Птицы России и сопредельных стран: малая конюга Aethia pygmaea (Gmelin, 1789). 2. Образ жизни
. 2. Образ жизни")
Автор: Н.Б. Конюхов
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 2575 т.34, 2025 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140312818
IDR: 140312818
Birds of Russia and neighboring countries: the whiskered auklet Aethia pygmaea (Gmelin, 1789). 2. Natural history
Текст статьи Птицы России и сопредельных стран: малая конюга Aethia pygmaea (Gmelin, 1789). 2. Образ жизни
*
†
%
I П 111 IV V VI VU VIII IX X I П III IV V VI VU VIII IX X
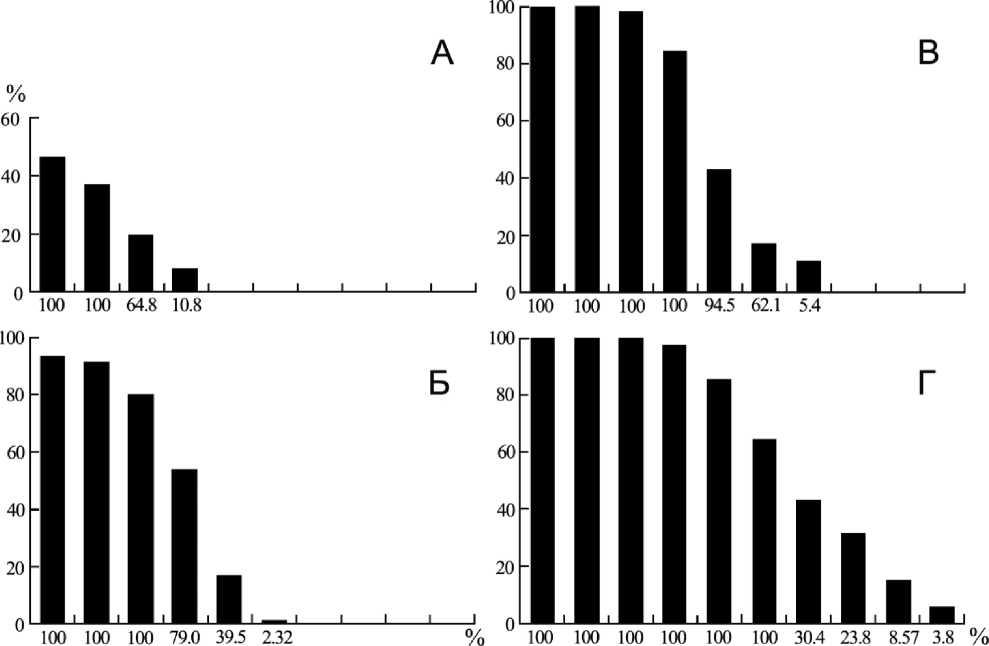
Рис. 10. Линька первостепенных маховых перьев у малой конюги.
А – насиживание, Б – выкармливание, В – слёт птенцов, Г – постгнездовой период.
I–X – первостепенные маховые. По оси абсцисс – относительное количество птиц (%), у которых данное маховое находилось в стадии линьки, по оси ординат – средняя относительная длина (%) данного нового махового пера (по: Конюхов 2001)
На острове Егг (восточные Алеуты) 10-25 августа 1992-1993 у малых конюг линька первостепенных маховых уже шла. Предположительно, она началась в июле (Pitocchelli et al . 2003). У командорских малых ко-нюг линька начинается в последней декаде июля со смены верхних кроющих крыла и внутренних первостепенных маховых (Stejneger 1885), на Курилах – тоже в июле (Гизенко 1955). Сроки линьки у малых конюг на острове Ионы, по-видимому, сходны с таковыми у курильских птиц. Взрослая птица, добытая на острове 1 сентября, интенсивно линяла; менялись первостепенные маховые и контурное оперение по всему телу, старые перья хохолка выпали, а новые были в роговых чехликах у основания и достигали длины 0.7-0.8 см (Нечаев, Тимофеева 1973). Однако необходимо отметить, что во всех этих случаях (кроме острова Бул-дырь) начало линьки могло быть пропущено, так как не проводилось периодических отловов птиц, целью которых было изучение именно линьки. Реально она могла начаться на несколько недель раньше указанных выше сроков.
Окончание линьки определить гораздо труднее, чем её начало. Видимо, это конец сентября – начало октября. У птиц с Булдыря внешние первостепенные маховые, судя по скорости их роста, должны достигнуть нормальной длины к началу октября (Конюхов 2001). Семь самцов, добытых 25 сентября, видимо, в районе Средних Курильских островов (точное место не указано), уже полностью закончили линьку (Гизенко 1955).
Первостепенные маховые и их кроющие . Линька первостепенных маховых идёт центробежно. У птиц сразу выпадает 2-3 самых внутренних первостепенных маховых. Когда новые маховые, выросшие на месте выпавших, достигают своей нормальной длины, выпадают следующие 2-3 пера и т.д. К концу августа у некоторых птиц уже выпадают все первостепенные маховые. Скорость их роста равна 1.67±0.69 мм/сут ( SD , n = 89). Линька этой категории маховых должна завершиться к началу октября (рис. 9).
Схема линьки маховых у птиц на острове Егг была сходна с таковой у птиц на Булдыре. Она начиналась с самых внутренних первостепенных маховых перьев и постепенно продвигалась наружу. Больших различий в линьке самцов и самок не обнаружено. У всех взрослых самок 10-25 августа 1992-1993 1-5-е первостепенные маховые были новыми. У 10 из 18 птиц сменилось 6-е первостепенное маховое. У 6 из оставшихся 8 самок длина нового 6-го первостепенного махового составляло 70-90% от его нормальной длины. Состояние 7-го первостепенного варьировала в наибольшей степени; у некоторых птиц оно было новым, в то время как у других находилось на разных стадиях роста, а у части птиц оставалось старым. У большинства самок 8-е первостепенное маховое оставалось старым, и у всех них старыми были 9-е и 10-е первостепенные маховые. Линька первостепенных маховых у самцов была сходной с таковой самок: 1-5-е первостепенные маховые были новыми у всех самцов, а 6-е было новым у 10 из 16 самцов. Как и у самок, 9-е и 10-е первостепенные маховые у большинства самцов были старыми, а 7-е и 8-е находились на разных стадиях роста (Pitocchelli et al . 2003).
На Командорах, как и на острове Булдырь, линька первостепенных маховых перьев происходит в конце периода выкармливания птенцов и также начинается с внутренних маховых. У птиц, добытых в гнёздах 21 июля, 4 или 5 внутренних первостепенных маховых находились на разных стадиях роста: три самых внутренних полностью выросли и хорошо отличались от выцветших старых маховых. Кроме того, шла линька контурного оперения по всему телу (Stejneger 1885).
Большие верхние кроющие первостепенных маховых у птиц сменяются в той же последовательности, что и первостепенные маховые. Каждое большое верхнее кроющее выпадает одновременно или почти одновременно с соответствующим первостепенным маховым. Однако при осмотре птицы на поверхности крыла растущих больших верхних кроющих бывает видно на 1-3 меньше, чем первостепенных маховых, поскольку они прикрыты средними верхними кроющими.
Средние верхние кроющие первостепенных маховых линяют в той же последовательности, что и первостепенные маховые. Их линька начинается в конце июля и к концу августа только единичные перья этой категории остаются старыми.
Линька больших нижних кроющих первостепенных маховых начинается в середине августа. В отличие от верхних кроющих, упомянутых выше, она идёт в противоположную сторону – навстречу линьке самих первостепенных маховых, то есть центростремительно, от 10-го к 1-му первостепенному маховому. В конце августа у птиц новыми были 1-3 больших нижних кроющих первостепенных маховых пера.
Смена средних нижних кроющих первостепенных маховых у большинства птиц идёт сразу с двух сторон – от карпального сгиба (центростремительно) и от переднего края крыла (центробежно); к концу августа около 80% средних нижних кроющих первостепенных уже новые.
Второстепенные маховые и их кроющие . Схема смены второстепенных маховых такая же, как и у первостепенных, с тем лишь отличием, что их линька идёт центростремительно, начинаясь с дистальных перьев. Второстепенные маховые обычно начинают выпадать после того, как новые 6-е и 7-е первостепенные маховые достигают примерно половины своей нормальной длины. У большинства размножающихся птиц это происходит в первой декаде августа. У неполовозрелых птиц в это время уже полностью выросло 1-е второстепенное маховое, а 2-5-е находились на разных стадиях роста. В третьей декаде августа у всех пойманных птиц, как неполовозрелых, так и взрослых, линяли второстепенные маховые, различалось только число выпавших и растущих перьев, а старыми оставались только 3-4 самых внутренних пера.
У большинства самок 10-25 августа 1992-1993 на острове Егг происходила линька второстепенных и рулевых, но ни одно из них не было заменено полностью (Pitocchelli et al . 2003).
На Булдыре с середины июля, то есть раньше, чем сами маховые, у птиц начали сменяться большие верхние кроющие второстепенных маховых. Ко времени выпадения первых второстепенных маховых у птиц уже не было 6-7-го, а иногда и всех больших верхних кроющих, причём первые 4-5 из них были уже новыми. Направление линьки этих кроющих совпадало с таковой второстепенных маховых, то есть было центростремительным. Они выпадали более интенсивно, чем сами эти маховые, то есть по 4-5 перьев одновременно. К началу августа смена больших верхних кроющих второстепенных маховых завершилась.
Большие нижние кроющие второстепенных маховых меняются позже, чем другие кроющие второстепенных маховых. В середине августа они у всех птиц были старыми. Средние нижние кроющие второстепенных маховых менялись без определённой последовательности. К концу августа у птиц уже преобладали новые перья.
Мелкое оперение крыла . Перья крылышка выпадают одновременно с 6-7-м первостепенным маховым. Мелкое оперение на крыле начинает сменяться в конце июля – начале августа без какой-либо определённой последовательности; оперение на пропатагиуме меняется чуть позже – в конце августа.
Рулевые . Смена рулевых перьев происходит в августе. Интересно то, что рулевые выпадали без какой-либо определённой последовательности и в очень сжатые сроки. У разных птиц, пойманных в середине августа, можно было встретить самую всевозможную комбинацию старых рулевых перьев: крайняя и центральная пара, только центральная пара или только крайняя, или же разные пары в беспорядке; число старых перьев могло быть и нечётным. В это время ни у одной из птиц не было никаких следов растущих новых рулевых.
Хотя старые рулевые перья выпадали в беспорядке, новые всегда начинали расти с дистальной пары. Различия в длине новых растущих рулевых перьев внешней и внутренней пар не так значительны, как у растущих маховых перьев. В конце августа рули у разных птиц как правило находились на разных стадиях роста – от полного их отсутствия на поверхности оперения до хорошо развитых 7 новых пар. Предположительно, линька рулевых у большинства птиц должна завершиться во второй половине сентября. Как исключение, в конце августа была поймана птица, у которой все рулевые ещё были старыми, однако у подавляющего большинства малых конюг в это время рулевые или уже выпали, или остались только единичные старые перья.
Контурное оперение . Линька контурного оперения идёт одновременно с линькой маховых и их кроющих. Она особенно заметна на голове, поскольку приводит к значительному изменению внешности птицы за счёт выпадения перьев косиц и султана, о линьке которых будет сказано ниже. В конце июля начинают выпадать перья на лбу по границе с рамфотекой, однако площадь, охваченная линькой, здесь незначительна. В середине августа у некоторых птиц на лбу уже росли новые перья, а зона линьки продвигалась к макушке. В конце августа интенсивная линька оперения началась перед глазом.
Украшающие образования. Схема расположения украшающего оперения представлена на рисунке 11. Султан является первым из украшающих образований, который начинает линять – его перья выпадают в начале июля. В конце месяца у некоторых птиц в султане остаётся только по 1-2 старых длинных пера. У некоторых взрослых малых конюг в начале августа султана уже нет, однако ещё до последней декады августа встречаются особи с длинным султаном, состоящим из более чем 5 старых перьев. Птицы с новым хохолком начинают встречаться с середины августа. К концу августа его длина у некоторых птиц уже составляла около 15 мм (Конюхов 2001). Новый султанчик длиной 7-8 мм и с роговыми чехликами в основаниях перьев был у птицы с острова Ионы, добытой 1 сентября (Нечаев, Тимофеева 1973). Видимо, к концу октября у всех птиц длина хохла достигает нормальных размеров. Е.В.Козлова (1957) сдвигает эти сроки к началу декабря.
Линька косиц тесно связана с линькой контурного пера. Напомним, что перья косиц – это не контурные перья, а filoplumulae. У разных птиц этот процесс менее синхронизирован, чем линька маховых, однако общие тенденции все же проследить удалось.
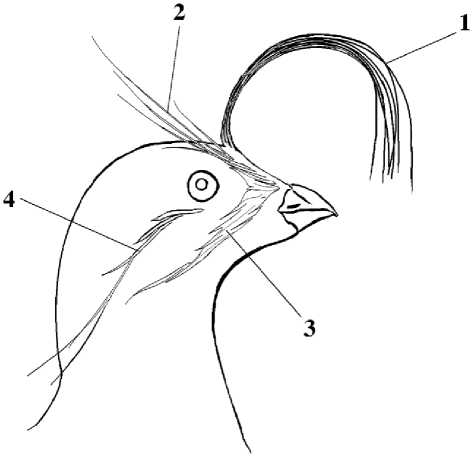
Рис. 11. Украшающие образования малой конюги. 1 – султан (хохолок); косицы: 2 – фронтальная, 3 – риктальная, 4 – аурикулярная (по: Конюхов 2001)
Косицы начинают линять в начале августа. Наиболее ранней редукции подвергается фронтальная косица за счёт выпадения самых длинных filoplumulae, растущих в её средней части, поскольку именно эти перья находятся ближе к зоне линьки контурного пера, распространяющейся по голове от клюва к затылку. У некоторых птиц в середине августа во фронтальной косице оставались только очень длинные filoplu-mulae в её задней части и короткие в передней, а в средней их уже нет. В конце августа у большинства птиц эта косица, как правило, состоит только из небольших filoplumulae в её передней части и 1-2 filoplumulae в задней части; у некоторых птиц фронтальная косица в это время почти полностью отсутствует. Filoplumulae риктальной косицы начинают выпадать в конце августа; была поймана только одна неполовозрелая малая конюга, передняя часть косицы которой находилась в состоянии линьки. До этого времени у всех птиц данная косица была в хорошем состоянии. У всех пойманных птиц аурикулярная косица была в хорошем состоянии и можно предположить, что её линька происходит позже. Линька украшающих образований заканчивается, вероятно, в конце октября – начале ноября (рис. 10).
Линька рамфотеки . Сроки и последовательность линьки рамфотеки приводятся по опубликованным (Конюхов 2001) и неопубликованным данным автора, собранным на острове Булдырь в 1993 и 1999 годах.
Первые признаки смены рамфотеки у малых конюг появляются в начале июля. На Командорских островах смена щитков клюва начинается в середине-конце июля (Stejneger 1885). У птицы, добытой на острове Ионы 1 сентября 1971, клюв был красный с грязно-белой вершиной, без следов линьки роговых пластин (Нечаев, Тимофеева 1973). Сроки линь -ки рамфотеки у птиц с Курильских островов остаются неизвестными.
У малой конюги, как и у близкородственного ей вида — большой ко-нюги, происходит сезонная смена щитков рамфотеки. Первые её признаки, проявляющиеся в изменение окраски щитков, отмечены в середине июля. Старое роговое вещество щитков, окрашенное в красный цвет, замещается новым роговым веществом беловатого цвета, которое со временем постепенно приобретает сначала серый, а затем и чёрный цвет. Схематически направление и последовательность линьки представлена на рисунке 12.
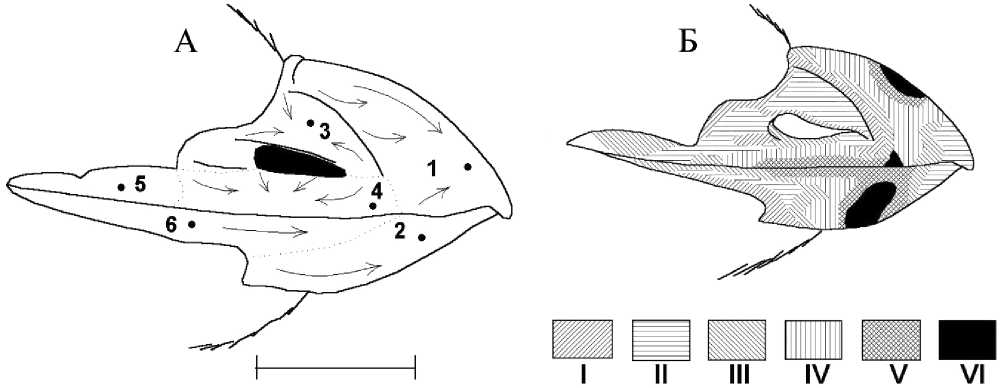
Рис. 12. Линька рамфотеки малой конюги.
А – направление линьки щитков (стрелками показано направление, в котором идёт смена щитка); Б – схема линьки (последовательность линьки от I к VI. Одинаковой штриховкой показаны участки рамфотеки, сменяющиеся примерно в одни и те же сроки; участки, обозначенные более плотной штриховкой, отпадают позже (масштабный отрезок равен 5 мм) (по: Конюхов 2001)
Щитки сменяются двумя способами одновременно. Во-первых, верхние слои рамфотеки постепенно снашиваются, а на смену им «поднимается» тёмное роговое вещество. При этом происходит постепенное изменение цвета данных участков щитка с ярко-красного через грязные оттенки красного цвета сначала на серый, а затем и чёрный цвет. Граница между старым и новым роговым веществом при этом размытая. Во-вторых, там, где слой старого рогового вещества на щитке особенно толст, его края при линьке обламываются. При таком способе смены щитка граница между старым и новым роговым веществом резкая.
Необходимо отметить, что толщина старого ярко окрашенного рогового вещества щитка больше, чем нового. Поэтому то место, где роговое вещество новое, обычно выглядит как углубление, а на границах щитков образуются щели, достигающие нового рогового вещества.
Кульменальный щиток, подобно рамфотеке надклювья всех птиц, сдвигается вперёд за счёт роста нового рогового вещества в его каудальной части, то есть у границы оперения. Начальные стадии его линьки зарегистрированы в начале июля; на границе щитка и оперения начинает формироваться новое роговое вещество. В конце июля у большинства птиц кульменальный щиток уже отделился узкой щелью от оперкулярного щитка; а у некоторых особей он начинает отделяться и от суб-нариального, но роговое вещество на «дне» образовавшейся щели остаётся ещё беловатым, что говорит о начальных стадиях этого процесса. В начале августа кульменальный щиток уже отделён щелью от оперкулярного и субнариального щитков, а к середине месяца эта щель между щитками доходит до режущего края надклювья.
Поскольку в описании окраски клюва малой конюги говорится, что «клюв кораллово-красный с белой вершиной...» (Головкин 1990), то считаю необходимым проследить изменение окраски этой вершины. С кон -ца июля её цвет постепенно изменяется с беловатого или желтовато-белого на роговой. Её потемнение начинается с участков, граничащих с красной рамфотекой, а светлой остаётся область, примыкающая к режущему краю и самому концу клюва. У единичных птиц, пойманных в конце августа, конец клюва был почти чёрным. Вполне вероятно, что зимой этот участок кульменального щитка имеет более светлую окраску, чем остальной щиток.
Оперкулярный щиток, сменяясь, постепенно истончается и одновременно обламывается с краёв. В начале июля его цвет варьирует у разных птиц от почти красного до серого со слабыми красноватыми оттенками. В начале третьей декады июля почти у всех птиц оперкулярный щиток уже становится чёрным, и только у 1 из 17 птиц, пойманных 28 июля, его оттенок был серовато-красным, то есть яркий слой старой рамфотеки полностью ещё не сошёл. По мере уменьшения размера этого щитка и «сползании» вперёд кульменального щитка между ними образуется щель, «дно» которой покрыто новой рамфотекой чёрного цвета. Эта щель разной ширины была отмечена у всех птиц. К началу августа замена этого щитка практически завершилась у всех птиц. С этого времени у всех птиц он тёмный (чёрный или темно-серый), таким образом, оперкулярный щиток сменяется одним из первых.
Верхний томиальный щиток, линяя, постепенно меняет свой цвет с беловатого на чёрный. Смена окраски происходит от его дорзального к вентральному (режущему) краю, так что наиболее тёмным является самый верхний край щитка. В третьей декаде июля этот щиток почти у всех птиц становится очень темно-серым, почти чёрным, а к началу августа та часть щитка, которая выступает из-под оперения, у всех птиц была чёрной. Впоследствии, до начала сентября (конца полевых работ), цвет этого щитка не изменялся.
Субнариальный щиток также сменяется двумя способами. Постепенный «сход» щитка идёт с его заднего края, от верхнего томиального щитка, где тёмное роговое вещество плавно переходит в старое красновато -оранжевое, тогда как у переднего края щитка эта граница между старой и новой рамфотекой очень контрастная. Основная поверхность щитка, видимо, заменяется постепенным снашиванием слоёв, так как наряду с птицами, у которых встречен яркий, почти красный щиток, встречались, хотя и редко, птицы с очень бледным светло-оранжевым щитком, который кажутся выцветшим.
Субнариальный щиток начинает сменяться в начале июля. К концу июля практически у всех птиц новое (чёрное) роговое вещество охватывает ноздрю спереди и сзади. В начале августа новое чёрное роговое вещество выстилает уже всю дорзальную часть щитка, соединяя два участка новой рамфотеки - за и перед ноздрёй. К концу первой декады августа у некоторых птиц этот щиток уже весь чёрный, но у большинства особей нижняя, режущая его часть всё-таки имеет в своей окраске какие-то тёплые тона.
В середине августа у разных птиц субнариальный щиток, находясь на разных стадиях линьки, окрашен очень разнообразно. Крайние варианты: щиток почти весь старый, ярко-красный с тёмной полоской новой рамфотеки по своему верхнему краю или весь новый, чёрный. Тот и другой варианты — исключения, встречающиеся редко. Более обычны промежуточные варианты: задняя и верхняя части щитка чёрные или темно-серые разной интенсивности. От этих участков новое роговое вещество отходит косыми полосами, направленными сверху вниз и сзади наперёд, и вдаётся в старое роговое вещество, которое остаётся на нижнем, режущем крае щитка. Различия заключаются в соотношении старого и нового рогового щитка на щитке. У части птиц поверхность щит -ка может быть однородного желтовато-серого или красновато-серого цвета, без какого-либо выделения по цвету режущего края.
Гониальный и нижний томиальный щитки не имеют ясной границы. Её можно заметить только при смене этих щитков. Сменяясь, старая рамфотека постепенно сдвигалась вперёд, а также снашивалась, постепенно истончаясь. В середине июля у некоторых птиц уже намечается граница между нижним томиальным и гониальным щитками в виде неясного сероватого углубления. К середине августа эта граница становится достаточно чёткой, особенно в своей передней части, где тёмное роговое вещество нижнего томиального щитка контрастирует с красным или беловатым роговым веществом гониального щитка.
У некоторых птиц смена гониального щитка закончилась к середине августа, однако у большинства этот щиток окрашен очень разнообразно. Ярко окрашенное, красное или розовое роговое вещество может покрывать большую или меньшую его поверхность, причём, как и в случае с кульменальным щитком, чем ярче эти участки, тем меньшую площадь они занимают.
Новое роговое вещество нижнего томиального щитка образуется в его задней и нижней части. В начале августа у небольшого числа птиц этот щиток уже почти весь новый, исключая режущую часть. Этот участок в виде полоски контрастирует с серым или чёрным роговым веществом на остальной поверхности щитка. Эта полоска, которая в задней части щитка узкая и сходящая на нет, а в переднем более-менее широкая, сохраняется у большинства птиц практически до конца августа. В середине августа у некоторых птиц этот щиток уже целиком тёмный.
Предбрачная линька
(Definitive Basic > Definitive Alternate)
При этой линьке затрагивается только контурное оперение. О её сроках точных данных нет, заканчивается она, вероятно, в первой половине мая. О последовательности линьки данных нет. Линька контурного оперения происходит с конца зимы и до начала сезона размножения; у птицы, добытой 6 мая на острове Беринга, свежие перья в пеньках были на брюшке, шее и голове (Stejneger 1885).
Украшающие образования, видимо, не претерпевают никаких изменений в течение всей зимы, поскольку их смена и рост заканчивается ещё осенью (см. выше). Данные о линьке рамфотеки очень скудны. У взрослых птиц в декабре-январе оперкулярный щиток и прилегающие к ноздре участки кульменального и субнариального щитков черноватокоричневые, далее кульменальный щиток тускло-красный, а на кончике рогового цвета, коричневато-белый. В последующие месяцы клюв приобретает яркую окраску: кончик становится почти чисто-белым, середина – алой, а оперкулярный щиток, перед самым началом сезона размножения окрашивается в нежный карминовый цвет (Stejneger 1885).
Питание
Морфологические приспособления. Модель челюстей малой конюги в натуральную величину, сделанная по результатам исследований биомеханических особенностей челюстного аппарата вида, показала, что минимальные объекты, которые могут ловить эти птицы, лежат в пределах 2-3 мм. Более мелкие объекты выталкиваются наружу выносящим потоком, который возникает при смыкании челюстей. Исходя из приведённых данных можно предположить, что конюги охотятся в толще воды, схватывая по одному рачку. По-видимому, во время движения птиц под водой они производят частые движения челюстями. В этом случае, особенно при большой плотности зоопланктона, могут захватываться объекты размерами 2-5 мм. Более крупные и подвижные рачки, вероятно, целенаправленно выбираются короткими клевками. Оптимальный выбор объектов (в среднем около 10 мм) удовлетворяет птиц с энергетической точки зрения и в соответствии с размерами клюва сводит на нет выталкивание рачков из ротовой полости с потоком воды (Соколов 1990).
Время кормёжки . Малые конюги в основном кормятся днём (Day, Byrd 1989), однако во время полнолуния они, вероятно, могут кормиться и ночью (Schacter 2017; Schacter, Jones 2018).
На Лисьих островах малые конюги собирались в стаи в местах стоячих волн и в районах сильных приливных течений как летом, так и в другие сезоны. Летние скопления порой были значительными; в середине июля к югу от острова Уналга и Бабьих островов встречались стаи, состоящие из более чем 1000 птиц. Д.Нисевандер с соавторами (Nyse-wander et al. 1982) также наблюдали большие скопления малых конюг, главным образом в проливе Аватанак; вблизи Бабьих островов их было меньше. Стаи конюг в период размножения к вечеру рассеиваются, и по крайней мере размножающиеся взрослые особи перемещались к гнездовьям на близлежащих островах (Bradstreet, Herter 1991).
В проливе Бабий в 1991 году тысячи малых конюг кормились в приливных волнах на плотных скоплениях планктона, которые наблюдались с помощью 120 кГц эхолота. Обычно стаи включали 10-50 особей, а крупные моновидовые скопления, состоящие из тысяч птиц, были отделены от других видов морских птиц (Hunt et al . 1993). Летом кормящиеся малые конюги наблюдались вблизи Бабьих островов, острова Эгг и прилегающих проливов. Небольшие стаи численностью до 200 особей в каждой в течение всего лета обычно встречались в проливах Седанка, Уналга, в устье Бобровой бухты и в проливах между островами Эгг и Уналга. В середине июля встречено несколько стай из более чем 1000 особей в каждой в 1 км к югу от Бабьих островов и у острова Уналга. Группы малых конюг чаще всего были связаны с приливными волнами и другими областями сильных течений. 14 июля ближе к вечеру учтено около 11400 малых конюг, стаи которых летели непрерывным потоком в течение почти часа с запада на восток вдоль южного берега острова Уналга от мест кормёжки в проливе Уналга (Bradstreet, Herter 1991).
На Четырёхсопочных островах (пролив Кагамил, восточные Алеуты) 2 июня 1978 наблюдалось скопление примерно из 10 тыс. малых конюг, кормившихся в стоячих волнах высотой приблизительно 2 м. На следующий день тысячи кормящихся особей отмечены в зоне приливных возмущений пролива между островами Амукта и Чагулак. В центральной части Алеутских островов, в районе острова Угидак (острова Деларова), 30-31 июля 1977 наблюдались около 200 особей, кормившихся в приливных волнах. На западных Алеутских островах у юго-западной оконечности острова Кыска (Крысьи острова) 10 августа 1978 держалось скопление примерно из 750 кормящихся малых конюг в зоне приливного фронта (Day, Byrd 1989).
В период размножения на Курилах малые конюги летают кормиться в расположенные недалеко от колоний проливы, где добывают пищу среди сулое в* , образующихся на фронтах приливно-отливных течений. Значительные кормовые концентрации птиц наблюдаются в проливах Экарма, северной части Крузенштерна, Головнина и северной части пролива Надежды (Артюхин 2009, 2016).
Кормовой разлёт и длительность кормовых полётов. Почти все малые конюги, которых видели во время обследований Алеутских островов, обычно кормились в пределах 9-18 км от берега (Byrd, Gibson 1980; Troy 1991). На Курильских островах кормовые разлёты значительнее, чем на Алеутских. Птицы с островов Чёрные Братья летают кормиться за 30 км к северо-восточной оконечности Урупа, а птицы с острова Рай-коке кормятся в 50 км у островов Ловушки (Артюхин 2003; Артюхин и др. 2001). Малые конюги, гнездящиеся на острове Чиринкотан, по-ви-димому, летают кормиться на мелководные участки ближайших проливов, где их часто регистрировали во время морских учётов (Артюхин и др. 2001). На Командорах 9 августа 1993 малые конюги кормились поодиночке и стаями до 30 особей в 2-2.5 км от северо-восточного побережья острова Беринга. Плотность птиц на этом участке акватории составляла 141.7 ос./км2 (Артюхин 1998). На острове Ионы одиночные птицы кормились в 50-500 м от острова. Судя по частоте прилётов и внушительному размеру шейного мешка, малые конюги не испытывали недостатка в корме (Андреев и др. 2012).
Способ лова добычи . Прямых наблюдений нет, но исходя из биомеханики челюстного аппарата можно предположить, что кормящаяся малая конюга схватывает по одному рачку за один раз (см. «Морфологические приспособления»).
Рацион. Основные пищевые объекты . Летом малые конюги, по всей видимости, становятся почти монофагами, питаясь копеподами Neoca-lanus , когда последние в изобилии встречаются у поверхности океана, но зимой, вероятно, переключаются на поедание амфипод и других зо-опланктонных ракообразных, когда Neocalanus мигрируют на глубину 600-1000 м для размножения (Day, Byrd 1989). Летом рацион малых ко-нюг более разнообразен, чем у других конюг Aethia , то есть этот вид более гибок в использовании других ресурсов, когда их основная добыча становится дефицитной. Такая гибкость в выборе объектов питания позволяет малой конюге шире, чем конюге-крошке и большой конюге, заселять Алеутский архипелаг (Williams et al . 2003).
Малые конюги, добытые в акватории острова Булдырь 26 мая – 4 августа 1976, питались по меньшей мере 10 видами планктона, среди которых наиболее важный компонент – зоопланктонные ракообразные, за которыми в порядке убывания следовали щетинкочелюстные (хетог-наты), моллюски и личинки рыб. В конце мая (конец откладки яиц и начало насиживания) в питании малых конюг 98% составлял Neocala-nus plumchrus , из которых определённых до вида – 62.0% от общего IRI, остальное – фрагменты копепод, вероятно, того же N. plumchrus . Около
2% пищи приходилось на неопознанных хетогнат и фрагменты ракообразных. В начале июля (конец насиживания и начало выкармливания птенцов) рацион малых конюг стал разнообразнее; на Parathemisto pacifica приходись 33.8%, Neocalanus cristatus – 30.1%, а Neocalanus plum-chrus – 25.2% от общего количества IR I* . Оставшиеся 11% были фрагментами копепод, неопознанными амфиподами и их фрагментами, неопознанными хетогнатами, фрагментами ракообразных и неопознанными тканями животных. Однако это разнообразие, скорее всего, результат крайне малой выборки. В середине июля (середина периода выкармливания птенцов) малые конюги опять поедают в основном N. plumchrus (88.1% от общего числа IRI). На фрагменты копепод, вероятно, того же вида приходилось 8.7%, а неопознанные эвфаузииды, мегалопы Erima-crus isenbeckii , фрагменты ракообразных и неопознанные птероподы (вероятно, Limacina helicina) , головоногие моллюски и хетогнаты – 3.2% пищи. В начале августа (конец периода выкармливания птенцов) малые конюги стали практически монофагами: 99.8% приходилось на копепод, среди которых 90.4% от общего количества IRI составляли целые N. plumchrus и ещё 9.3% их фрагменты. N. cristatus присутствовал в следовых количествах. Оставшиеся 0.2% пищи состояли из неопознанных хетогнатов, личинок рыб, фрагментов ракообразных и тканей животных (Day, Byrd 1989).
В центре своего изобилия малые конюги выбирают в качестве основной добычи эвфаузиид, одна из которых является глубоководным видом, а не копепод. В районе пролива Бабий в 1991 году, видимо летом, у 16 добытых особей основной пищей была эвфаузиида Thysanoessa inermis (Hunt et al . 1993).
Остаётся невыясненным вопрос о базовых компонентах рациона малых конюг, поскольку в разных районах основу питания составляют то копеподы, то эвфаузииды. Вероятно, птицы добывают те организмы, которые в данный момент наиболее многочисленны и соответствуют размерным ограничениям, определяемым биомеханическими особенностя -ми их челюстного аппарата.
Конкурентные отношения с другими видами
На местах гнездования . На острове Булдырь в голых осыпях, а также осыпях, находящихся на разных стадиях зарастания травой, малые конюги гнездятся вместе с большими конюгами и конюгами-крошками, а на заросших травой прибрежных склонах - с белобрюшками, качурками и топорками Fratercula cirrhata (Зубакин, Конюхов 1999). На острове Матуа малые конюги гнездятся в колониях совместно с качурками, на острове Ушишир — с большими конюгами (Гизенко 1955).
При совместном гнездовании в осыпях малые конюги всегда вытесняются из гнездовых камер большими конюгами ( n = 38), тогда как сами только в 70% случаев ( n = 25) вытесняют конюг-крошек (Knudson, Byrd 1982). Отмечены случаи, когда насиживающая малая конюга была вытеснена белобрюшкой, отложившей в гнездо своё яйцо (Hipfner, Byrd 1993). В 1999 году одно из гнёзд малой конюги с находившимся там однодневным птенцом было разрушено, а птенец убит рывшим нору топорком (Н.Б.Конюхов, неопубл. данные).
На местах кормёжки . Хотя зимой в проливе Унимак существуют смешенные кормовые скопления больших и малых конюг, чаще встречаются моновидовые (Troy, Bradstreet 1991). Однако тут сказывается, скорее всего, не конкуренция между этими видами, а предпочтение каждого добывать пищу в предпочтительных кормовых биотопах.
За объекты питания. Незначительные различия в размерах конюг- крошек и малых конюг должны привести к значительному перекрытию наборов пищевых объектов (Bedard 1969). Это предположение подтвердилось на Булдыре, где гнездятся все 5 видов конюг: малая, большая, американская, конюга-крошка и белобрюшка. Межвидовые различия в количестве пойманной добычи были вызваны различиями в размерах кормовых объектов, а не в их доступности. Копеподы Neocalanus были основным кормовым объектом для всех конюг. Малая конюга, как и ко-нюга-крошка, ловили в основном более мелких N. plumchrus (~5 мм в длину), доля которых в рационе составляла 90.4 и 86.6% (IRI), соответственно. Большая конюга и белобрюшка специализировались на более крупных N. cristatus (~10 мм в длину) и Parathemisto pacifica (~10 мм в длину); белобрюшки тоже ловили мегалоп Erimacrus isenbeckii, длина которых тоже ~10 мм (Day, Byrd 1989). Ж.Бедар (Bedard 1969) предположил, что эта конкуренция привела к малочисленности малых конюг на Алеутских островах, где конюга-крошка многочисленна, и к обилию малых конюг на Курильских островах, где конюг-крошек нет.
Паразиты
Паразиты взрослых птиц . У малых конюг, добытых на острове Бул-дырь, найдены цестоды Alcataenia pygmaeus и Diorchis pelagicus . Первый вид, видимо, имеет ограниченный ареал и паразитирует в слизистой оболочке двенадцатиперстной кишки только малой конюги, второй паразитирует также и в большой конюге (Hoberg 1982, 1984, 1986). Це-стода Alcataenia larina также паразитирует в малых конюгах. Промежуточные хозяева этих цестод – эвфаузииды Euphausiidae, являющиеся важным компонентом рациона малой конюги. Кроме того, паразитами малых конюг является нематода Stegophorus stellae-polaris и пероеды Saemundssonia wumisuzume (Philopteridae) (Price et al . 2003; Muzaffar, Jones 2004).
Источник цитрусового запаха у конюг в настоящее время неизвестен; в неволе такого запаха у птиц нет. Альдегиды или их предшественники могут быть выделены из естественной пищи конюг, которая состоит в основном из зоопланктона (Jones 1993). Анализ экстрактов из перьев с затылка малой конюги выявил присутствие там гептилового (22.3%), пеларгонового (22.3%), децилового (9.7%) альдегидов и цетилового спирта (45.6%). Как показали эксперименты, компоненты этого одоранта малой конюги отпугивают эктопаразитов, а их смеси могут существенно превосходить эффект от воздействия этих веществ в отдельности. Содержащиеся в оперении эти репелленты препятствуют паразитированию клещей и вшей. Исследования показывают, что альдегидные одоранты больших и малых конюг видоспецифичны и указывают на эволюционное расхождение. Оба вида в своём одоранте содержат дециловый альдегид. Однако экстракты перьев малых конюг отличаются от таковых большой конюги тем, что у них преобладают альдегиды с нечётные числом углеродных атомов, отсутствуют ненасыщенные альдегиды и есть цетиловый спирт. Всё это указывает на видоспецифичные пути биосинтеза или поглощения одорантов конюгами Aethia (Douglas III. et al. 2004).
Паразиты птенцов . Данных нет.
Болезни / Заморы . Данных нет.
Х и щ н и к и
Аборигенные . На Командорских и Алеутских островах на малую ко-нюгу охотятся болотная сова Asio flammeus , сапсан Falco peregrinus и белоголовый орлан Haliaeetus leucocephalus . Основной враг птенцов на Алеутах – серокрылая чайка Larus glaucescens (Knudtson, Byrd 1982; Артюхин 1998, 1999; Hunter et al . 2002). Сапсан, белоголовый орлан и серокрылая чайка – основные враги взрослых конюг на Булдыре. Из всех останков конюг Aethia , найденных в погадках белоголовых орланов ( n = 33) и сапсанов ( n = 61), только по 3% в каждом случае принадлежали малым конюгам (Knudtson, Byrd 1982). Даже когда в 1991 и 1992 годах сапсан гнездился непосредственно над плотной колонией малых конюг на Крестед Поинт, только 11% ( n = 54) и 8% ( n = 53) останков Aethia принадлежали малым конюгам (Hunter et al . 2002).
Беринговский Vulpes lagopus beringensis и медновский V. l. semenovi песцы являются коренными обитателями Командорских островов (Ог-нёв 1931), а рыжие лисицы Vulpes vulpes – для восточных Алеутских островах, на запад по крайней мере до острова Умнак, самого западного из Лисьих островов (Murie 1959). По этой причине эти острова и получили такое название. Вполне вероятно, что природная популяция лисиц обитала и на острове Чугинадак, крупнейшем из Четырёхсопочных островов (Bailey 1993). Остатки малых конюг в небольшом числе неоднократно встречались в норах песцов на островах Медный (Шиенок 2022) и Беринга (Пилипенко 2025).
На Курильских островах аборигенные популяции полёвки-экономки Microtus oeconomus населяли острова Алаид, Парамушир, Шумшу и Онекотан. В настоящее время к ним добавились острова Шиашкотан, Ушишир, Матуа, Кетой и Симушир. Её появление на этих островах связано с искусственным расселением с целью создания кормовой базы для разводимых там лисиц и песцов. Полёвку-экономку стали расселять с 1916 года, но точная дата выпуска и количество выпущенных особей неизвестны. Для этой интродукции донорскими были популяции островов Парамушир и Шумшу. Полёвка-экономка не является прямой угрозой для малой конюги, но поддерживает популяцию песца зимой, когда птицы на островах немногочисленны (Воронов 1982).
Интродуцированные. Малые конюги используют самые разнообразные места гнездования и исторически гнездились с относительно низ- кой плотностью, вероятно, почти на каждом острове Алеутского архипелага и большинстве Курильских островов. Завезённые наземные хищники существенно повлияли на распространение и численность малых конюг в силу их уникальной особенности спать на островах. Как взрослые, так и неполовозрелые птицы, а также слётки в течение круглого года возвращаются ночью на сушу. Эта особенность делает малых конюг значительно более уязвимыми к хищничеству по сравнению с другими представителями рода Aethia.
Песцы и рыжие лисицы – основные интродуцированные хищники на Алеутских и Курильских островах (Воронов 1982; Bailey 1993). Материнскими популяциями для заселения этих островов послужили песцы в основном с Командор, а также островов Прибылова. Серые крысы Rat-tus norvegicus были случайно завезены на 16-18 островов Алеутской гряды (Bailey 1993; Ebbert, Byrd 2002), а домовая мышь Mus musculus – по крайней мере на 3 алеутских острова (Bailey 1993).
Массовая интродукция песцов на Алеуты началась с острова Атту в 1750 году, хотя на основании коммерческих записей купцов Олаус Дж. Мьюри (Murie 1959) высказывает предположение, что песец на Атту мог быть и аборигенным видом, однако археологические исследования на островах Атту, Агатту не обнаружили костей наземных млекопитающих в мусорных ямах коренных жителей (Bailey 1993). Позднее песцы постоянно завозились на новые острова на протяжении 1800-х годов. Заселение песцами и лисицами новых островов архипелага достигло пика в период с 1913 по 1940 год, когда почти на каждом острове обитали завезённые песцы или лисы (Bailey 1993; Williams et al . 2003). Первым видом, интродуцированным на Курильские острова с Командорских островов в 1916 году, был песец. В 1916-1917 годах с целью создания оптимальной кормовой базы для песца и лисицы была интродуцирована и полёвка-экономка (Воронов 1982).
Истреблять песцов на Алеутах с помощью ловушек и яда начали в 1949 году на острове Амчитка, и эта деятельность медленно, но неуклонно продолжалась до 1970-х годов, а позже наземных хищников стали уничтожать примерно ежегодно на одном острове. На ряде небольших островов, где песцы или лисы полностью истребили птиц, они сами вымерли естественным образом. К 2002 году Аляскинский морской заказник очистил от песцов и лис 40 островов, что позволило населению птиц постепенно восстанавливаться до состояния, существовавшего до интродукции псовых. Песцы сохранились лишь на островах Шемья, Танага, Канага, Адак, Атка, а лисицы – на острове Чугинадак (Bailey 1993; Eb-bert 2000; Ebbert, Byrd 2002).
На острове Амчитка до интродукции песцов в 1921 году гнездились малые конюги (Funk 2018). На острове Крысий до того, как туда примерно в 1780 году после крушения японского корабля попали крысы, тоже гнездились малые конюги (Bailey 1993; Croll et al. 2016). Вполне возможно, что малые конюги продолжали гнездиться в крайне ограниченном числе в недоступных для крыс щелях скальных стенок острова (Salmon, Ellen 2010). Песцов на этот остров завезли в 1922 году, а в 1984 году их там уничтожили (Bailey 1993; Williams et al. 2003; Salmon, Ellen 2010). Присутствие крыс на островах, с которых удалены песцы и лисы, может препятствовать восстановлению распространения и численности малых конюг (Williams et al. 2003), поскольку, как это отмечено для больших конюг и конюг-крошек (I.Jones, pers.comm.), крысы могут убивать и малых конюг, находящихся в гнездовых камерах. На таких островах, где остались только серые крысы, их численность начинает расти, поскольку сдерживающие её ранее песцы и лисицы уничтожены, а человеку уничтожить крыс крайне трудно.
В октябре 2008 года с вертолётов на острове Крысий, площадь которого составляет 2800 га, было сброшено 46 тонн приманки, содержащей родентицид бродифакум. В 2009 и 2010 годах проводили обследования острова на наличие грызунов, и 30 августа 2010 официально объявили о полном уничтожении популяции крыс на острове (Salmon, Ellen 2010).
Домовая мышь обнаружена на Симушире и Итурупе, но она тесно связано с населёнными пунктами. К счастью, сейчас на Курильских островах серая крыса не заселила небольшие необитаемые острова – Кетой, Ушишир, Среднева, Ловушки, Чёрные Братья и Птичьи (Воронов 1982), на которых находятся основные колонии морских птиц, в том числе и малой конюги.
Способы хищничества. Сапсаны ловят конюг в воздухе. Серокрылые чайки подстерегают в колонии слетающих птиц и преследуют их, стре- мясь поймать в воздухе. Если конюги садятся на воду, то чайки преследуя их, заставляют непрерывно нырять. Если конюги не уплывают под водой от чаек так далеко, что те их не смогут заметить, то чайки в конце концов их ловят. Песцы, лисицы ловят птиц в колониях, находящихся в осыпях; серые крысы поедают яйца или птенцов, а также взрослых птиц в гнёздах.
Песцы довольно быстро сокращают число гнездящихся на островах птиц: на острове Янкича в 1959 году в их питании первое место занимали глупыши Fulmarus glacialis , затем шли воробьиные и чистиковые, а в 1970-х годах, после того, как численность глупышей сократилась до тысячи особей и остались только те птицы, которые гнездились в недоступных местах, на первое место в рационе хищника вышли массовые здесь виды – малая и большая конюги. В 1971 году на островах Ушишир насчитывалось не менее 5 тыс. песцов (Воронов 1982).
Ответ на хищничество. Малые конюги больше других видов конюг страдают из-за хищничества интродуцированных рыжих лисиц и песцов (Williams et al. 2003), но их пластичность в выборе биотопов гнездования позволяют им продолжать гнездиться (хотя и не в таком большом числе) на тех островах, куда эти хищники были завезены. Обследование, охватившее острова почти четверти Алеутской гряды от острова Ка-гамил на востоке до острова Касаточий на западе, было осуществлено в 1982 году и выявило влияние хищников на популяции морских птиц. Несмотря на то, что остров Амукта был одним из рекордсменов по добыче песцов на Алеутах, там в трещинах лавы, застывшей вдоль побережья, в 1982 году продолжали гнездиться малые конюги, а на острове Касаточий при высокой численности песцов в осыпях у подножья кальдеры всё ещё продолжали гнездиться большие и малые конюги, численность которых составляла 20 и 15 тыс. особей соответственно. Песцы, выпущенные на остров Кагамил в 1922 году, в 1982 году оставались довольно многочисленными, и все морские птицы, включая малых конюг, гнездились исключительно на скалах. Несмотря на то, что лисицы, завезённые ещё русскими в XIX веке, всё ещё обитали на острове Чугина-дак в 1982 году, малые конюги продолжали гнездиться там в щелях скальных стенок и в значительном числе в лавовых потоках на южном побережье острова. На островах Сигуам, Херберт и Уляга до заселения их песцами находились большие колонии больших конюг, белобрюшек и топорков, соответственно, но к 1982 году их там не стало, хотя малые конюги продолжали гнездиться (Bailey, Trapp 1986). На острове Уналга лисы были интродуцированы в 1842 году, а песцы – в 1914 году. Здесь малые конюги гнездятся в щелях скал на высоте, недоступной для наземных хищников (Oehler 2008). Песцы ввезены на остров Юнаска в 1919 году, однако в 1982 году малые конюги были наиболее многочисленными среди морских птиц. Они гнездились в трещинах застывших лавовых потоков вдоль значительной части побережья. В июне ~2500 малых конюг встречено в приливных возмущениях у юго-западной оконечности острова; сходное число птиц было зарегистрировано в том же районе десятью годами ранее (Bailey, Trapp 1986), что говорит о том, что эти птицы гнездятся на этом острове.
Влияние человека
Хозяйственного значения малая конюга не имеет.
Сбор яиц . Специального сбора яиц этого вида, по-видимому, никогда не производилось.
Охота . Сейчас малая конюга охотничьего значения не имеет. В прежние времена добывалась алеутами. Судя по отложениям в мусорных ямах, в небольшом числе добывалась на острове Крысий (Croll et al . 2016). На острове Булдырь в мусорных отложениях одной ямы среди чистиковых наиболее многочисленной была малая конюга, на долю которой приходилось 28% от числа обнаруженных там птиц, в другой яме – 6%. Скорее всего, добывалась малая конюга в основном не для еды, поскольку её вес лишь немного превышает 100 г, а из-за их украшающих перьев, которые использовались для украшения одежды (Lefevre, Siegel-Causey 1993; Lefevre et al . 1997).
Загрязнение . Исследования, проведённые на Аляске в 1969-1977 и 1988-1990 годах, а также на Командорских островах в 1988-1994 годах, показали, что малые конюги не заглатывают кусочки пластика (Day 1980; Robards et al . 1995; Robards et al . 1997; Артюхин 2014).
Пролив Унимак – оживлённое место, через который проходит большое количество грузовых, рыболовецких, военных и связанные с нефтяной промышленностью судов. Здесь малая конюга подвергается огромному риску воздействия нефти и нефтепродуктов в случае их разлива, поскольку в проливе и сопредельной акватории в течение круглого года держится значительная часть её популяции. Поскольку малые конюги встречаются в море плотными скоплениями, то попадание их в нефтяное пятно приведёт к гибели большого числа птиц (Troy 1991).
Коммерческое рыболовство. В снасти при коммерческом лове рыбы в прибрежных водах Курильского архипелага малые конюги не попадают (Артюхин 2014), однако они подвержены фатальному ночному влечению к искусственному освещению на судах (Byrd, Williams 1993). Так, в начале ноября 1964 года на судно, стоявшее на якоре с подветренной стороны Четырёхсопочных островов и прятавшееся там от шторма (скорость ветра достигала 50 м/с), с 23 до 6 ч прилетело и разбилось 1140 малых конюг. Однако в июле 1946 года на борту судна, стоявшего у тех же островов, была найдена только одна разбившаяся малая конюга, привлечённая огнями судна (Dick, Donaldson 1978). В первом случае массовая гибель птиц произошла, видимо, из-за того, что конюги, как и судно, прятались от шторма за островами и там собрались в большом количестве. На Курилах в проливе между островами Матуа и Топорковый 25 сентября 1947 в туманную ночь на палубу шхуны за 2 ч приземлилось, было отловлено и выпущено больше 40 особей (Гизенко 1955).
Абиотические факторы
На острове Касаточий 7-8 августа 2008 произошло извержение вулкана. Исследования, проведённые на острове летом 2009 года, показали, что все бывшие места размножения морских птиц были погребены тридцатиметровым слоем тефры. Не было никаких визуальных свидетельств того, что птицы копали норы или откладывали яйца. Однако автоматический фиксатор звуковой активности (SM -1), установленный в бывшей гнездовой колонии Troll Talus, зафиксировал ночное присутствие птиц на суше. Голоса малых конюг регистрировались до 29 июля, с наибольшей активностью в конце каждой ночи (04:00-04:15). Утром 12 июня голоса сотен малых конюг доносились с моря, но при дневном обследовании акватории ни одной птицы не встречено (Williams et al . 2010).
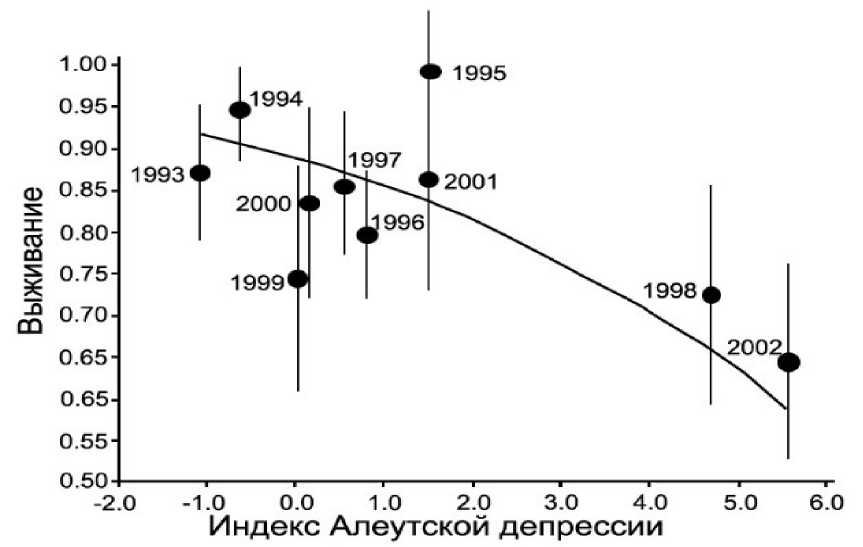
Рис. 13. Корреляция между ежегодной выживаемостью взрослых малых конюг (± SE) на острове Булдырь в 1993-2002 годах и индексом Алеутской депрессии (Jones et al . 2007)
Выживаемость, продолжительность жизни
Выживаемость взрослых малых конюг в течение первого года после мечения составила 0.71±0.04% (SE), в то время как после этого — в среднем 0.84±0.03%, что позволяет предположить, что около 13% вновь меченых особей в любой год были «не местными», то есть не гнездились на том участке побережья, на котором проходили исследования. Средний показатель выживаемости малых конюг был немного меньше (но статистически не отличался), чем был получен для конюги-крошки – 0.87± 0.04% (SE) (Jones et al. 2002) и большой конюги – 0.86±0.02% (SE) (Jones et al. 2004) за тот же промежуток времени (Jones et al. 2007). Годовалые птицы не размножались и, скорее всего, не были постоянно привязаны к какому-то конкретному месту в колонии, поэтому их оценка выживаемости была ниже.
Выживаемость малых конюг на острове Булдырь была самой высокой в годы со слегка отрицательным или нейтральным значением индекса Алеутской депрессии (слабая алеутская аномалия низкого давления), а самой низкой в годы с положительным индексом Алеутской депрессии (относительно сильная или интенсивная алеутская аномалия низкого давления) (рис. 13). Также выживаемость положительно коррелировала с температурой поверхности моря в августе - апреле близ острова, то есть более высокая ежегодная выживаемость следует за периодами повышенной температуры воды (Jones et al . 2007).
Доля неполовозрелых особей сильно варьировала в разные годы, но она не коррелировала с продуктивностью предыдущего года, позволяя предположить, что пополнение колоний зависит не от высокой успешности размножения, а от какого-то фактора, определяющего выживаемость сеголетков в зимнее время (Jones et al . 2007).
Продолжительность жизни . При отловах паутинными сетями пойманы 2 малых конюги, помеченные 10 лет назад взрослыми (Jones et al . 2007).
Сохранение и охрана
Заповедывание . Вместе с другими природными объектами малая ко-нюга охраняется на Командорских островах в национальном парке «Командорские острова» имени С.В.Маракова и на Алеутских островах в Аляскинском морском национальном природном заповеднике (Alaska Maritime National Wildlife Refuge).
Восстановление популяций . Мероприятия по удалению песцов и лисиц с островов, куда те были завезены людьми, должно привести к расширению (или восстановлению былого) ареала и численности.
После того, как на Алеутах стали уничтожать интродуцированных туда лисиц и песцов, численность малых конюг стала увеличиваться, а их распространение расширяться. К 2003 году малых конюг во всё большем числе стали наблюдать на острове Агатту и близлежащих островах. Новые колонии отмечены на островах Кыска, Канага, Улак, Касаточий, Конюжий и Амля, где песцов теперь нет. Как и прежде, много малых конюг встречено в Бабьем проливе, вблизи Четырехсопочных островов, островов Юнаска и Большой Ситкин. У острова Сигуам, где песцы были уничтожены в 1996 году, замечена стая малых конюг численностью 30-
40 тыс. особей, то есть больше, чем В.Бёд и Д.Гибсон (1980) оценивали численность этих птиц на всех Алеутских островах (Williams et al . 2003).
Поскольку через 5 лет после уничтожения крыс на острове Крысий стали гнездиться северные качурки и топорки (Croll et al . 2016), то можно предположить, что и численность малых конюг увеличится, и они расселятся по всему острову; «затравкой» для этого могут послужить малые конюги, гнездящиеся на островке у мыса Аюгадак, где они продолжали жить, когда остров Крысий населяли наземные хищники (Salmon, Ellen 2010).
Время, в течение которого произойдёт восстановление конкретной популяции, остаётся неизвестным. Так в 1916 году остров Конюжий был сдан в аренду жителям острова Атка для ведения пушного хозяйства, но были ли песцы туда завезены – точно не известно. О том, что они туда всё же попали, свидетельствуют незанятые в 1982 году травянистые склоны, представляющие собой отличное место для гнездования птиц-норников, а на скалистых участках побережья птицы гнездились излишне скученно, как на Касаточьем острове, где песцы были многочисленными. Возможно, песцы исчезли с Конюжьего острова несколько десятилетий назад, но территория, доступная для гнездования птиц, до сих пор полностью повторно не заселена (Bailey, Trapp 1986).
Я искренне благодарен сотрудникам Аляскинского морского национального природного заказника (Alaska Maritime National Wildlife Refuge) за предоставленную возможность работы на острове Булдырь, а также за техническую поддержку исследований. Я крайне признателен Ю.Б.Артюхину, В.А.Зубакину, К.Е.Литвину и С.П.Харитонову за замечания и ценные советы во время написания данной публикации.
