые, голубовато-серые или серовато-голубые с более темными суставами за счёт чёрной кожи между щитками; снизу — чёрные. Перепонки сверху и снизу чёрные. Когти чёрные.
Первый зимний наряд (Basic I)
Оперение. Этот наряд, по-видимому, формируется в результате частичного обнашивания птенцового оперения и практически не отличается от него, за исключением более развитых косиц и хохолка. К концу августа (то есть примерно через месяц после слёта) среди сеголеток уже встречаются особи, султан и косицы которых начинают расти (рис. 9). У молодой птицы, пойманной 26 августа 1999 на острове Булдырь, косицы были хорошо развиты, достигая практически таких же размеров, как у неполовозрелых птиц, пойманных в это же время. Таким образом, молодые птицы всё больше начинают походить на неполовозрелых малых конюг, встречающихся в колонии в течение всего сезона размножения и считавшихся двухлетками, но на самом деле являющимися годовалыми особями (Конюхов, Зубакин 1994; Конюхов и др. 2000). Слётки, добытые на Командорах 18 и 22 июля 1882, были похожи на взрослых, только их косицы состояли их небольших сероватых перьев. В таком наряде молодые остаются по крайней мере до начала января (Stejneger 1885).

Рис. 9. Молодая малая конюга Aethia pygmaea примерно через месяц после слёта. Остров Булдырь. Фото Н.Конюхова
Неоперённые части . Клюв: структура рамфотеки сходна с таковой у взрослых птиц. Тенденция, отмеченная в течение первого месяца жизни молодых птиц, говорит о том, что высота их клюва после слёта продолжает увеличиваться. У молодых птиц примерно через месяц после слёта она была статистически значимо больше, чем у слётков ( t = 2.44; P = 0.0166): соответственно 6.6±0.3 мм ( SD , n = 71) и 6.4±0.2 мм ( SD , n = 28) (Конюхов и др. 2000; Н.Б.Конюхов, неопубл. данные). Окраска клюва, видимо, сходна с таковой у взрослых птиц зимой. В пользу этого свидетельствует тот факт, что к началу сентября у некоторых молодых птиц медиальные участки кульминального и гониального щитков рамфотеки начали менять окраску с роговой на красноватую. (Конюхов, Зубакин 1994; Конюхов и др. 2000). На Командорах у птицы, добытой 22 августа 1882, клюв был красновато-чёрный; в начале января он становится ярче (Stejneger 1885).
Зев и язык: данных нет, но поскольку цвет полости клюва и языка у слётков и взрослых птиц одинаков, то и у птиц промежуточных возрастов он, видимо, будет таким же, то есть бледно-розовым.
Орбитальное кольцо не выражено. О цвете радужины точных данных нет. Судя по тенденции изменения цвета глаз у малой конюги с возрастом, радужина должна иметь промежуточную окраску между таковой у слётков и первогодков, то есть доля птиц с голубоватым или жемчужно-серым оттенком радужины постепенно уменьшается, а доля с белым – постепенно возрастает.
Цевка, пальцы, перепонки и когти: данных нет, но надо полагать, что их цвет сходен с таковыми у взрослых птиц.
Первый летний наряд (Alternate I)
Оперение . Этот наряд сходен с брачным, отличаясь отсутствием серебристо-голубого оттенка на спине, пояснице и надхвостье и меньшим развитием украшающих образований. В отличие от взрослых, оперение крыльев более обношено, постольку оно на несколько месяцев «старше», чем у взрослых птиц.
Летние отловы птиц паутинными сетями на острове Булдырь (Конюхов и др. 2000) показали, что в колонии присутствуют только взрослые (ad) и неполовозрелые (sad) птицы. У последних отсутствовали насед-ные пятна, а украшающие образования (султанчик и косицы) были развиты в меньшей степени, чем у первых. Птиц, имеющих какую-либо другую внешность, отличную от перечисленных выше, отмечено не было. Логично предположить, что если молодые птицы текущего сезона размножения возвращаются в колонию, то и более старшие птицы, на следующую весну после их слёта, тоже посещают колонию. Следовательно, птицы, которые ранее принимались за неполовозрелых, являются лишь годовалыми птицами, то есть малые конюги весной сразу одевают наряд, аналогичный второму летнему других конюг, «пропуская» первый зимний (Конюхов, Зубакин 1994).
Это предположение подтвердилось. Из 154 молодых птиц (слётков), окольцованных в 1993 году на острове Булдырь, на следующий год было поймано 8 особей, 4 из которых по степени развития украшающих перьев были определены И.Джонсом (I.Jones, устн. сооб.) как неполовозрелые, а остальные – как взрослые особи (Зубакин, Конюхов 2001). Многолетние отловы малых конюг на одном и том же месте выявили, что слётки, пойманные на следующий год, выглядели как неполовозрелые птицы, а все из 54 пойманных и окольцованных неполовозрелыми птиц на следующий год были во взрослом оперении. Следовательно, малые конюги приобретают оперение неполовозрелых птиц на следующий год после слёта, то есть неполовозрелые птицы – это птицы, которым исполнился только один год (Jones et al . 2007). Всё сказанное выше, однако, не позволяет утверждать, что половозрелость малых конюг наступает в двухлетнем возрасте, а не в трёхлетнем, как у остальных видов конюг рода Aethia (Конюхов и др. 2000).
Неоперённые части тела – см. «Взрослый летний (брачный) наряд (Definitive Alternate)».
Второй зимний наряд (Basic II)
Оперение и неоперённые части . Точных данных нет, но, вероятно, сходны с таковыми у взрослых птиц.
Цветовые аномалии . Данных нет.
Распространение, численность и их динамика
Область гнездования . Малая конюга гнездится на Алеутских, Командорских, Курильских островах и внутренних островах Охотского моря (рис. 10). Вследствие кормовой специализации встречается в непосредственной близости от колоний. Наблюдения, проведённые в гнездовой сезон, показали, что малые конюги в границах гнездовой части ареала на Алеутских островах встречались в непосредственной близости от берега (до 10-12 км) – там, где существуют возмущения водной массы, вызванное приливными течениями (Н.Б.Конюхов, неопубл. данные). Аналогичные данные на Курильских островах были получены А.И.Гизенко (1955), который наблюдал крупные стаи малых конюг летом в открытом море на расстоянии 5-10 миль от берега. К сожалению, точного места этих наблюдений автор не приводит. В пределах этой обширной области, вероятно, известны далеко не все места гнездования малой конюги. Большинство островов редко посещаются в период размножения вида, и случайное обследование потенциальной колонии в течение дня не выявляет присутствия малых конюг из-за их скрытного гнездования в щелях и ночной активности в восточной части ареала. Следовательно, малые конюги, возможно, размножаются в большем числе мест, чем известно до сих пор.
На Алеутских островах малые конюги гнездятся на 37% островов, где имеются колонии морских птиц; всего зарегистрировано 48 их колоний. Наибольшая численность этого вида отмечена на острове Булдырь, а также на островах, расположенных между островами Адак и Большой Ситкин, на острове Сигуам, на Четырёхсопочных и Бабьих островах. Примечательно, что за исключением Булдыря, в основных районах скопления малых конюг в большинстве случаев отсутствуют конюги-крошки и большие конюги (Byrd et al . 2005).
Несмотря на то, что рассматриваемый вид зарегистрирован на всей Алеутской цепи, его распространение там было явно сгруппированным. Это хорошо было видно перед началом сезона размножения; 97% малых конюг на Лисьих островах встречены между западной оконечностью острова Тигальда и западной частью Бабьего пролива, 98% птиц на Четырёхсопочных островах отмечены между островами Херберт и Юнаска, а 95% всех птиц на Андреяновских островах обнаружены в двух местах: 30% близ Сигуама и 65% вблизи острова Большой Ситкин. На долю этих трёх групп приходилось в сумме 85% малых конюг, которые были встречены на Алеутских островах в апреле 1974 года (Byrd, Gibson 1980).
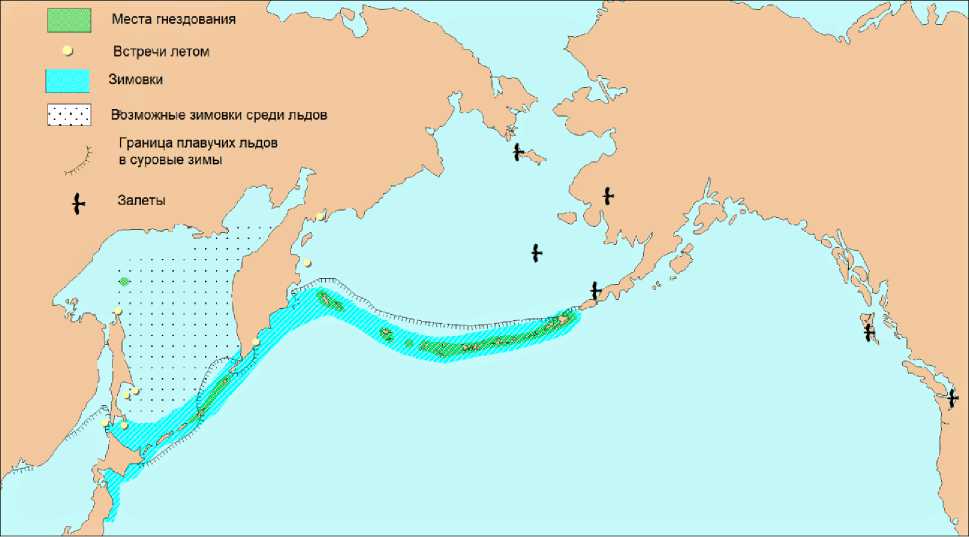
Рис. 10. Распространение малой конюги
Стаи из тысяч или даже десятков тысяч малых конюг часто встречаются в проливах Уналга и Акутан. Если предположить, что это местные птицы, то большинство из них должны гнездиться на Бабьих островах, что маловероятно, учитывая предыдущие оценки численности (Nyse-wander et al . 1982; Bradstreet, Herter 1991), или же на близлежащих островах Уналашка, Уналга и Акутан, хотя на них обитают аборигенные популяции лисиц Vulpes vulpes , а на Уналашке ещё есть и серая крыса Rattus norvegicus . Берега этих островов обрываются в море значительными скальными стенами с многочисленными расселинами и с участками, покрытыми лишайником рода Caloplaca оранжевого цвета, который может указывать на присутствие морских птиц; скальные стенки на острове Эгг, где гнездятся малые конюги, покрыты именно этим лишайником (Jones, Seneviratne 2005).
В азиатской части ареала сведения о гнездовании малой конюги порой противоречивы. А.Г.Велижанин (1978), ссылаясь на Л.А.Портенко (1934, 1937, 1973), пишет, что малая конюга встречается на острове Рат-манова, однако в цитированных публикациях о регистрации малой ко-нюги на этом острове ничего не сказано. При последующих обследованиях острова в 1987 и 1991 годах малая конюга там обнаружена не была (Зубакин и др. 1992; Konyukhov et al . 1998).
Малая конюга на Командорских островах – наиболее обычна среди мелких чистиковых птиц (Артюхин 1991). Она, вероятно, всегда была редка на острове Беринга. Леонард Штейнегер (Stejneger 1885) писал, что местные жители не так хорошо, как жители острова Медный, были знакомы с этим видом, и поэтому у них не было особого названия для него; иногда этих птиц называли «малыми конюгами». В то же время жителям острова Медный этот вид был хорошо известен под алеутским названием «турутурк».
В настоящее время малую конюгу можно встретить на гнездовании по всему побережью Медного, где есть подходящие расщелины по его крутыми скалистыми берегами. На острове Беринга малая конюга довольно редка (Stejneger 1885; Иогансен 1934), гнездится также в расщелинах острова Арий Камень (Stejneger 1885). В.Л.Бианки (1909), видимо, вторя более ранней публикации (Stejneger 1885), пишет, что малая конюга «обыкновеннее на Медном, чем на острове Беринга, где держится между прочим на Арьем Камне, у Саранной рыбалки и на Диком мысу». На Командорах чаще встречается зимой, чем летом (Иогансен 1934). На воде малые конюги отмечались как в акватории Медного, так и акватории острова Беринга (Артюхин 1999), но гнездятся в основном на острове Медный (Артюхин 2016а). Во время учётов колониальных морских птиц на острове Беринга в 2015-2022 годах эти конюги в гнездовой период неоднократно встречались в акватории острова на расстоянии до 0.5 км от берега (Пилипенко 2025). Ночью в начале июля 1988 года на острове Топорков отмечена вокализация одиночных птиц, но на острове Арий Камень этот вид не найден (Артюхин 1990).
Курильские острова, в особенности их центральная часть, представляют собой ключевой район гнездования данного вида в азиатском секторе его ареала. В 1896 году малые конюги встречены на скалах Мушир (острова Ловушки) 22 августа, на Райкоке 23 августа, на Северном Уши-шире (острове Рыпонкича) 24 августа, на Южном Ушишире (острове Ян-кича) 25 августа (Stejneger 1899).
Северная граница ареала вида на Курилах – остров Парамушир, где эти птицы в незначительном числе гнездятся в осыпях и трещинах скал на склонах вулкана Фусса (Велижанин 1971 – по: Артюхин и др. 2001). Единичные малые конюги, встреченные в северной части пролива Лужина 7 июля 2000 (Артюхин и др. 2001), могли гнездиться как на склоне вулкана Фусса, так и на острове Анциферова. Ёсимаро Ямасина (Yamashina 1931) считал, что на Парамушире вид вообще не гнездится.
На Средних Курилах малая конюга гнездится на большинстве островов и крупных скалах. Колонии находятся на следующих островах: Экарма, Чиринкотан, Шиашкотан, острова Ловушки, Райкоке, Матуа, Расшуа, острова Среднего, Янкича и Чёрные Братья (Артюхин и др. 2001, 2016б; Клумов 2001; Трухин 2003). Непонятен статус малой конюги на островах Анциферова, Онекотан и Броутона, для которых упоминается гнездование конюг, но не конкретизуются виды (Клумов 2001); у острова Броутона малая конюга встречена после завершения периода размножения, что не может быть подтверждением её гнездования на этом острове (Артюхин и др. 2001). На основании того, что малые конюги во время сезона размножения были встречены в море недалеко от берега, сделано предположение, что они также могут гнездиться на островах Маканруши и Симушир (Артюхин и др. 2001).
Утверждение о гнездовании малой конюги на острове Уруп (Austin, Kuroda 1953; Гизенко 1955; Кондратьев 1990; и др.), основано, вероятно, на публикации Ё.Ямасины (Yamashina 1931), в которой не приведены какие-либо ссылки на наблюдения. Возможными основаниями, видимо, послужили летние встречи малых конюг в прибрежной акватории острова: в коллекции Токийского императорского университета хранятся экземпляры, собранные 31 июля - 10 августа 1895 (Stejneger 1899), а также экземпляр взрослой птицы от 16 августа 1924 в коллекции Forestry and Forest Products Research Institute (Артюхин и др. 2001). Однако А.Г. Велижанин (1971, 1972 — по: Артюхин и др. 2001) проводит южную границу ареала малой конюги по острову Уруп на основании опросных данных, согласно которым небольшое число малых конюг гнездилось на островах Близнецы, находящихся у северного побережья острова Урупа, а также на добыче взрослой птицы 25 июля 1955 в заливе Наталии (Артюхин и др. 2001). Между тем, при обследовании острова в 2003 году достоверных свидетельств гнездования малой конюги зафиксировано не было (Артюхин 2003а,б). Южнее Урупа две взрослые малые конюги в период размножения были добыты в акватории южного Кунашира (Артюхин и др. 2001).
В акватории Охотского моря достоверно подтверждено гнездование вида только на острове Ионы (Нечаев, Тимофеева 1973; Харитонов 1980; Андреев и др. 2012). Все прочие указанные в литературе локации требуют дополнительного подтверждения и в настоящее время считаются спорными. Не вполне ясен статус малой конюги на Ямских островах. А.Г.Велижанин (1975, 1977, 1978), работавший там в 1974 году, оценивал численность малых конюг на островах Матыкиль и Атыкан в 1 тыс. особей на каждом; на последнем острове 4 июля из-под птицы коллек-тировано сильно насиженное яйцо. При последующих обследованиях острова Матыкиль (Кондратьев и др. 1993; А.Н.Головкин, устн. сообщ.) этот вид там отмечен не был. Обследование берегов островов Атыкан, Баран и Хатемалью и прилегающей акватории 3 августа 2016, как и все экспедиции, посещавшие Ямские острова после 1974 года (Голубова, Плещенко 1997; Зеленская 2009, 2017, 2018; Андреев 2016), малой ко-нюги там не обнаружили. Сильные приливные течения — характерный кормовой биотоп малой конюги — вокруг этих островов есть, а сами острова обеспечивают необходимые гнездовые биотопы и совершенно непонятно, почему этот вид там не встречается, тогда как на острове Ионы со сходными гнездовыми и кормовыми биотопами он гнездится. Кратко упоминается, что малая конюга гнездится по юго-западному побережью залива Шелихова и на полуостровах Кони и Пьягина (Васьковский 1956), однако последующие обследования данного района не обнаружили там этот вид (Кондратьев и др. 1993; Зеленская 2013, 2021а,б, 2023). Существование колонии малых конюг на островах в Пенжинской губе (Яхонтов 1973, 1974) не подтверждено; при обследовании островов и прилегающей акватории в июле – начале августа 2019 года малая конюга не встречена (Артюхин 2019). На Шантарских островах ситуации тоже непонятная. При обследовании островов Большой Шантар и Феклистова в 19241926 годах малая конюга не отмечена, но упоминается большая конюга (Дулькейт, Шульпин 1937). Только большая конюга отмечена на архипелаге во время работ в 1959, 1971 и 1972 годах (Яхонтов 1977). В то же время обследование архипелага, проведённое в 1991-1992 годах, показало, что малая конюга гнездится на островах Утичий, Птичий, Кусова, Прокофьева, Сахарная Голова и др. (Росляков, Росляков 1996), но в публикации не упоминается большая конюга. При просмотре фотографий на многочисленных сайтах, где туристы публикуют фотоотчёты о своих поездках на Шантары, были обнаружены изображения только больших конюг. По гидрологическим особенностям этого архипелага, характеризующимися сильными приливными течениями, гнездование малых ко-нюг кажется вполне вероятными.
Помимо достоверно подтверждённых мест гнездования, в литературе встречаются указания на возможное гнездование малой конюги в других районах тихоокеанского побережья Азии. Вид был включён в список гнездящихся птиц восточного побережья Камчатки (Аверин 1957), однако последующие исследования не подтвердили факта его размножения в данном регионе. Все документально зафиксированные встречи относятся исключительно к морским акваториям: 5 особей наблюдались 11 августа 1972 между бухтами Асача и Мутная (Вяткин 1981), а 21 июля 1977 птицы отмечены в заливе Корфа вблизи посёлка Вывенка (Лобков 1986).
На Сахалине малые конюги отмечались в заливе Терпения 24 июня 1948 и у мыса Анива в июле 1948 года (Гизенко 1955), однако факт гнездования на острове остаётся недоказанным. Тем не менее, потенциальные места размножения могут существовать в районе мыса Терпения, где 15 июня 1981 встречены 2 особи, а также на мысах Анива и Елизаветы (полуостров Шмидта) (Нечаев 1986). Высказанное ранее предположение о возможном гнездовании вида на острове Монерон, на Восточных островах, где птицы неоднократно встречались в течение июня 1973 года (Нечаев 1975), не получило подтверждения в ходе последующих исследований (Шибаев, Литвиненко 1996). Однако полностью исключать вероятность присутствия малой конюги в этом районе преждевременно, поскольку в опубликованных данных не указано время суток, когда проводились исследования, давшие отрицательный результат.
Основными факторами, определяющими очертание гнездовой части ареала малой конюги, являются особенности кормовой специализации вида, связанные с районами приливных возмущений. Зимой птицы покидают окрестности только тех колоний, где образуется ледовый покров.
Динамика гнездовой части ареала . Малые конюги, по-видимому, очень подвержены хищничеству со стороны интродуцированных лисиц, песцов Vulpes lagopus и крыс, отчасти потому, что молодые особи, в отличие от других конюг Aethia , возвращаются на сушу после слёта (Зубакин, Конюхов 2001; Williams et al . 2003). Интродукция на острова Курильской и Алеутской гряд наземных хищников привела к сокращению как ареала, так и численности вида. После уничтожения лисиц и песцов этот вид, вероятно, постепенно восстановит большую часть своего прежнего ареала, за исключением островов, где теперь обитают серые крысы (Byrd et al . 2005), уничтожение которых связаны с большими финансовыми затратами.
Свидетельств о численности малых конюг на Алеутских островах до того, как на них стали завозить песцов, нет, но есть немногочисленные данные того периоды, когда интродукция только начиналась. Натуралист Люсьен Тернер (Turner 1886) писал, что в 1880-1881 годах малая конюга считалась «довольно многочисленной» на Ближних островах и «обычной» на центральных Алеутах. Обследование всего Алеутского архипелага, проведённое в 1911 году, не выявило присутствия малой ко-нюги (Bent 1919). Примечательно, что в этой публикации исследователь не упомянул и белобрюшку, которая является достаточно обычным видом на островах гряды (Sowls et al . 1978; Jones et al . 2020). Никаких других данных о численности малой конюги не существовало до 1936 года. В это время популяция песцов на островах достигла своего пика, а популяция малых конюг – своего минимума; малые конюги исчезли на Ближних островах. В то время на Алеутах гнездилось всего несколько тысяч этих конюг (Williams et al . 2003). Тщательное обследование этих островов в 1972-1974 годах, специально направленное на поиск малой конюги, показало, что по результатам учётов птиц в море там обитало около 25 тыс. особей этого вида (Byrd, Gibson 1980). Летом 1975 года единичные птицы были встречены в прибрежных водах как островов Атту, так и Агатту (Ближние острова) (Kessel, Gibson 1978). Уничтожение лисиц, песцов и крыс, завезённых на Алеутские острова, которое ведётся в настоящее время персоналом Аляскинского морского заказника (Alaska Maritime NWR), несомненно, приведёт к увеличению как количества мест гнездования, так и численности и, видимо, к расширению ареала этого вида до первоначального размера (Williams et al . 2003).
На Курильских островах прослеживается та же тенденция. До заселения островов различными чужеродными наземными млекопитающими на всех островах к северу от острова Урупа большие и малые конюги были многочисленными (Snow 1897). Вот какой была авифауна островов Ушишир до того, как туда добрались преобразователи природы. В конце XVIII века Григорий Шелихов (1793) писал, что на острове «зверей не водится ни каких» и «остров сей безлесен, а вокруг гор и хребтов по низким местам разных морских птиц плодится великое множество, как то: ары [кайры – Н.Б.Конюхов], глупыши, топорки, урилы [бакланы], курукуры [скорее всего, большие конюги], турутуры [малые коню-ги], разные чайки, серые гуси и величиною с косаточку птичка, с сероватыми на крыльях и спине перьями, белым брюхом и кривым носом, называемая кагарка [скорее всего, сизая качурка Hydrobates furcatus]».
Спустя век на острове ничего существенно не изменилось. Генри Джеймс Сноу (Snow 1897) так описывает авифауну острова: «Это место – подлинный рай для птиц. Здесь нет ни лисиц, ни других наземных хищников, что и объясняет невероятное количество пернатых обитателей. Бургомистры (Larus glaucus) гнездятся по всему острову, особенно у горячих источников и на участках с сернистой почвой. Я часто собирал их яйца и варил прямо в кипящих источниках. На уступах скал десятки тысяч кайр (Uria troile и U. brünnichi) отложили яйца и выводят птенцов, а в то же время между ними тут и там расположились многочисленные колонии моевок (Rissa tridactyla) , чьи гнёзда ютятся на недоступных карнизах и в расщелинах.
Сотни тысяч глупышей (Fulmarus pacificus) занимают каждый пригодный травянистый холмик и выступ на крутых склонах – как внутри кратера, так и снаружи. Миллионы мелких чистиковых нескольких видов (Phaleris cristatella и P. mystacea были самыми многочисленными) откладывают яйца во всевозможных углублениях, трещинах и под валунами по всему периметру острова.
С наступлением вечера конюги отправляются на свои лётные упражнения. Они собираются в стаи из многих тысяч особей, и сотни таких стай одновременно заполняют воздух, образуя тучи, которые буквально затмевают небо. Птицы кружат вокруг острова и над ним, то взмывая высоко над горами, то стремительно пикируя к самой воде, чтобы вновь взлететь и виртуозно разойтись в воздухе – каждая стая движется как единый организм, – словно исполняя сложные фигуры замысловатой кадрили.
По вечерам кайры, часто в компании тупиков, тоже совершают свои тренировочные полёты, но гораздо более степенно. Они летят ровной бесконечной лентой, размеренно кружа вокруг острова. Такие чётко организованные полёты характерны только для видов с относительно тяжёлым телосложением и короткими крыльями.
На острове также в большом числе гнездятся топорки, чёрные чистики и бакланы. Ипатки, белобрюшки, обыкновенные старики, вилохвостые и северные качурки были обычны, но не многочисленны. Каме-нушки весьма многочисленны, однако мне ни разу не удалось обнаружить их кладки. Дикие гуси (Bernicla hutchinsi) гнездятся здесь в не- большом количестве. Из наземных птиц здесь обитают лишь вороны, соколы, трясогузки и крапивники».
Распространение в море летом . Обследование, проводившееся вдоль всей Алеутской гряды в 1972-1974 годах, показало, что наибольшее число малых конюг было обнаружено у Лисьих островов в апреле 1974 года (перед сезоном размножения), а также 24-27 мая 1972 и 3 июля 1973 (во время сезона размножения). В районе пролива Бабий в 1974 году было учтено почти 60% из 13118 малых конюг, встреченных в акватории Лисьих островов; приблизительная плотность составляла 1000 ос./км2. Там же в апреле 1976 года плотность малых конюг оценивалась в 265 ос./км2 (Byrd, Gibson 1980).
В акватории Командорских островов малые конюги кормятся как правило в пределах шельфа, где в июне-июле 2004 года средняя плотность составляла 11 ос./км2, а в августе – 2 ос./км2. Крупные кормовые скопления наблюдали у северного побережья острова Медный и на Китоловной банке (Artukhin 2006; Артюхин 2016а).
Конюги Aethia доминируют в прибрежных водах Курильских островов, встречаясь в основном у Средних Курильских островов в пределах гнездовой части ареала: поблизости от их крупных колоний – в проливах Экарма, Головнина, Буссоль и Уруп. По данным исследований 20002003 годов плотность распределения конюг в летний период в прибрежных водах Северных, Средних и Южных Курильских островов соответственно составляла 1.527, 206.079 и 0.066 ос./км2. При этом доля малой конюги достигала 5-8% (Артюхин 2003б).
В Охотском море летом плотность и численность конюг увеличивается с юга на север; в его северной части, включая залив Шелихова, ко-нюги летом преобладают над другими чистиковыми, но за пределами шельфа почти не встречаются (Шунтов 1972, 1998). Можно предположить, что среди этих конюг есть и малые конюги.
Характер распространения и его причины . Несомненно, ключевой адаптацией малой конюги была добыча корма в приливно-отливных возмущениях водных масс. Все достоверно известные места гнездования вида находятся именно там, где подобные явления очень сильны (Деревнин и др. 1968, 1976; Андреев и др. 1969). В пределах гнездового ареала можно ожидать гнездование малой конюги на островах, вокруг которых существуют сильные приливно-отливные течения.
Миграции и зимовки
В разных частях ареала в зависимости от местных климатических условий птицы ведут себя по-разному. «По словам местных жителей, малые конюги зимуют на Алеутских островах и прячутся на ночь в каменные расщелины, из-за чего становится жертвой голубых песцов» (Murie 1959). Исследования с применением геологгеров на острове Булдырь в
2013-2015 годах показали, что малые конюги из колонии, где проходило их мечение, не мигрируют, а остаются в акватории острова всю зиму и ночуют на суше. В это время птицы перемещаются то на юг, то на север от острова, где они, вероятно, кормятся в приливных возмущениях. Перемещений на запад и восток вдоль Алеутской гряды отмечено не было (Schacter 2017; Schacter, Jones 2018). На Средних Курильских островах малая конюга, скорее всего, тоже оседла; как зимой, так и летом она большими стаями встречается в 5-10 милях от берега (Гизенко 1955). Часть популяции, гнездящаяся в Охотском море и, видимо, на Командорских островах, откочёвывает на юг. Поскольку в море различать ко-нюг крайне трудно, то в публикациях часто упоминаются просто «ко-нюги». Часть их, можно предположить, относилась к малым конюгам.
Природа и причины миграций . Приливно-отливные течения регулярны и не зависят от времени года, поэтому в тех районах, где зимой море не покрыто льдом, в частности, в окрестностях Алеутских и Средних Курильских островов (рис. 10), для малой конюги нет необходимости совершать сезонные перемещения, и она остаётся на зиму в районе колоний. В замерзающей зимой северной части Охотского моря малая конюга – кочующая или даже перелётная птица. Однако и там благодаря сильным приливным течениям постоянно имеются значительные пространства открытой воды в виде трещин и полыней, где неоднократно отмечались какие-то конюги (Трухин, Косыгин 1986). Поскольку малые конюги не боятся плавучих льдов (Велижанин 1971), можно предположить, что по крайней мере часть из этих конюг были малыми. Однако вероятность того, что этот вид зимует в открытой акватории Охотского моря, маловероятна, поскольку малые конюги кормятся в приливных возмущениях, которые возникают у островов и банок.
Алеутские острова . Исследования в негнездовой период (осень, зима и весна 1986/87 года) выявили, что в районе пролива Унимак встречено около 50% от численности аляскинской популяции малой конюги (Troy, Bradstreet 1991), которая была известна на тот момент. Это говорит о том, что данная акватория имеет большое значение для вида.
В проливе Унимак вне сезона размножения малые конюги всегда присутствовали и всегда преобладали в приливно-отливной водной массе, хотя были отмечены и в других водных массах, но в относительно небольшом числе (рис. 11). Осенью пролив Акутан является основной акваторией, где собираются тысяч малых конюг. Также малые конюги были тесно связаны с островами Креницына. Их крупные скопления отмечались южнее проливов между островами. Зимой большие и малые ко-нюги образовывали значительные скопления. Наиболее многочисленным видом в этот период была большая конюга. Малые конюги также встречались в большом числе, однако их плотность была на порядок ниже. Конюги были широко распространены в проливе Акутан, при этом наибольшие концентрации отмечались в проливе Бабий. Обычно скопления больших и малых конюг пространственно разделены, но иногда наблюдались и смешанные скопления.
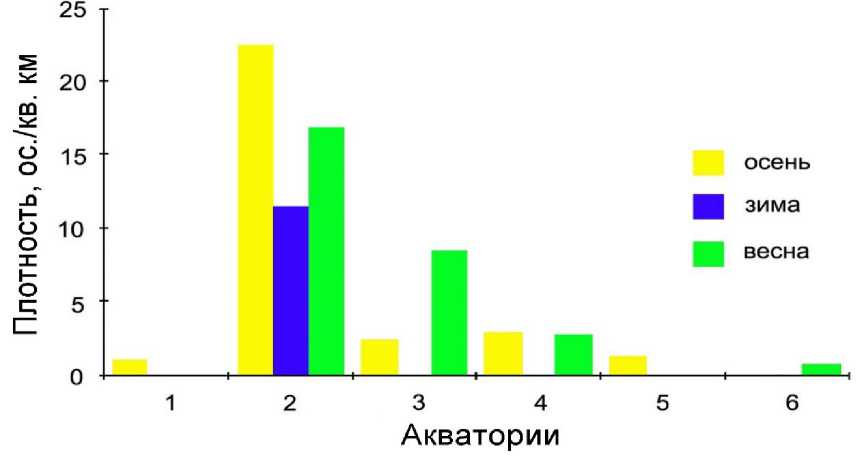
Рис. 11. Средняя плотность малых конюг (ос./км2) в разных водных массах осенью 1986 года, зимой, и весной 1987 года в районе пролива Унимак (Аляска). Акватории: 1 – шельфового склона (Shelf Break Water), 2 – приливно-отливная (Tidally Mixed Water), 3 – залива Аляска северная (Gulf of Alaska Water northern (Bering Sea), 4 – залива Аляска юж-ная (Gulf of Alaska Water south), 5 – Аляскинская прибрежная северная (Alaska Coastal Water north (Bering Sea)), 6 - Аляскинская прибрежная южная (Alaska Coastal Water south) (Troy, Bradstreet 1991)
Кроме островов Креницына, другим районом скопления малых ко-нюг был пролив Дербенский. Основное место зимовок больших конюг находилось в прибрежных водах к северу от острова Унимак; в проливе Акутан их скопления были заметными, но рассеянными. Весной конюги по-прежнему были многочисленны. Районы скопления больших конюг не совпадали с теми, которые были зимой. Этот вид встречался в основном к югу от островов Креницына, напротив проливов. Малые конюги были распространены более широко, чем зимой; их находили по всему проливу Аватанак и в Беринговом море напротив проливов островов Кре-ницына. Южнее пролива Акутан была относительно небольшая зона перекрытия скоплений этих двух видов. Средняя плотность малых ко-нюг осенью, зимой и весной соответственно составляла: 16.289, 11.007, 15.34810 ос./км2; в скоплениях достигая максимума – 2375.81, 1337.43, 804.25 ос./км2 (Troy, Bradstreet 1991).
На центральных Алеутских островах зимой малые конюги встречаются в проливах между островами: в южной части пролива Адак 18 февраля 1999 встречено 160 особей в стаях до 35 птиц, а 19 февраля в северной и центральной частях пролива Адак – 353 особи в стаях до 50 птиц, однако о распространении конюг в это время года известно недостаточно (Gibson, Byrd 2007). Вне скоплений встречается крайне редко: 1 особь встречена в проливе Канага 19 февраля 1999, группы из 6 особей к северо-востоку от острова Малый Ситкин и 8 особей в проливе Ам-читка 27 февраля 1999 и по 1 птице у островов Амчитка 27 февраля 1954 и Адак 13 ноября 1974 (Kessel, Gibson 1978; Gibson, Byrd 2007).
Малая конюга – единственный вид рода Aethia , который зимой на западных и центральных Алеутских островах встречается с плотностью, аналогичной летней, а на восточных Алеутах её плотность зимой больше, чем летом. В акватории Булдыря в 1998 году плотность малых ко-нюг в июле – начале августа и в марте составляла 10.8 и 24.2 ос./км2 соответственно; у острова Касаточий летом 9.0 (1996 год), 18.9 (1997), 26.7 ос./км2 (2003) и зимой – 1.5 (1998), 2.8 ос./км2 (1999 год); у острова Аиктак летом – 13.5 (1995 год), 4.5 (1996), 5.9 ос./км2 (1998 год), зимой – 248.7 (1997 год), 63.7 (1998), 92.2 ос./км2 (1999 год) (Renner et al . 2008).
Распространение малых конюг в границах Алеутского архипелага всё ещё остаётся не до конца ясным. На западе архипелага, на Булдыре, птицы остаются в прибрежной акватории в течение всего года, и их перемещения зимой на запад или восток вдоль гряды не отмечено (Scha-cter, Jones 2018). Восточнее, в акватории острова Касаточий, плотность птиц летом и зимой оказалась сопоставимой (Renner et al . 2008), что позволяет предположить осёдлость местной популяции. Однако в акватории восточных Алеутских островов (остров Аиктак) малых конюг зимой намного больше, чем летом (Renner et al . 2008). Именно здесь, видимо, происходят местные сезонные кочёвки вида. Это подтверждается исследованиями 1972-1974 годов. В период размножения в акватории Четырёхсопочных островов 24 мая – 7 июня 1972 учтено 12100 малых конюг, тогда как в конце зимы, 4-30 апреля 1974, здесь встречено лишь 2519 особей. При этом зимой птицы в этом районе встречались несколько западнее, чем летом. Обратная ситуация отмечена в акватории Андреяновских островов, где в сезон размножения обнаружено менее 800 конюг, а до его начала – более 9000. Почти 70% этих птиц встречено близ югозападной оконечности острова Большой Ситкин. В ходе интенсивного обследования акватории у этого и всех близлежащих островов в июле 1971 года малые конюги обнаружены не были. Следовательно, птицы, замеченные здесь до начала сезона размножения в 1974 году, гнездились, вероятно, на Четырёхсопочных островах. После завершения сезона размножения 21-22 сентября 1974 восточнее этих островов встречено около 2000 малых конюг, а 27 сентября – 638 птиц восточнее Лисьих островов. По-видимому, эти птицы зимуют недалеко от места своего гнездования (Byrd, Gibson 1980). Логично предположить, что с местом гнездования связана только часть популяции. Поскольку привязаны к территории больше самцы, по крайней мере у больших конюг (Зубакин 1990; Зубакин, Зубакина 1993), не исключено, что именно они остаются зимовать вблизи колоний, а самки и неполовозрелые птицы перемещаются в смежные районы (Зубакин, Конюхов 2001).
Командорские острова . В акватории Командорских островов малые конюги зимуют или по крайней мере проводят какую-то часть времени вне сезона размножения. Кормящиеся конюги стайками по 2-4 особи отмечались вдоль скалистых берегов острова Беринга в декабре – начале января. Позже птиц стало меньше, и последняя добыта 30 января (Stej-neger 1885). Вполне вероятно, что они перемещаются в акваторию острова Медный. Г.Х.Иогансен (1934) отмечает, что по наблюдениям и сборам на Командорах малая конюга чаще встречается зимой, чем летом, а С.В.Мараков (1965) считает её здесь зимующей.
Камчатка . У восточного побережья Камчатки, видимо, немногочисленна. Одиночных малых конюг неоднократно видели в Кроноцком заливе в октябре – начале ноября 1973 и 1975 годов. В это же время мёртвых птиц находили на побережье вблизи посёлка Жупаново (Лобков 1980, 1986). Малая конюга добывалась, вероятно, зимой в Авачинской губе (Bergman 1935 – по: Дементьев 1951). У южной оконечности полуострова в Первом Курильском проливе в феврале 1982 года над шельфом в открытых водах отмечено около 15 стай конюг численностью до нескольких сотен птиц в каждой (Трухин, Косыгин 1986), вероятно, тут могли быть и малые конюги.
Курильские острова . На Курилах малая конюга встречается круглогодично (Yamashina 1931; Austin, Kuroda 1953; Гизенко 1955). Осенью больше всего этих конюг держится в средней части Курильской гряды. В сентябре 1947 года в большом числе отмечалась у островов Матуа, Си-мушир и Онекотан, а 4 и 5 октября большие скопления отмечены близ Итурупа в его средней части, в 5-6 милях от восточного побережья; в это время птицы были многочисленны также у берегов острова Чирпой, у северной оконечности Урупа. В январе-феврале малые конюги отмечены у островов Матуа, Пуффина (Топорковый) и Симушир. В сентябре и начале октября, у берегов Кунашира и Шикотана малые конюги не наблюдались. Вероятно, они появляются близ этих островов только в суровые зимы или при изменении ледовой обстановки (Гизенко 1955). Илья Гаврилович Вознесенский наблюдал малую конюгу в декабре, январе и феврале на Курильских островах, особенно на Урупе (Stejneger 1899).
Охотское море. В Охотском море конюги держатся всю зиму; в начале зимы их стаи встречаются во льдах всюду, а в феврале численность птиц несколько снижается. В отличие от других чистиковых, конюги во льдах встречались всегда в стаях численностью не менее нескольких десятков птиц (Трухин, Косыгин 1986). Поскольку малые конюги не боятся плавучих льдов (Велижанин 1971), можно предположить, что по крайней мере некоторые из этих конюг были малыми. В декабре такие стаи не представляли редкости в глубине льдов в северо-западной части моря. В первой половине декабря здесь наблюдались плотные, почти сплошные льды с очень редкими узкими трещинами, в которых и дер- жатся конюги. В феврале 1982 года конюги отмечались в заливе Шели-хова и у острова Ионы (Трухин, Косыгин 1986).
Сахалин . В акватории острова малая конюга – редкий пролётный и зимующий вид (Шунтов 1965; Нечаев 1986, 1991).
Японское море . Проникновение малой конюги в Японское море, скорее всего, случайно. Зимой основу населения в северной части этого моря у берегов Приморья, Сахалина и Хоккайдо составляют конюги, которые во второй половине марта начинают откочёвку на север (Шунтов 1972). Есть ли среди них малые конюги, пока не неизвестно.
Япония . Малая конюга редка и случайно встречается только в водах северной Японии (Brazil 2009). Малые конюги добираются до побережья Хонсю и Сикоку (Yamashina 1931). О.Л.Остин и Н.Курода (Austin, Kuroda 1953) пишут, что утверждения о встречах малых конюг на Хонсю и Сикоку сомнительны. Установить источник регистрации вида на Сикоку им не удалось. Экземпляр, добытый у Симоды (Хонсю) в 1853 году и считавшийся малой конюгой, на самом деле оказался большой конюгой, поскольку из-за некорректной синонимизации из Uria mystacea превратился в Aethia pygmaea . Исследования в акватории Хоккайдо зимой не подтвердили присутствия здесь малых конюг (Austin, Kuroda 1953). Первая достоверная встреча этого вида произошла у восточного побережья Хонсю в префектуре Мияги 8 февраля 1957. На Хоккайдо в марте 1968 и феврале 1980 года малые конюги отмечались у мыса Очииси, а 28 февраля 1984 – у мыса Носаппу (Brazil 1991).
На восточном побережье Пацифики (центральная и юго-восточная Аляска, Британская Колумбия, Вашингтон, Орегон и Калифорния) малая конюга на зимовках не отмечалась.
Залёты
Малые конюги встречены на острове Святого Лаврентия (рис. 10). Одна птица, морфологические признаки которой ближе к особям курильской, чем алеутской популяции, добыта у посёлка Гэмбэлл 9 июля 1931 (Feinstein 1959), хотя есть сомнение, действительно ли она происходит с этого острова (Friedmann 1932). Ещё одна птица отмечена у мыса Татик 20 июля 1971 (Johnson 1974). Малая конюга сфотографирована на острове Святого Георгия (острова Прибылова) 31 мая 2015*. Экземпляр из посёлка Бетель (дельта Юкон-Кускоквима) относится к алеутской популяции (Feinstein 1959), кроме этого, одна птица встречена 4 июля 1973 у острова Амака в Бристольском заливе (Kessel, Gibson 1978). К югу от Аляски малая конюга – случайный гость на западном побережье Северной Америки. Взрослая птица встречена 7 июня 2001 в водах залива Тасу на западном побережье острова Морсби в архипелаге Хайда- ;
Гуаи (острова Королевы Шарлотты, Британская Колумбия). В штате Вашингтон одна малая конюга встречена в Пенн-Коув (округ Айленд) 16-17 мая 1999. По залётам в Орегон и Калифорнию подтверждённых данных нет (Toochin, Cecile 2018).
В азиатской части ареала залёты малой конюги малочисленны. В Приморье останки малой конюги без головы обнаружены 8 июня 1962 на берегу бухты Ильмовая Уссурийского залива (Назаров 2004), однако непонятно, когда птица туда попала и была ли это действительно малая конюга. Добыча двух птиц в феврале 1883 года в Нагасаки (Гизенко 1955) вызывает большие сомнения, поскольку это место находится значительно южнее достоверных встреч этого вида в водах Японии и даже примерно в тысяче километров южнее самой южной встречи большой конюги – более обычного вида в водах Японии (Brazil 1991).
Численность
Численность малых конюг на Алеутских островах составляет не менее 116 тыс. особей, распределённых следующим образом: Ближние острова – 500 особей, Булдырь – 30 тыс., Кыска – Канага – 500, Адак – Атка – 30 тыс., Сигуам – 35 тыс., Четырёхсопочные острова – 10 тыс., острова Умнак – Унимак – 10 тыс., острова Эгг – менее 1 тыс особей. Эти оценки, кроме Булдыря, основаны на учётах наибольшего числа птиц в море в период размножения, когда многие из них посещали места гнездования, поэтому эту численность следует рассматривать как минимальную (Williams et al . 2003; Byrd et al . 2005; Jones, Seneviratne 2005).
Численность командорской популяции малой конюги оценивается в десятки тысяч особей (Головкин 1990; Артюхин 2016а). Хотя по результатам исследований 1986-1994 годов она оценена в более чем 2 тыс. особей (Артюхин 1999); летом 2017 года на острове Медный численность малой конюги оставалась прежней, а возможно, даже увеличилась (Пилипенко 2021).
На Средних Курилах малая конюга гнездится на большинстве островов и крупных скал. Её колонии находятся на следующих островах: Экарма (первые десятки тысяч), Чиринкотан (несколько тысяч), Шиа-шкотан (нескольких сотен), острова Ловушки (первые сотни), Райкоке (первые десятки тысяч), Матуа (несколько тысяч), Расшуа, острова Среднего, Янкича (первые десятки тысяч; значительно уступает по численности большой конюге) и Чёрные Братья (первые тысячи). Для точной оценки численности курильской популяции малой конюги данных недостаточно, но она вряд ли превышает 50 тыс. особей (Артюхин и др. 2001; 2016б).
Оценки численности популяции малых конюг на острове Ионы значительно разнятся: в 1971 году численность оценивалась в 100 тыс. особей (Нечаев, Тимофеева 1973), а в 1974 – 1 тыс. особей (Харитонов 1980).
Наиболее обоснованная оценка, которая основана на учётах птиц на мониторинговых площадках и экстраполяции на площадь занимаемых видом биотопов, осуществлена в первой половине июля 2009 года и составила около 20.5 тыс. особей, однако авторы более осторожно оценивают её в 10-12 тыс. особей (Андреев и др. 2012).
Суммируя все известные данные о численности малой конюги, можно заключить, что в настоящее время общая численность населения этого вида составляет около 200 тыс. особей.
Я искренне благодарен сотрудникам Аляскинского морского национального природного заказника (Alaska Maritime National Wildlife Refuge) за предоставленную возможность работы на острове Булдырь, а также за техническую поддержку исследований. Я крайне признателен Ю.Б.Артюхину, В.А.Зубакину, К.Е.Литвину и С.П.Харитонову за замечания и ценные советы во время написания данной публикации.
