Птицы России и сопредельных стран: малая конюга Aethia pygmaea (Gmelin, 1789). 3. Размножение
. 3. Размножение")
Автор: Н.Б. Конюхов
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 2578 т.34, 2025 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140313030
IDR: 140313030
Birds of Russia and neighboring countries: the whiskered auklet Aethia pygmaea (Gmelin, 1789). 3. Breeding biology
Текст статьи Птицы России и сопредельных стран: малая конюга Aethia pygmaea (Gmelin, 1789). 3. Размножение
Поступила в редакцию 29 октября 202 5*
Предгнездовой период
Прибытие птиц в окрестную акваторию . На Алеутских и, возможно, на Курильских островах малые конюги присутствуют в колонии круглый год. Там, где прилегающая к колониям акватория замерзает, птицы, видимо, появляются сразу же после разрушения сплошного ледового покрова, но конкретных данных нет.
Микробиотоп. По всему ареалу малые конюги занимают пять типов биотопов (Конюхов 2025б). На острове Булдырь они предпочитают каменные субстраты (χ2, P <0.01), в то время как большие конюги Aethia cristatella и конюги-крошки Aethia pusilla не проявляют существенного предпочтения ни к одному из субстратов (Knudson, Byrd 1982). В верхней части каменистых пляжей, выше полосы прибоя в самый высокий прилив, птицы заселяют как небольшие (7.5-12 м2) плоские каменные россыпи – «грядки», имеющие 20-30°наклон к морю и обычно являющиеся продолжением береговых осыпей или разрушающихся прибрежных каменных стенок, так и группы валунов. Они соответственно состоят из камней диаметром 30-50 см и 1-1.5 м. Гнездовые камеры на пляже располагаются под первым или вторым слоем камней таких «грядок» или в пустотах под крупными валунами (Зубакин, Конюхов 1999). Даже в таких биотопах, как заросшие травой склоны, где есть значительный слой почвы, скальные породы (выходы скал, камни разного размера) являются обязательной частью гнездовых камер; в 94.4% случаях (n = 54) они формировали какую-то часть гнездовой камеры и только в трёх случаях (5.6%) гнездовые камеры не были связаны со скалами и их можно назвать норами, которые первоначально могли быть вырыты либо вилохвостой Oceanodroma furcata, либо северной O. leucorhoa качурками. В двух случаях вилохвостых качурок находили в заброшенных гнездовых камерах конюг. На Основной осыпи малые конюги предпочитали гнез- довые камеры с негустой травой вокруг их входов; растительности не было в 45.3% случаев (n = 54), короткая трава – в 37.7% случаев и высокая трава – только в 17% случаев (Hunter et al. 2002).
На острове Эксельсиор, самом большом из Бабьих островов, большинство щелей, в которых гнездились малые конюги, находилось на высоте от 4 до 20 м над уровнем моря (Oehler 2008).
На Командорах малые конюги занимают расщелины скал и пустоты каменных осыпей на скалистых участках побережья (Артюхин 2016).
На Курилах в колониях на скалах Лев и Морская Выдра (острова Чёрные Братья) и на островах Ловушки малые конюги чаще всего гнездятся в вертикальных трещинах шириной 5-15 см. Гнездовые камеры находятся в 30-100 см от начала трещин. Обычно гнёзда располагаются на высоте 10-15 м, а в отдельных случаях до 50 м над морем (Велижанин 1977). На острове Райкоке гнезда находили в каменисто-шлаковых осыпях в нижних частях склонов острова (Трухин 2003).
Выход птиц на колонию . Инструментальные исследования, проведённые на Булдыре, доказали, что малые конюги присутствуют на острове круглый год (Schacter 2017; Schacter, Jones 2018), и возможно, то же самое имеет место на всех Алеутских островах. При визуальном обследовании акватории острова 30 апреля 1974 наблюдались тысячи ко-нюг в море в пределах 5 км от острова, но ни одной птицы не было замечено на Основной осыпи, где эти конюги гнездятся. К 15 мая в 19741976 годах большие, малые конюги и конюги-крошки присутствовали на Основной осып и* , когда та в основном была свободна от снега (Knudtson, Byrd 1982). Однако в этих исследованиях не было ночных обследований гнездовых биотопов малых конюг, где та уже должна была присутствовать (см. ниже).
На Командорских островах малые конюги появляются на своих гнездовых участках в первых числах мая (Stejneger 1885; Бианки 1909). В тех частях ареала вида, где прилегающая к островам акватория замерзает, птицы начинают выходить на сушу, видимо, сразу же после прилёта, но точных данных нет.
Неразмножающиеся птицы в колонии. Время прилёта неполовозрелых (sad) и неразмножающихся взрослых (ad) птиц в колонию остаётся неизвестным. Принимая во внимание то, что слётки малой конюги начинают возвращаться на сушу вскоре после слёта, то неполовозрелые особи должны присутствовать в колонии всё то время, в которое присутствуют и размножающиеся птицы. Неполовозрелые малые конюги в небольшом числе отмечаются в течение всего периода размножения и в послегнездовой период. Их численность в течение сезона размножения оставалась примерно на одном уровне, не увеличиваясь в период уси- ления акустической активности малых конюг в конце июля (Зубакин, Конюхов 1999). В зависимости от типа биотопа доля неразмножающих-ся птиц во время сезона размножения на острове Булдырь варьирует от 14.3 до 61.1% (Конюхов Н.Б., неопубл. данные).
На Булдыре в 1992-2003 годах доля пойманных неполовозрелых (в возрасте 1 года) птиц резко различалась по годам и была статистически значимой; χ 2 = 78.8, df = 10, P <0.001; в разные годы она составляла от менее 5% до 31% птиц.
Гнездовой период
Возраст первого размножения . Поскольку молодые птицы сразу же после слёта начинают посещать колонию и принимать участие в акустической активности колонии, то это создаёт предпосылки для более раннего начала размножения по сравнению с другими видами конюг. Вероятно, малая конюга способна приступать к размножению уже в 2летнем возрасте (Конюхов и др. 2000). Позже эта гипотеза получила своё подтверждение. Среди малых конюг, отловленных на острове Булдырь с 15 мая по 15 июня, полностью васкуляризированные наседные пятна наблюдались у 3.5% годовалых птиц, 52.4% 2-летних, 94.0% 3-летних, 97.0% 4-летних и у 100% особей более старшего возраста. Из 1730 пойманных птиц 820 (47%) были размножающимися, 297 (17% от общего числа, или 27% от обследованных) – неразмножающимися, у 524 (30%) наседные пятна не обследовались, а 89 (5%) особей были отловлены после 15 июня (Jones et al . 2007).
Соотношение полов . Если вид моногамен, то соотношение полов в популяции должно быть близким к 1:1. Исследований по этому поводу немногочисленны и результаты их неоднозначны. Среди 102 взрослых малых конюг, пойманных и генетически разделённых по полу, 40 (39%) были самками, а 62 (61%) – самцами, то есть в выборке взрослой популяции преобладали самцы (Binomial, Z = 2.2, P = 0.03). В выборке неполовозрелых (sad) птиц ( n = 49) 20 (41%) были самками и 29 (59%) – самцами, что соответствует аналогичному соотношению, но существенно не отличается от соотношения 1:1 в группе неполовозрелых (sad) птиц. Соотношение самцов и самок в выборках только взрослых птиц и только неполовозрелых особей различалось незначимо (Binomial, Z = 1.3, P = 0.2). В то же время оба эти соотношения также не отличались от гипотетического стандартного соотношения полов 1:1 ( χ 2 = 0.04, df = l, P = 0.86) При объединении всех малых конюг, пол которых был определён, 60 (40%) птиц были определены как самки и 91 (60%) как самцы, что указывает на значительное преобладание самцов в общей выборке (Binomial, Z = 2.5, P = 0.01). Из 134 птенцов малых конюг 59 (44%) были самцами, а 75 (56%) – самками (Binomial, Z = 1.4, P = 0.2), то есть без значимого отклонения от 1:1 в этом возрасте (Jones et al . 2007).

Рис. 1. Малые конюги Aethia pygmaea . Остров Янкича, Курильские острова.
14 июня 2009. Фото Ю.Б.Артюхина
Ухаживание и выбор партнёра . Во время ухаживания на воде у всех четырёх видов конюг Aethia можно наблюдать три вида демонстраций – сгорбленную позу, преследование и взаимное ухаживание, но покачивание головой (ритмичное движение головой вперёд-назад как во время ходьбы, head-bob) было отмечено только у малой конюги. Сгорбленная поза встречалась почти исключительно как часть демонстрации преследования (плаванье в сгорбленной позе – 2.3%, 1/43; преследование – 20.9%, 9/43). Из всех наблюдавшихся пар 43 малых конюг 67.4% (29/43) демонстрировали покачивание головой (head-bobbing display). Взаимное ухаживание проявлялось в том, что оба члена пары поворачивались головой друг к другу и вокализировали. Если взять все проявления вместе, то 4 вида конюг различались по частоте ухаживания ( χ 2 = 55.0, P < 0.001), причём малые конюги демонстрировали самую высокую частоту ухаживания (95.3%, 41/43 из наблюдаемых пар, участвующих в ухаживании) (Hunter, Jones 1999).
Система брачных отношений . Малая конюга социально моногамна (Gaston, Jones 1998).
Место и время спаривания. Из-за ночного образа жизни практически ничего не известно о брачном поведении малых конюг на суше. Малые конюги, как и все представители подсемейства Fraterculinae, спариваются в море (Конюхов 1996; Hunter, Jones 1999; неопубл. данные автора) в утренний период активности сразу после слёта на воду рядом с гнездовой колонией (Hunter, Jones 1999). Однако однозначно говорить, что это единственный вариант места и времени спаривания, мы не можем, поскольку та часть жизни, которую птицы проводят в море вдали от берега, остаётся практически неизвестной. Вполне вероятно, что ко-нюги могут спариваться и в течение всего дня, находясь вне поля зрения исследователей. Маловероятно, что они также спаривались в своих гнездовых камерах, поскольку большинство гнездовых камер слишком малы, чтобы в них поместилась спаривающаяся пара. Высота гнездовой камеры составляет всего лишь 85.2% от высоты спаривающейся пары, равной примерно 140 мм (Hunter, Jones 1999).
Паттерн копуляции у всех четырёх видов конюг одинаков. Самец делает садку и частыми взмахами крыльев поднимает тело вертикально, придавая лапами себе устойчивость, заводит свою клоаку под хвост самки и осуществляет клоакальный контакт. Вероятно, взмахи крыльев, которыми самец поддерживает себя в вертикальном положении, также приподнимают и самку, благодаря чему её клоака находится выше поверхности воды, что предотвращает контакт спермы с водой (Hunter, Jons 1999).
Неудачи при спаривании могут происходить по ряду причин: неспособности самца достичь или поддерживать правильное (вертикальное) положение при спаривании, уклонении самки от копуляции и вмешательства сторонней особи. Для успешности спаривания необходимо содействие самки. Ей надо удерживать правильное положение на воде и держать хвост вертикально вверх для осуществления клоакального контакта. Самки могут как избегать ухаживания самца, так и прерывать попытки спаривания: они либо улетают, либо уплывают, либо ныряют, или просто прижимают хвост к поверхности воды или погружают его в воду. От действия или бездействия самок происходит 18.8% случаев неудач от всех попыток спаривания (Hunter, Jones 1999).
На всех этапах ухаживания и спаривания пару могут беспокоить конспецифичные особи. Мешать могут как особи, подплывающие к ухаживающей паре, так и птицы, налетающие и сбивающие самца непосредственно перед или во время клоакального контакта. Пару во время ухаживания беспокоят в 10% процентах случаев, во время спаривания – 4.5%. После «сбоя» нормального течения ухаживания или спаривания один или оба партнёра отгоняют «незваного гостя» или одна птица из пары оттесняет другую от «незваного гостя» (Hunter, Jones 1999).
Внебрачные спаривания. Добровольные внебрачные спаривания отмечались, но насильственных копуляций из-за специфики спаривания быть не может. Внебрачные спаривания происходили следующим образом. Самец быстро подплывал или подлетал к паре и, оказавшись рядом с ней, пытался спариться с самкой. Самец из пары обычно нахо- дился поблизости и в большинстве случаев пытался помешать чужому самцу спариться со своей самкой и либо прогонял его, либо отгонял свою самку от этого самца. В каждом случае после попытки внебрачного спаривания самка остаётся со своим «законным» самцом. Самцы из пары часто пытаются спариваться со своей самкой сразу после попытки внебрачного спаривания (Hunter, Jones 1999).
Социальное поведение . На острове Булдырь с ночной активностью малых конюг в колонии разного рода социальные взаимодействия происходят под камнями или в пустотах скальных стенок. Здесьпроисходят агрессивные столкновения между соседями или между хозяевами гнездовой камеры и пришлыми птицами. Об этом можно судить по вспышкам вокализации нескольких птиц в одной точке колонии; нередко после этого из ближайшей к месту конфликта щели на поверхность выскакивает конюга, которая затем улетает или уходит в другую щель (Зубакин, Конюхов 1999).

Рис. 2. Малые конюги Aethia pygmaea . Остров Ионы, Охотское море. 16 июля 2001. Фото Ю.Б.Артюхина
На острове Ионы, где активность малых конюг не отличается от активности больших конюг и конюг-крошек, социальное взаимодействия между птицами происходят утром и вечером в «клубах», которые обычно образуются на пологих камнях, где пары токуют, сблизив клювы. В середине дня много малых конюг сидит на отвесных скалах у моря, цепляясь за малейшие неровности поверхности (Харитонов 1980).

Рис. 3. Малые конюги Aethia pygmaea . Остров Янкича, Курильские острова.
14 июня 2009. Фото Ю.Б.Артюхина
Социальная активность в стаях на воде . В конце утреннего периода активности, после слёта на воду, малые конюги на 60-75 мин объединяются в плотные стаи примерно в 200-500 м от берега. Птицы в таких стаях держатся в 15-30 см друг от друга. Они быстро и хаотично двигаются и постоянно вокализируют, издавая только трельки, но не мяукающие крики. Все это указывает на какие-то формы активного социального взаимодействия между птицами в стае. По плотности и конфигурации (округлые, овальные, реже в виде ленты) стаи малых конюг очень напоминают аналогичные стаи больших конюг, также образующиеся в нескольких сотнях метров от берега в периоды вечерней и утренней активности. Стаи включают от нескольких десятков до нескольких сотен малых конюг. Они практически всегда моновидовые, очень редко к ним может присоединяться небольшое число больших конюг. Численность птиц в стаях постоянно изменяется, так как стаи то разделяются, то вновь соединяются. Нередко все или большая часть слетевших утром особей колонии объединяются в единую стаю. В стаях малых конюг время от времени можно видеть коллективные купания, когда одновременно начинает купаться большая часть птиц, и синхронные ныряния всех или почти всех птиц стаи – аналогично тому, что часто происходит в подобных стаях больших конюг (Зубакин, Конюхов 1999).
Утренние стаи малых конюг отмечаются в течение всего июня и первой половины июля; в конце июля они формировались не ежедневно, а в августе такие стаи вблизи берега перестают образовываться вовсе. Не исключено, однако, что птицы собираются в стаи на значительном расстоянии от берега, вне пределов видимости и слышимости наблюдателей (Зубакин, Конюхов 1999).
Структура гнездовой колонии
Пространственная структура колонии . Для поселений малых конюг характерно группирование гнездящихся птиц в субколонии. Наиболее чётко субколонии выявляются на осыпях, обрывах и пляжах, менее – на прибрежных склонах, заросших высокой и густой травой. Величина центральных субколоний составляет десятки пар, периферических – от 2-3 до 7-9 пар. Расстояние между субколониями на Булдыре варьирует от 2 до 13 м. Субколонии объединяются в колонии, которые, как правило, находятся на расстоянии нескольких сотен метров друг от друга. В центральных частях плотных колоний субколонии обычно очерчиваются границами пригодных для гнездования участков биотопов (осыпи, каменные стенки, россыпи камней подходящего размера и т.д.). На периферии таких колоний группирование гнездящихся пар имеет явно социальные причины, поскольку подходящих мест для гнездования здесь больше, чем используется малыми конюгами. Одиночного гнездования на расстоянии сотни и более метров от ближайшего соседа не отмечено ни разу. Крайне редким было и гнездование на расстоянии более 10 м от ближайшего соседа (Зубакин, Конюхов 1999).

Рис. 4. Малые конюги Aethia pygmaea . Остров Ионы, Охотское море. 16 июля 2001. Фото Ю.Б.Артюхина
Социальная структура колонии . Целостность колонии в Алеутской части популяции с ночной активностью особей в колонии поддерживается благодаря вокализации (Зубакин, Конюхов 1999). В западной половине ареала, где активность малых конюг дневная, подобных исследований не проводились.
Гнездовая территория . Гнездовой консерватизм у малой конюги достаточно высок. В 66.7% случаях (28 из 42) пары отмечались в прошлогодней гнездовой камере, в 23.8% – один из партнёров был новым, и в 4 случаях (9.5%) гнездовую камеру заселила новая пара. Две пары гнездились ежегодно в течение 5 лет в одной и той же гнездовой камере, ещё две – 4 года, и ещё три были найдены и в 1994, и в 1998 году и, вероятно, гнездились или пытались гнездиться там и в промежуточные годы. Один партнёр из пары 1996 года в 1997 году отсутствовал, а на его месте была другая птица, но в 1998 году прежняя пара восстановилась. В 3 случаях птицы сменили гнездовые камеры; в одном из них пара разделилась: каждая птица на следующий год образовала новую пару, причём одна из птиц осталась в старом гнезде, а другая загнездилась в новом. В 2 других случаях только один из партнёров образовал новую пару в другой гнездовой камере, второй не был обнаружен, а старая камера осталась пустой (Hunter et al . 2002).
Владелец гнездовой территории и гнезда . Данных нет, но возможно, что, как и у большой конюги – это самец (Зубакин 1990; Зубакин, Зубакина 1993; Клёнова, Зубакин 2013).
Выбор места для гнезда и плотность гнездования . Точных данных о выборе места для гнезда нет. В период выкармливания и слёта птенцов на участке скальной стенки неоднократно наблюдались птицы, переходившие от одной трещины или щели к другой и надолго исчезавшие в них. Возможно, это были неразмножавшиеся особи, подыскивающие место для будущего гнездования (Зубакин, Конюхов 1999). Однако остаётся неясным, были ли это птицы, собирающиеся на следующий год здесь загнездиться, либо просто подыскивающие место для ночлега.
Плотность гнездования в целом зависит от и частоты встречаемости гнездовых камер в данном биотопе. На острове Булдырь наиболее разрежено конюги гнездятся на заросших травой склонах, где расстояние между входами ближайших соседних гнездовых камер в среднем составляло 5.9±0.71 м (SE, n = 31; 1.1-17.0 м; медиана 5.5 м). На скальных обрывах птицы могут гнездиться как отдельными парами на расстоянии нескольких метров друг от друга, так и с большей плотностью. Участок каменной стенки площадью 9 м2, изобилующий трещинами и пустотами, населяло 7-9 пар малых конюг. С наибольшей плотностью конюги заселяют каменистые пляжи; минимальное расстояние между гнездовыми камерами здесь в среднем составляет 0.72±0.03 м (SE, n = 33; 0.51.2 м; медиана 0.7 м) (Зубакин, Конюхов 1999). На острове Ионы плот- ность населения малой конюги на глыбовых полях и травянистых склонах составляет в среднем 0.26±0.13 особи на 1 м2 (SE, n = 3; lim 0.130.39 м), на скальных стенках в среднем 0.1±0.04 ос./м2 (SE, n = 2; lim 0.070.13 м). Средняя плотность для всех биотопов оценивается в 0.20±0.13 особи на 1 м2 (SE, n = 5) (Андреев и др. 2012).
Гнездо
Расстояние гнезда от моря . На большей части ареала малые конюги гнездятся в непосредственной близости от моря, на склонах, обращённых в сторону моря. Расстояние от гнездовых камер до уреза воды составляет от нескольких до сотен метров. На острове Конюжий птицы гнездятся не только среди валунов и на скалистых участках по периметру острова, но и во внутренних частях острова, поднимаясь до его вершины в 275 м (Bailey, Trapp 1986).
Гнездовые камеры . Малые конюги гнёзд не строят, а занимают естественные убежища и, как исключение, норы качурок. Траву в клюве никогда не носят. В гнёздах на травянистых склонах часто бывают растительные остатки, но они, видимо, заносятся туда пассивно либо самими птицами, либо ветром. Малые конюги, гнездившиеся в осыпи на острове Булдырь, откладывали яйца на ровную поверхность, без углублений и «подстилки» (Knudson, Byrd 1982). В 7.4% ( n = 54) гнездовых камер, находящихся в нижней части Основной осыпи, примыкающей к берегу, дно гнездовой камеры устилали мелкие камешки, в 92.6% случаях оно было земляным. Количество входов в гнездовую камеру во многом зависит от биотопа гнездования. На почвенных склонах, заросших травой, он всегда один. На осыпях и на берегу их может быть несколько. Так, на Основной осыпи один вход был у 69.1% ( n = 55) гнездовых камер, два – 23.6%, три – 5.5% и четыре – 1.8% (Hunter et al . 2002).
На Командорских островах яйца откладываются в большинстве случаев прямо на голый камень, реже – на дресву или почву, попавшую в расщелины (Велижанин 1977).
Поскольку для гнездования используются естественные укрытия, то размеры входа в гнездовую камеру характеризуют её недоступность для основных конкурентов – больших конюг и белобрюшек Aethia psittacula . Иногда гнездовая камера так узка и низка, что в неё может втиснуться только одна насиживающая птица. Однако длина этой расщелины может составлять десятки сантиметров. Нередко гнездовая камера имеет изгибы. Ширина входа гнездовых камер на Булдыре, которые удалось измерить, составляет 54.4±2.7 мм (SE , n = 33), а высота – 79.8±4.4 мм (SE , n = 33), средняя глубина (расстояние от входа до яйца) – 284±112 мм (SD , n = 22, lim 120-546 мм) (Byrd, Williams 1993; Hipfner, Byrd 1993). На Основной осыпи медиана высоты основного входа в гнездовую камеру составляла 61 мм ( n = 52, lim 32-179 мм), медиана ширины – 64.5 мм ( n =
38, lim 39-406 мм), медиана высоты потолка в камере – 108 мм ( n = 54, lim 60-237 мм). В 75.9% случаев ( n = 54) гнездовые камеры были круглыми, а в 24.1% – овальными/удлинёнными (Hunter et al . 2002). Средняя высота гнездовой камеры 112.7±36.3 мм (SD , n = 54, lim 60-237 мм) (Hunter, Jones 1999). Средний объём гнездовой камеры – 162.5±70.3 см3 (SD , n = 12), что значимо меньше, чем у большой конюги и значимо не отличается (P >0.01) от такового конюги-крошки (Knudson, Byrd 1982).
Температура в гнездовых камерах ( n = 7) на Булдыре в 1992 году была на 1° ниже, чем температура воздуха снаружи (Byrd, Williams 1993).
Чистки и углубления гнездовых камер не отмечено. Яйцо как правило откладывается в тот же «лоток», что и в прошлом году. Нередко на дне гнездовой камеры присутствуют скорлупа или яичные оболочки, оставшиеся от предыдущего сезона размножения.
Относительная чистота гнездовых камер в период инкубации говорит о том, что насиживающие птицы здесь не испражняются; не исключено, однако, что птицы используют под «уборные» дальние отнорки. Специальных выходов из гнездовой камеры для дефекации насиживающими малыми конюгами, как это наблюдается, например, у больших конюг на острове Талан (Зубакин 1990), не отмечено ни разу (Конюхов и др. 2000). Птенцы в гнездовой камере испражняются в строго определённых местах, обычно либо у выхода, либо в отнорках или в дальних углах гнездовой камеры, на что указывают скопление помёта и относительная чистота самом гнезде.
Гнездовые камеры не для размножения . Птицы периодически посещают некоторые камеры, на что указывают перья, скапливающиеся в них. Среди этих камер есть как вполне подходящие для гнездования, так и непригодные для этого, к которым относятся укрытия недостаточной глубины или с широким входом. Птицы в них ночуют.
Откладка яиц и их характеристики
Характеристики яйца . Скорлупа яйца гладкая, без блеска, при откладке матово-белая, затем во время насиживания пачкается почвой лотка, приобретая иногда коричневый цвет (Конюхов и др. 2000).
Данные о размерах и массе яиц представлены в таблице 1. Сведения о массе свежеотложенных яиц крайне ограничены. Вес единственного свежего брошенного яйца, найденного 24 июня 1993 в гнездовой камере на острове Булдырь, составила 23.5 г (Конюхов и др. 2000). Средний вес яиц, собранных на островах Чёрные Братья в период с 31 мая по 5 июня 1974, составил 25.8±0.4 г (SD; n = 20; lim 21.5-29.4 г) (Велижанин 1977). На острове Ионы вес яйца, найденного в момент проклёва, был 25.4 г при размерах 48.2×32.7 мм (Андреев и др. 2012).
На острове Булдырь длина яйца в разные годы (1995-1998) не различалась, но ширина значимо варьировала по годам (ANOVA: длина:
F = 1.77, df = 2 и 63, P = 0.2; ширина: F = 3.75, df = 2 и 63, P = 0.03), а критерий Шеффе (Scheffe F -test) показал, что при уровне значимости 0.05 яйца в 1995 и 1998 годах не отличались от яиц двух других лет.
Таблица 1. Размеры яиц малой конюги (среднее ± SD *, в скобках n , lim)
|
Длина, мм |
Ширина, мм |
Год |
Источник |
|
Остров Булдырь |
|||
|
44.0±1.5*, (26), 41.0-46.3 |
31.2±1.0*, (26), 29.2-32.8 |
1993 |
Byrd, Williams 1993 |
|
43.8±1.9*, (20), 39.0-46.3 |
31.6±0.97*, (20), 28.7-32.7 |
1993, 1999 |
Конюхов и др. 2000; Конюхов, неопубл. данные |
|
45.1±08*, (7), 44.0-46.4 |
31.9±0.6*, (7), 31.1-32.8 |
1995 |
Hunter et al. 2002 |
|
43.7 (1) |
29.3 (1) |
1996 |
Hunter et al. 2002 |
|
43.9±1.7*, (27), 40.5-46.8 |
30.8±1.2*, (27), 28.8-32.2 |
1997 |
Hunter et al. 2002 |
|
144.0±1.5*, (30), 40.6-46.8 |
31.3±1.1*, (30), 28.7-33.3 |
1998 |
Hunter et al. 2002 |
|
44.1±1.56*, (65), 40.5-46.8 |
31.1±1.14*, (65), 28.7-33.3 |
1995-1998 |
Hunter et al. 2002 |
|
44.0±1.5*, (26), 41.0-46.3 |
31.2±1.0*, (26), 29.2-32.8 |
– |
Gaston Jones 1998 |
|
Курильские острова |
|||
|
46.0–48.5 |
31.5-32.8 |
– |
Гизенко 1955 |
|
46.6±0.4*, (20), 42.8-50.2 |
32.5±0.2*, (20), 30.5-34.8 |
31.05-5.06 1974 |
Велижанин 1977 |
|
45.2-48.5 |
31.5-33.5 |
– |
Головкин1990 |
|
45.2-48.0 |
32.0-33.5 |
– |
Козлова 1957 |
На острове Булдырь индекс объёма яйца, вычисленный по формуле: 0.512 × (длина яйца × 0.1) × (ширина яйца × 0.1)2, в 1995 году составил 23.5±1.0 мм3 (SD , n = 7, lim 22.2-25.1 мм3), в 1996 – 19.2 мм3 ( n = 1), 1997 – 21.3±2.2 мм3 (SD , n = 27, lim 17.9-25.71 мм3), 1988 – 22.1± 2.0 мм3 (SD , n = 7, lim 17.8-25.91 мм3); средний индекс объёма за 1995-1998 годы составил 21.9±2.10 мм3 (SD , n = 65, lim 17.8-25.91 мм3). В 1995, 1997 и 1998 годах объём яиц значимо различался между годами (ANOVA: F = 3.54, df = 2 и 63, P = 0.04). Критерий Шеффе показал, что при P = 0.05 яйца 1995 года были значительно крупнее яиц 1997 года, а яйца 1998 года были среднего объёма и не отличались от яиц двух других лет (Hunter et al . 2002).
Величина кладки . Кладка всегда состоит из одного яйца. Повторных кладок взамен утраченных не отмечено.
Откладка яиц . Скорее всего, яйца откладываются ночью, после возвращения птиц в колонию после кормёжки.
Исследования 1988-1992 годов на острове Булдырь показали, что откладка яиц у малых конюг происходит в период с 12 мая по 5 июня (Williams et al . 1994). Если продолжительность насиживания у этого вида составляет 35-36 сут ( n = 3) (Knudson, Byrd 1982), то начало откладки яиц на Булдыре в 1993 году, судя по датам вылупления первых птенцов, пришлось на конец апреля – первые числа мая (Конюхов и др. 2000); там же в 1993-1998 годах откладка яиц пришлась на 4 мая – 20
июня (Hunter et al . 2002), на Командорских островах – на начало мая (Stejneger 1885), на острове Ионы – на конец мая – начало июня (Харитонов 1980; Андреев и др. 2012).На Командорах первые кладки отмечали в начале июня (Иогансен 1934). На Курильских островах откладка яиц, вероятно, происходит позже, чем на Алеутах. На острове Брат Чир-поев 31 мая и 3 июня найдены, соответственно, 5 и 6 яиц; свежими из них были 4 и 2 яйца, а в остальных у эмбрионов начала закладываться кровеносная система. На скале Морская Выдра 5 июня собраны 7 яиц, из них 1 свежее и 2 с эмбрионами на стадии закладки головы (Велижа-нин 1977). Из-за ранних сроков начала размножения прямых данных об откладке яиц у малых конюг нет. Судя по тому, что вылупление птенцов происходит асинхронно, то и откладка яиц должна происходить в колониях асинхронно.
Относительно факторов, влияющих на откладку яиц, есть только косвенные данные. Многоснежная зима и холодная весна 1999 года на острове Булдырь сдвинули начало сезона размножения, что привело к более поздним срокам слёта птенцов (неопубл. данные автора).
Насиживание
В насиживании кладки принимают участие оба партнёра. У обоих развивается по два наседных пятна (рис. 5) (Конюхов и др. 2000). Их максимальные размеры 33.1±3.3 мм (SD , n = 10, lim 29-39 мм) × 23.8±2.7 мм (SD , n = 10, lim 18-28 мм) (Byrd, Williams 1993; Gaston, Jones 1998).
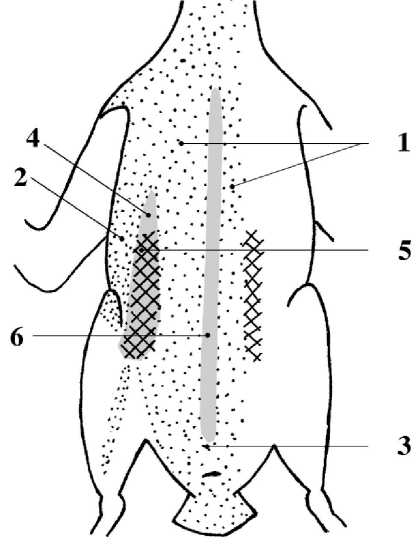
Рис. 5. Расположение наседных пятен (заштрихованы) у малой конюги.
Птерилии: 1 – грудинная Pt. steralis, 2 – грудная Pt. pectoralis, 3 – брюшная Pt. abdominalis; аптерии: 4 – срединная Apt. pectorale medianum, 5 – боковая туловищная Apt. truncale laterale, 6 – грудинная Apt. sternale. Названия птерилий и аптерий даны по: Кашенцева 1988 (по: Конюхов 1996)
Наседные пятна у малой конюги, как и у других видов подсемейства Fraterculinae, расположены только на боковых туловищных аптериях (apt. truncale laterale) и формируются за счёт выпадения пуховидных перьев. Контурные перья с грудинной (pt. sternalis) и грудной (pt. pec-toralis) птерилий не выпадают (Конюхов 1996; Конюхов и др. 2000).
Время начала образования наседных пятен остаётся неизвестным, поскольку сезон размножения у малых конюг начинается раньше, чем начинались исследования. Если время их формирования относительно откладки яйца сходно с таковым у большой конюги, то наседные пятна начинают закладываться в конце апреля – первой половине мая.
Зарастать наседные пятна начинают после вылупления птенцов и полностью покрываются перьями, так что к концу августа размножавшиеся особи становятся неотличимыми от не размножавшихся.
Поведение родителей во время насиживания . Обязанности по насиживанию партнёры делят поровну. Продолжительность смены составляет в среднем около 24 ч (по наблюдениям за 6 парами). Мониторинг одной пары в течение 12-дневного периода на острове Булдырь в 1992 году показал, что партнёры регулярно чередовались каждые 24 ч, за исключением одного случая, когда один из них насиживал яйцо в течение 48 ч (Byrd, Williams 1993). Смена партнёров на яйце происходит главным образом в начале ночи, вскоре после возвращения в колонию кормившихся птиц. Однако в ряде случаев прилетевший для смены партнёр может несколько часов провести вне своей гнездовой камеры, забираясь в неё только под утро. Освободившейся партнёр слетает либо вечером сразу после смены, либо утром; в последнем случае остаётся неизвестным, какой из партнёров насиживает яйцо ночью, когда они оба находятся в гнездовой камере (Конюхов и др. 2000).
Иногда родители могут оставлять яйцо (даже наклюнутое) без присмотра в течение дня; в последующем из таких яиц благополучно вылупляются птенцы (Конюхов и др. 2000).
Точных данных о начале насиживания нет, но, вероятнее всего, яйцо начинает насиживаться сразу же после его откладки. О продолжительности периода насиживания данные крайне ограничены, поскольку ко-нюги откладывают яйца до того, как до колоний добираются исследователи. Е.Кнудтсон и В.Бёд (1982) оценивают его в 35-36 сут ( n = 3).
Успешность инкубации . На Булдыре в 1976 и 1988-2024 годах под наблюдением находилось 2224 яиц малой конюги (рис. 6). Средняя успешность вылупления от числа отложенных яиц составляла 82.2±8.0% (SD , n = 36 лет, lim 100-25%) (Williams et al . 1997, 1998; Koike et al . 2025). В другом исследовании на этом же острове в период с 1988 по 1998 год вылупилось 84.4% ( n = 11 лет, lim 100-69%) яиц; успешность вылупления существенно не различалась в разные годы ( χ 2 = 17.4, df = 10, P = 0.1) (Hunter et al . 2002).
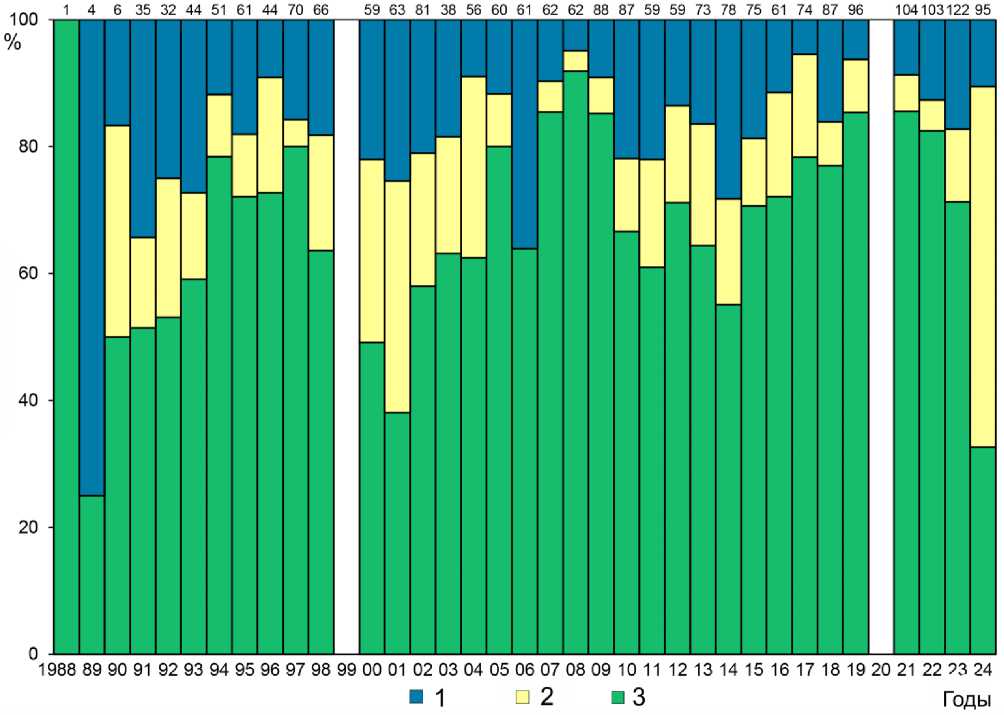
Рис. 6. Репродуктивные показатели малых конюг на острове Булдырь.
1 – гибель яиц, 2 – гибель птенцов, 3 – успешность размножения.
Гибель яиц = (А–Б)/А; гибель птенцов = (Б–С)/B; успешность размножения = С/А, где А – число отложенных яиц, Б – число вылупившихся птенцов, С – число слетевших птенцов. Объём выборки – числа над столбцами (по: Koike et al . 2025)
Причины гибели яиц . На Булдыре в 1976 и 1988-1997 годах птенцы не вылупились из 69 яиц (14.8%): 35 яиц исчезли из гнездовой камеры, 26 были брошены родителями, а 8 – разбиты (Williams et al . 1998). На острове Касаточий в 2007 году из 6 яиц, находившихся под наблюдением, 2 были выброшены из гнездовой камеры (Drummond, Larned 2007), а в 2008 году 1 из 5 яиц исчезло (Buchheit, Ford 2008).
Птенцы
Вылупление . Календарные сроки вылупления птенцов у малой ко-нюги представлены в таблице 2, а отклонение дат от многолетней средней показаны на рисунке 7. Фенология размножения на Булдыре заметно варьировала по годам: средние даты вылупления различались на 11 дней между сезонами (ANOVA, F = 17.57, df = 4 и 182, P = 0.0001). Согласно критерию Шеффе ( P = 0.05), в 1994 году вылупление происходило значительно раньше, чем в 1995 и 1997 годах, и в 1995 году значительно позже, чем во все остальные годы. Даты вылупления у птенцов, успешно покинувших гнездо, и у тех, которые не слетели, существенно не различались ( t = 0.84, n = 103 и 12, P = 0.4) (Hunter et al . 2002).
Таблица 2. Сроки вылупления птенцов малой конюги (среднее ± SD *, в скобках n , lim)
|
Дата |
Медиана |
Год |
Источник |
|
Остров Касаточий |
|||
|
(4), 16-22.06 |
– |
2008 |
Buchheit, Ford 2008 |
|
(4), 12-29.06 |
– |
2007 |
Drummond, Larned 2007 |
|
Остров Булдырь |
|||
|
30.06±4.6*, (6), 27.06-6.07 |
27.06 |
1976 |
Williams et al . 1998 |
|
24.06±5.4*, (5), 10.06-30.06 |
20.06 |
1990 |
Williams et al . 1998; Bond et al . 2011 |
|
27.06±3.6*, (9), 18.06-08.07 |
26.06 |
1991 |
Williams et al . 1998; Bond et al . 2011 |
|
18.06±10.7*, (5), 10.06-05.07 |
14.06 |
1992 |
Williams et al . 1998; Bond et al . 2011 |
|
22.06±8.2*, (13), 13.06-09.07 |
19.06 |
1993 |
Williams et al . 1998; Bond et al . 2011 |
|
19.06±7.6*, (37), 09.06-08.07 |
17.06 |
1994 |
Williams et al . 1998; Bond et al . 2011 |
|
25.06±6.3*, (50), 15.06-25.07 |
21.06 |
1995 |
Williams et al . 1998; Bond et al . 2011 |
|
22.06±9.8*, (27), 10.06-20.07 |
19.06 |
1996 |
Williams et al . 1997, 1998; Bond et al . 2011 |
|
24.06±7.9*, (33), 11.06-18.07 |
21.06 |
1997 |
Williams et al . 1998; Bond et al . 2011 |
|
23.06±9.8*, (61), 09.06-11.07 |
19.06 |
1998 |
Williams et al . 1998; Bond et al . 2011 |
|
19.06±6.1*, (25), 11.06-04.07 |
17.06 |
1993 |
Конюхов и др., 2000 |
|
16.06±6.4*, (28), 08.06-02.07 |
– |
1994 |
Hunter et al . 2002 |
|
27.06±6.5*, (29), 20.06–-.07 |
– |
1995 |
Hunter et al . 2002 |
|
21.06±8.4*, (32), 10.06-13.07 |
– |
1996 |
Hunter et al . 2002 |
|
20.06±4.4*, (46), 14.06-05.07 |
– |
1997 |
Hunter et al . 2002 |
|
18.06±4.2*, (48), 10.06-29.06 |
– |
1998 |
Hunter et al . 2002 |
|
20.06±6.8*, (183), 08.06-15.07 |
– |
1994-98 |
Hunter et al . 2002 |
|
17.06±5.2*, (27), 06.06-13.07 |
– |
2000 |
Bond et al . 2011; Warzybok 2011 |
|
22.06±6.2-, (36), 09.06-15.07 |
– |
2001 |
Bond et al . 2011; Warzybok 2011 |
|
24.06±8.5*, (36), 15.06-15.07 |
– |
2002 |
Bond et al . 2011; Warzybok 2011 |
|
25.06±2.9*, (4), 09.06-04.07 |
– |
2003 |
Bond et al . 2011; Warzybok 2011 |
|
21.06±4.7*, (28), 16.06-12.07 |
– |
2004 |
Bond et al . 2011; Warzybok 2011 |
|
18.06±6.0*, (29), 09.05-05.07 |
– |
2005 |
Bond et al . 2011; Warzybok 2011 |
|
27.06±4.7-, (20), 19.06-05.07 |
– |
2006 |
Bond et al . 2011; Warzybok 2011 |
|
17.06±7.8*, (24), 09.06-10.07 |
– |
2007 |
Bond et al . 2011; Warzybok 2011 |
|
21.06±3.2*, (40), 14.06-30.06 |
– |
2008 |
Bond et al . 2011; Warzybok 2011 |
|
17.06±6.7*, (62), 05.06-21.07 |
– |
2009 |
Warzybok 2011 |
|
23.06±5.0*, (24), 17.06-05.07 |
– |
2010 |
Warzybok 2011 |
|
19.06±7.5*, (28), 07.06-05.07 |
– |
2011 |
Warzybok 2011 |
|
20.06±9.6*, (18), 09.06-12.07 |
2012 |
Koike et al . 2025 |
|
|
21.06±10.2*, (30), 09.06-13.07 |
2013 |
Koike et al . 2025 |
|
|
16.06±6.0*, (29), 10.06-11.07 |
2014 |
Koike et al . 2025 |
|
|
16.06±5.7*, (37), 07.06-01.07 |
2015 |
Koike et al . 2025 |
|
|
16.06±9.7*, (21), 07.06-10.07 |
2016 |
Koike et al . 2025 |
|
|
12.06±4.9*, (23), 04.06-21.07 |
2017 |
Koike et al . 2025 |
|
|
22.06±5.3*, (34), 13.06-06.07 |
2018 |
Koike et al . 2025 |
|
|
18.06±8.6*, (50), 07.06-13.07 |
2019 |
Koike et al . 2025 |
|
|
20.06±7.5*, (54), 09.06-15.07 |
2021 |
Koike et al . 2025 |
|
|
14.06±5.4*, (52), 06.06-03.07 |
2022 |
Koike et al . 2025 |
|
|
18.06±4,5*, (46), 11.06-03.07 |
2023 |
Koike et al . 2025 |
|
|
01.07±4.8*, (93), 26.06-22.07 |
2024 |
Koike et al . 2025 |
|
|
Остров Ионы |
|||
|
~15.07 |
– |
1974 |
Харитонов 1980 |
Сроки вылупления в 1993 году несколько различались в разных биотопах: на Основной осыпи средняя дата вылупления — 18 июня ± 1.22 сут (SE , n = 16), а на травянистом склоне — 21 июня ± 2.6 сут (SE , n = 9).
Однако эти различия оказались статистически незначимыми ( t = 1.16, P = 0.26, df = 23) (Конюхов и др. 2000). Птенцы на острове Егг в 2005 году начали вылупляться примерно в середине июня, поскольку первая птица с кормом была поймана 18 июня (Jones, Seneviratne 2005). Размножение малых конюг на Андреяновских островах, вероятно, происходит в те же сроки, что и на острове Булдырь; 12 июня 1879 на острове Атка добыт пуховой птенец (Turner 1886).
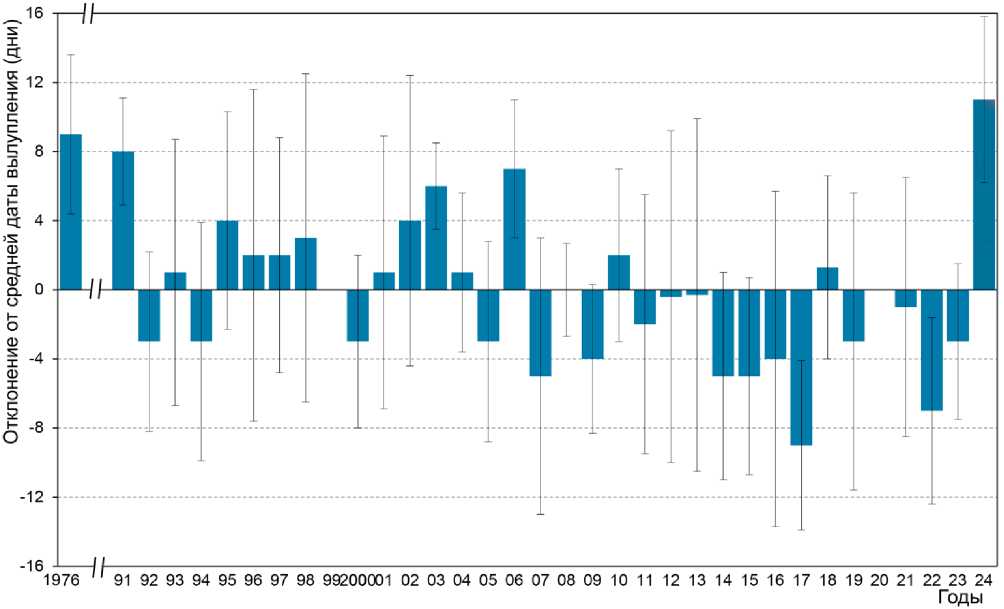
Рис. 7. Отклонение даты вылупления (от среднего значения за 1976-2013 годы – 21 июня) у малых конюг на острове Булдырь. Отрицательные значения указывают на более раннюю по сравнению со средней дату вылупления, положительные – на более позднюю. Планки погрешностей показывают стандартное отклонение от среднего значения даты вылупления за каждый год (по: Koike et al. 2025)
На Командорах птенцы вылупляются во второй половине июня; в конце июня в гнёздах были пуховые птенцы (Иогансен 1934). На Медном острове 28 июня 1883 из гнезда были взяты пуховой птенец возрастом в несколько дней и обогревающий его самец (Stejneger 1885).
На острове Райкоке в 2001-2003 годах вылупление птенцов начиналось в первой декаде июля (Трухин 2003).
На острове Ионы в 1974 году массовое вылупление у малых конюг происходило в середине июля (Харитонов 1980), 8-9 июля 2009 отмечены проклёв яиц и вылупление птенцов (Андреев и др. 2012).
Вылупление, по-видимому, не приурочено к определённому времени суток, так как при осмотре гнёзд, который проходил примерно в одно и то же время, встречались как мокрые, только что вылупившиеся, так и уже обсохшие птенцы (неопубл. данные автора).
От появления проклёвки скорлупы до вылупления птенца проходит от 1 до 3 суток. Вероятно, время вылупления зависит от плотности насиживания яйца птицей. Вылупление птенцов малой конюги на Булдыре происходило асинхронно (Variance ratio test: F 104,125 = 0.72, P = 0.1). Сроки вылупления птенцов, которые впоследствии успешно слетели, и погибших до оставления гнезда, не различались ( t = 0.84, n = 103, 12, P = 0.4) (Hunter et al . 2002).
Вынос скорлупы из гнезда . Зачастую скорлупа яиц исчезает из гнездовой камеры в течение 48 ч после вылупления птенца; она либо удаляется родителями, либо случайно выталкивается из гнезда в результате движений птиц (Byrd, Williams 1993). Специальные наблюдения (Конюхов и др. 2000) показывают, что в 72.7% гнездовых камер ( n = 11) скорлупа осталась внутри и была либо раздавлена, либо закатана в дальний угол. В остальных случаях через несколько дней после вылупления скорлупа обнаруживается снаружи гнездовой камеры. Это свидетельствует о том, что она не удалялась из гнездовой камеры намеренно. Если скорлупа и оказывается снаружи, то это, по-видимому, происходит случайно вследствие перемещений взрослых птиц и/или птенцов по тесной гнездовой камере. Различия в судьбе яичной скорлупы, скорее всего, связаны с тем, что в первом случае данные собирались главным образом в осыпях, где у гнездовых камер конюг несколько выходов и вероятность случайного выкатывания скорлупы наружу выше, чем в гнездовых камерах на заросших высокой и густой травой прибрежных склонах с их единственным входом. На Бабьих островах взрослые птицы не утруждают себя удалением яичной скорлупы после того, как птенцы вылупились (Oehler 2008).
Характеристика птенцов при вылуплении . Только что вылупившиеся птенцы малоподвижны, их глаза открыты, но разрез глаз у ́ же, чем у взрослых птиц. На кисти коготь есть только на втором пальце (Конюхов и др. 2000). Через 3-5 сут после вылупления птенцы становятся подвижными и при беспокойстве уходят вглубь гнездовой камеры. Вес птенца вскоре после вылупления на острове Булдырь составлял 15.6% от веса взрослых особей; в разные сезоны размножения не было обнаружено различий как в весе птенцов (ANOVA: F = 0.05, df = 2 и 57, P = 1.0), так и в длине их крыла ( t = -0.61, n = 12, 26, P = 0.5) (Hunter et al . 2002). Однодневный птенец на острове Ионы весил 16.7 г (Андреев и др. 2012).
Масса тела и размеры птенцов в возрасте 0-2 сут представлены в таблице 3.
Развитие пуховых структур. Постэмбриональный пух, отличаясь от эмбрионального пуха меньшей длиной (1-3 мм) и беловатым цветом, растёт как на птерилиях, так и на аптериях. Он является предшественником пуховидного пера praeplumulae. Аптерии покрываются пухом с краёв, от птерилий к центру. С возраста примерно 5 сут постэмбриональный пух появляется на боковой крестцовой птерилии pt. pelvica lateralis. В возрасте около 10 сут сосочки постэмбрионального пуха начинают расти по всем аптериям, отчего те синеют. В этом возрасте постэмбриональный пух есть на спинной аптерии apt. dorsale и в средней части боковой туловищной аптерии apt. truncale laterale, а также на боковой крестцовой птерилии. Кроме того, он начинает расти на подбородке, где есть также и эмбриональный пух (Конюхов и др. 2000).
Таблица 3. Вес и размеры птенцов малой конюги в возрасте 0-2 сут на острове Булдырь (среднее ± SD */ SE **, в скобках n , lim)
|
Вес, г |
Крыло, мм |
Цевка, мм |
Клюв, мм |
Год |
Источник |
|
|
Длина |
Высота |
|||||
|
15.9±0.51** (16) 11.5–20.0 |
15.3±0.28** (16) |
14.7±0.16** (16) |
5.8±0.2** |
4.7±0.05** (5) |
1993 |
Конюхов и др. 2000 |
|
(2) 12.5-14.0 |
(2) 13-15 |
(2) 14.2-14-7 |
6.6-6.8 |
(2) 4.7 (оба) |
1992 |
Byrd, Williams 1993 |
|
17.4±2.4* (58) |
– |
– |
– |
– |
1996-98 |
Hunter et al . 2002 |
|
17.6±2.6* (19) 10.7-21.2 |
– |
– |
– |
– |
1996 |
Hunter et al . 2002 |
|
17.5±2.2* (12) 14.0-21.0 |
17.0±1.0* (12) 15.7-18.8 |
– |
– |
– |
1997 |
Hunter et al . 2002 |
|
17.3±2.5* (27) 13.0-23.0 |
17.3±1.1* (27) 15.1-20.0 |
– |
– |
– |
1998 |
Hunter et al . 2002 |
При вылуплении передний край пропатагиума голый. По верхней поверхности крыла в районе верхних кроющих пропатагиума tectris marginales dorsales propotagii эмбриональные пушины растут на некотором удалении от переднего края пропатагиума. Когда эти эмбриональные пушины уже вытесняются пеньками контурных перьев, между ними и краем пропатагиума начинают расти постэмбриональные пушины. На нижней поверхности крыла ряд эмбриональных пушин, растущих на нижней пропатагиальной птерилии pt. propatagialis ventralis, отстоит от края пропатагиума примерно на 1 мм; чуть ближе растёт ещё ряд пушин, но меньшего размера. Это – постэмбриональные пушины. Впоследствии, по мере роста, постэмбриональные пушины замещаются средними и малыми нижними кроющими первостепенных и второстепенных маховых (Конюхов и др. 2000).
У птенцов есть также ряд пушин, образующих передний край боковой крестцовой птерилии от голенной pt. cruralis к спинной pt. dorsalis птерилиям. Начинается этот ряд (от района коленного сустава) с 1-2 эмбриональных пушин нормального размера, затем идут ещё 2-3 коротких эмбриональных пушины, которые раз в 5 короче первых. По мере роста птенца в этом ряду (в его верхней, дорсальной части) добавляется ещё несколько пушин, но это уже постэмбриональные пушины (Конюхов и др. 2000).
Мезоптильный пух (praepennae, тип I) представлен сегментами истончённого пуха, имеющего тёмную окраску. Стадию мезоптиля прохо- дят крупные перья – первостепенные, второстепенные, третьестепенные маховые, верхние и нижние большие кроющие маховых, рулевые, а также крупные перья плечевых птерилий (Конюхов и др. 2000).
Развитие терморегуляции . В первые дни жизни птенца один из родителей постоянно находится с ним. С возраста 7-9 сут ( n = 6) птенец в гнезде днём остаётся один. Ночью же вне зависимости от его возраста в гнездовой камере в большинстве случаев с ним находится по крайней мере один из родителей, реже – оба (Зубакин, Конюхов 1999; Конюхов и др. 2000). На Бабьих островах птенцы малой конюги остаются в гнездовой камере на весь день одни в возрасте 6 сут (Oehler 2008).
Питание птенцов
Как родители приносят корм . Как и у других видов Aethia , у малых конюг в период выкармливания птенцов развивается особое образование – шейный, или горловой мешок, служащий для переноса корма. Он впервые описан у большой конюги и белобрюшки (Портенко 1934, 2001). Шейный мешок развивается перед вылуплением птенцов как у самцов, так и у самок. Он представляет собой вырост ротовой полости бутылкообразной формы, вход в который расположен под языком. По встречам птиц с шейными мешками, заполненными кормом, очень легко определять начало вылупления птенцов. После завершения периода размножения шейный мешок уменьшается в размерах и теряет полость. Его стенки срастаются с помощью рыхлой соединительной ткани, а вход становится практически незаметен. У молодых птиц шейного мешка нет.
У малой конюги для манипуляции с собранной в шейный мешок пищей выработались специальные приспособления подъязычного аппарата. В нём параглоссалии paraglossalia лишены окостенений, тонки и эластичны. Под действием переднего m. ceratoglossus anterior и заднего m. с. posterior рожково-язычных мускулов весь энтоглоссум entoglossum способен сгибаться и проталкивать пищу в шейный мешок. Задний рожково-язычный мускул участвует в сгибании языка, а передний рожковоязычный мускул отвечает за тонкие манипуляции. Корм из шейного мешка извлекается, по-видимому, в два этапа: сначала из его заднего отдела пища выдавливается сокращением мышц стенок самого мешка, а затем в работу вступает язык. Для поворота языка вверх развился единый и мощный подъязычный мускул m. hypoglossus, полукольцом охватывающий область сустава языка (Соколов 1990).
Когда и как часто происходит кормление. На острове Булдырь малые конюги кормят птенцов главным образом поздно вечером или ранней ночью, сразу после возвращения взрослых птиц в колонию, то есть каждый из родителей кормит птенцов только один раз в сутки (Byrd, Williams 1993; Конюхов и др. 2000). Крайне редко конюги могут прилетать с кормом как рано утром, ещё в темноте, так и днём. Непонятно, дополнительное ли это кормление (помимо основного поздним вечером или в начале ночи) или же эти птицы по какой-либо причине не смогли покормить птенцов вовремя. Не ясно, собирался ли этот корм ночью для утреннего кормления или же корм собирался вечером до наступления темноты и птицы просто задерживаются с возвращением в колонию (Конюхов и др. 2000).
На острове Ионы родители с кормом прилетают после 15 ч; интервалы между прилётами составляют 30-90 мин (Андреев и др. 2012).
Таблица 4. Состав корма птенцов малой конюги в разные этапы периода выкармливания; остров Булдырь, 1995-1997 годы (по: Williams et al. 1998; Hunter et al. 2002)
Этапы периода выкармливание птенцов
|
Вид добычи |
Начало ( n = 40) |
Средина ( n = 71) |
Конец ( n = 44) |
|||
|
Вес, г |
% |
Вес, г |
% |
Вес, г |
% |
|
|
Gastropoda |
||||||
|
Limacina helicina |
3.9 |
1.3 |
4.2 |
0.8 |
2.1 |
0.6 |
|
Copepoda |
||||||
|
Neocalanus cristatus |
101.6 |
34.2 |
55.9 |
10.7 |
79.3 |
22.8 |
|
N . plumchrus / N . flemingeri |
147.1 |
49.5 |
327.1 |
62.6 |
216.2 |
62 |
|
Calanus pacifica |
0 |
0 |
0.5 |
0.1 |
0 |
0 |
|
Неопределённые копеподы |
0 |
0 |
3.1 |
0.6 |
0 |
0 |
|
Всего копепод Amphipoda (Hyperiidea) |
83.7 |
74 |
84.8 |
|||
|
Hyperoche medusarum |
1.5 |
0.5 |
2.6 |
0.5 |
5.2 |
1.5 |
|
Parathemisto pacifica |
0 |
0 |
0.5 |
0.1 |
0.3 |
0.1 |
|
Primno macropa |
2.1 |
0.7 |
1.6 |
0.3 |
0.3 |
0.1 |
|
Всего амфипод Gammaridea |
1.2 |
0.9 |
1.7 |
|||
|
Talitridae |
0 |
0 |
<0.1 |
<0.1 |
0 |
0 |
|
Euphausiacea Thysanoessa spp, |
31.5 |
10.6 |
118.1 |
22.6 |
37.4 |
10.7 |
|
Decapoda |
||||||
|
Zoea креветок |
9.2 |
3.1 |
3.1 |
0.6 |
6.6 |
1.9 |
|
Zoea крабов |
0.1 |
0.1 |
1 |
0.2 |
0.7 |
0.2 |
|
Megalopa крабов |
0.3 |
0.1 |
4.7 |
0.9 |
0.3 |
0.1 |
|
Всего декапод |
3.3 |
1.7 |
2.2 |
|||
|
Общий вес, г |
297.2 |
522.4 |
348.4 |
|||
Состав пищи . Малые конюги питаются сами и кормят птенцов в основном мелкими океанскими копеподами (Day, Byrd 1989). Исследования не выявили закономерностей в относительном типе или обилии добычи на разных этапах периода выкармливания птенцов (табл. 4). Относительная доля каждого из них в разные годы не была постоянной. В 1993-1997 годах на острове Булдырь взято 199 пищевых проб, зоопланктон в которых относился к 14 различным видам. Большую часть (60.999.0%) пищи составляли три группы организмов: копеподы Neocalanus plumchrus / flemingeri и N. cristatus , а также эвфаузииды Thysanoessa spp.
Практически всегда преобладали копеподы (45.3-92.3% от общего рациона) и только в 1994 году 45% добычи составляли копеподы и чуть более 42% эвфаузииды, тогда как в 1997 году 92% добычи составляли копеподы и 1% эвфаузииды. Среди копепод в рационе птенцов преобладали Neocalanus plumchrus/flemingeri и N. cristatus . Доля первого в разные годы варьировала от 21.8 до 81%, тогда как второго от 11.3 до 36.4%. Доля эвфаузиид составляла от 0.6 до 42.4%. В 1994 году было относительно меньше N. plumchrus/flemingeri , но больше N. cristatus и Thysanoessa spp., тогда как в 1997 году эта картина изменилась на противоположную - стало больше N. plumchrus/flemingeri , реже встречался N. cristatus и почти не встречались Thysanoessa spp. Остальные группы планктона составляют незначительную часть рациона птенцов: гастроподы в основном крылоногие моллюски Limacina helicina (0.3-2.1%), бокоплавы Primno macropa, Parathemisto pacifica , Hyperoche medusarum (по мере убывания) (0.6-8.4%), декаподы (зоеа креветок и крабов) - 0.3-3.7% и только следы Gammaridea (Talitridae — морские блохи) 0.1%. Только в 1993 году в рационе птенцов на долю рыб — бровастых терпугов Hexa-grammos spp. приходилось 20.6% (Hunter et al . 2002). Копеподы остаются главным компонентом питания птенцов (табл. 5, рис. 8, 9).
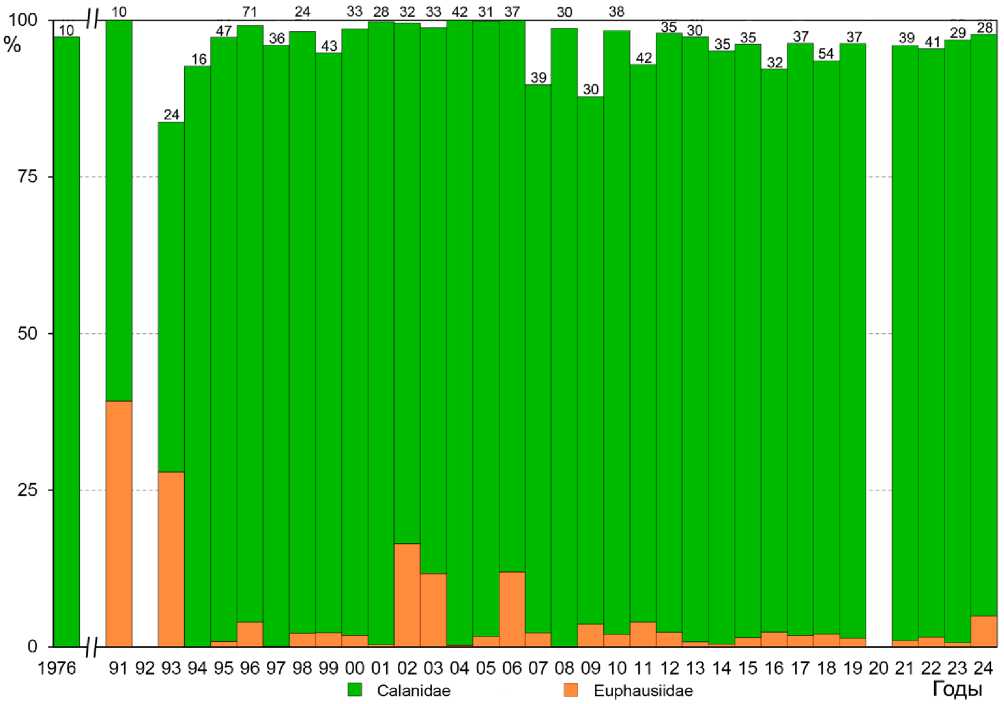
Рис. 8. Процентный состав основных видов добычи в рационе птенцов малой конюги на Булдыре. Значения выражены как процент от общего числа индивидуальных объектов добычи, приходящийся на каждый вид и сгруппирована на уровне семейства
(по: Koike et al . 2025)
Таблица 5. Относительный вес добычи в корме птенцов малых конюг; остров Булдырь, 1993-1997 годы (по: Williams et al . 1998; Hunter et al . 2002)
|
Виды |
1993 г. ( n = 24) |
1994 г. ( n = 16) |
1995 г. ( n = 48) |
1996 г. ( n = 71) |
1997 г. ( n = 36) |
|||||
|
Вес, г |
% |
Вес, г |
% |
Вес, г |
% |
Вес, г |
% |
Вес, г |
% |
|
|
Gastropoda |
||||||||||
|
Limacina helicina |
0.8 |
1.5 |
0 |
0 |
2.7 |
0.7 |
1.4 |
0.3 |
6.3 |
2.1 |
|
Неопределённые гастроподы |
0 |
0 |
0.2 |
0.2 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Copepoda |
||||||||||
|
Neocalanus cristatus |
9.9 |
18.6 |
34.2 |
36.4 |
101.5 |
26.2 |
102 |
21.2 |
33.9 |
11.3 |
|
N. plumchrus / flemingeri |
11.6 |
21.8 |
8.4 |
8.9 |
187.2 |
48.3 |
260.9 |
54.2 |
242.9 |
81 |
|
Calanus pacifica Неопределённые |
0 |
0 |
0 |
0 |
0.4 |
0.1 |
0 |
0 |
0 |
0 |
|
копеподы |
7.4 |
13.8 |
0 |
0 |
3.5 |
0.9 |
0 |
0 |
0 |
0 |
|
Всего копепод Amphipoda (Hyperiidea) |
54.2 |
45.3 |
75.5 |
75.4 |
92.3 |
|||||
|
Hyperoche medusarum |
0 |
0 |
0 |
0 |
6.6 |
1.7 |
2.4 |
0.5 |
0 |
0 |
|
Parathemisto pacifica |
2.1 |
3.9 |
0.5 |
0.5 |
0.4 |
0.1 |
.0.1 |
0.1 |
.0.1 |
0.1 |
|
Primno macropa |
0.2 |
0.3 |
7.4 |
7.9 |
0 |
0 |
0 |
3.9 |
1.3 |
|
|
Всего амфипод Gammaridea |
4.2 |
8.4 |
1.8 |
0.6 |
1.4 |
|||||
|
Talitridae |
0 |
0 |
0 |
0 |
0 |
0 |
0.5 |
0.1 |
0 |
0 |
|
Euphausiacea Thysanoessa spp, Decapoda |
10.4 |
19.5 |
39.9 |
42.4 |
71.7 |
18.5 |
113.6 |
23.6 |
1.8 |
0.6 |
|
Zoea креветок |
0 |
0 |
3.4 |
3.6 |
13.9 |
3.6 |
0.5 |
0.1 |
4.5 |
1.5 |
|
Zoea крабов |
0 |
0 |
0 |
0 |
0 |
0 |
.0.1 |
0.1 |
1.8 |
0.6 |
|
Megalopa крабов |
0 |
0 |
0 |
0 |
0 |
0 |
0.5 |
0.1 |
4.8 |
1.6 |
|
Всего декапод Actinopterygii |
3.6 |
3.6 |
0.3 |
3.7 |
||||||
|
Hexagrammos spp, |
11 |
20.6 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Общий вес, г |
53.4 |
94 |
387.9 |
481.3 |
299.9 |
|||||
Анализ 1128 пищевых проб, взятых на Булдыре с 1976 по 2024 год (в 1977-1990, 1992 и 2020 годах образцы корма не собирались) показал присутствие биологических объектов, относящиеся к 32 таксонам. Пищевые пробы состояли в основном из копепод Neocalanus и эвфаузиид Thysanoessa (рис. 8): в разные годы копеподы (в основном видов рода Neocalanus) составляли 19.14-99.8%, эвфаузииды (в основном представители рода Thysanoessa) – 0-39.2%, амфиподы – 0-4.37%, декаподы – 07.52% и гастроподы – 0-1.78%. Среди копепод основным кормовым объектом был более мелкий Neocalanus plumchrus / flemengeri (4-6.3 мм), на долю которого в разные сезоны приходилось 96.3-35.8% от основных видов добычи в рационе птенцов малой конюги, тогда как на долю более крупного Neocalanus cristatus (6.7-10.4 мм) – 0-56.7% (Bond et al . 2012; Warzybok 2011; Koike et al . 2025).
На острове Егг малые конюги кормили птенцов в основном копеподами (~89% от общей массы, n = 26), а на эвфаузиид приходилось ~9% (Dragoo et al . 2012). В пищевой пробе, взятой 16 июля 2009 на острове
Ионы, найдены десятки эвфаузиид Thysanoessa raschii и единицы ка-лянусов Calanus sp. (Андреев и др. 2012).
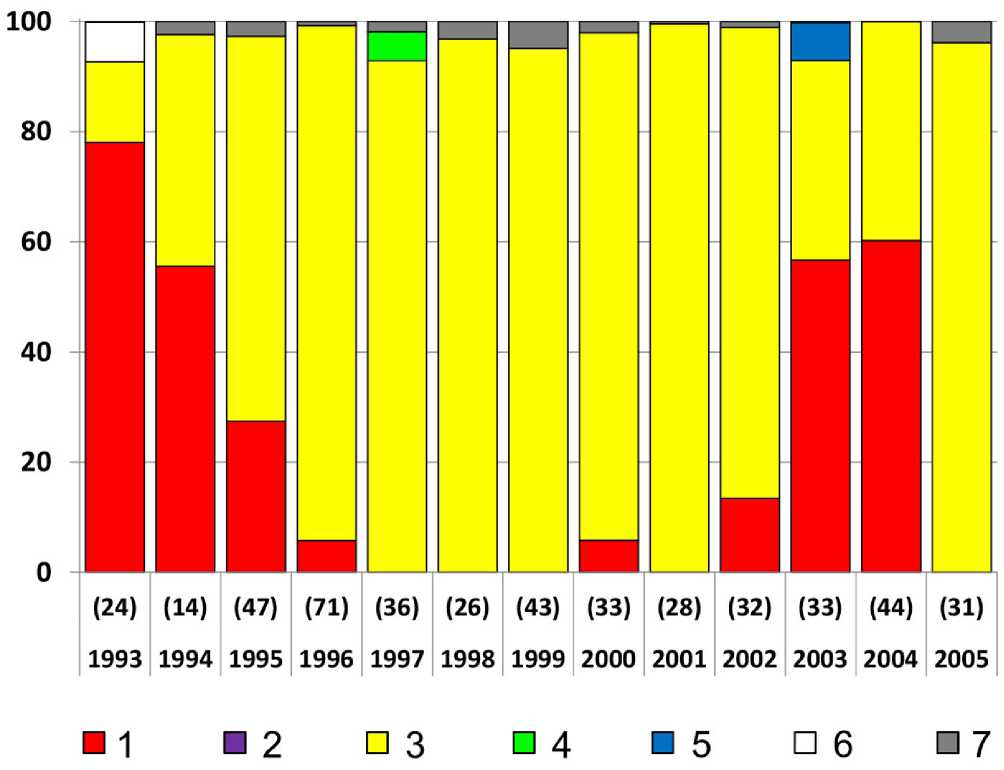
Рис. 9. Рацион (в процентах от общей массы) птенцов малой конюги на острове Булдырь. Под каждым столбцом диаграммы указаны объём выборки и год сбора данных (Dragoo et al . 2012).
1 – эвфаузииды, 2 – амфиподы, 3 – копеподы, 4 – креветки, 5 – чилимы, или пандалиды,
6 – рыба, 7 – прочее
Вес порций корма, которые приносили родители птенцам на острове Булдырь в 1993-2006 годах, несколько различался в течение периода выкармливания: в его начале (8.02±0.66 г (SE , n = 113)) он был значимо меньше, чем в его середине (10.90±0.67 г (SE , n = 108)), однако в конце (9.29±0.70 г (SE , n = 95) величина порций корма значимо не отличалась от таковой как в начале, так и в середине периода выкармливания птенцов. Средняя порция корма составляла 7.4-10.1% от веса взрослой птицы (Bond et al . 2012). Пищевые пробы, собранные на острове Касато-чий в 2007 году, весили 10.3±4.5 г (SE , n = 4, lim 6.3-16.3 г) (Drummond, Larned 2007). Поскольку малые конюги добывают корм в приливных возмущениях ближе к берегу, чем другие виды конюг (Byrd, Williams 1993), это, возможно, позволяет им переносить более тяжёлый груз, а возможно, что большая кормовая порция вызвана также тем, что малые конюги кормят птенцов реже, чем другие виды конюг.
Рост и развитие птенцов . Птенцы малой конюги характеризуются полувыводковым типом развития. На острове Булдырь во время линейной фазы роста птенцы ежедневно набирали в 1976 году 4.8 г ( n = 3-4), в 1992 – 3.5 г ( n = 8, lim 2.8-4.0 г), в 1996 – 3.3±0.8 г (SD , n = 29, lim 0.24.3 г), в 1997 – 4.0±0.7 г ( SD , n = 43, lim 2.5-5.3 г), в 1998 – 3.9±0.9 г (SD , n = 47, lim 0.7-5.2 г), в 1996-1988 годы – 3.8±0.9 г ( SD , n = 119; lim 0.745.31 г) (Byrd, Williams 1993; Hunter et al . 2002), а крыло росло в 1997 году со скоростью 3.1±0.1 мм (SD , n = 41, lim 2.2-3.8 мм), в 1998 – 3.2±0.2 мм (SD , n = 44, lim 2.5-3.7 мм), в 1997-1998 годы – 3.2±0.3 мм (SD , n = 85; lim 2.17-3.82 мм). В разные годы на Булдыре во время линейной фазы роста отмечены значительные различия в скорости увеличения массы птенцов (ANOVA: F = 7.80, df = 2 и 118, P = 0.001), в то время как в скорости роста длины крыла значимых различий не выявлено ( t = 21.66, n = 41, 44, P = 0.1) (Hunter et al . 2002). Птенцы, погибшие от голода, во время линейной фазы роста набирали вес значительнее медленнее (0.9± 1.16 г/сут; SE , n = 3), чем успешно слетевшие птенцы (3.9±0.06 г/сут; SD , n = 103) (критерий U Манна-Уитни: z = -2.73, n = 3, 103, P = 0.006). Развитие птенцов, исчезнувших из гнезда раньше срока слёта (3.5±0.46 г/сут; SD , n = 4), не отличалось от такового благополучно слетевших (критерий U Манна-Уитни: z = -0.62, n = 4, 103, P = 0.5). Эти птенцы, видимо, были съедены серокрылыми чайками Larus glaucescens (Hunter et al . 2002) или во время посещения исследователями гнёзда уходили вглубь осыпи, поскольку в конце гнездового периода они становятся подвижными и при любом беспокойстве стараются спрятаться.
В возрасте 32.5±0.81 сут (SE , n = 13), примерно за 7-10 дней до слёта, птенцы достигают пика веса (табл. 6), при котором он составляет 98% веса взрослых птиц (Byrd, Williams 1993; Конюхов и др. 2000). К этому времени у птенцов практически прекращается рост цевки, тогда как крыло и клюв продолжают расти. Птенцы, выросшие на прибрежных склонах, заросших высокой и густой травой, достигают пика массы значимо позже, чем, выросшие на осыпях ( t = 3.09; df = 11, P = 0.01), при этом статистически значимо большей у них была лишь длина крыла ( t = 3.27, df = 11, P = 0.0074;) (Конюхов и др. 2000). После этого вес птенцов постепенно снижается и перед слётом птенец в среднем терял 8.9±5.9% веса (SD , n = 75; lim 0-21.9%); к моменту слёта вес птенцов достигал 90.5% от веса взрослых особей, длина крыла 93.5% и цевки – 100%.
Развитие оперения. Характер и общее количество постэмбрионального пуха из-за его незначительного размера и быстрого замещения пуховидным пером в целом остаётся неизвестными. Вероятно, этот пух – предшественник полупуховидного и пуховидного перьев на боковых туловищных, лопаточных, спинной, грудинной и, возможно на других аптериях. Что касается птерилий, то постэмбриональный пух есть на спинной и боковой крестцовой, грудной, грудинной, но возможно, он есть и на всех остальных птерилиях. Там же, по этим птерилиям и аптериям, растёт и пуховидное перо II типа, однако определить соотношение пуховидных перьев I и II типов на живых птенцах практически невозможно. Можно только сказать, что пуховидные перья I типа преобладают на периферической части грудинной аптерии, а на боковых туловищных, на коленке и «под мышкой» – пуховидные перья II типа (Конюхов и др. 2000).
Таблица 6. Вес и размеры птенцов малой конюги при пике массы на острове Булдырь (среднее ± SD */ SE **, в скобках n , lim)
|
Вес, г |
Крыло, мм |
Цевка, мм |
Клюв, мм |
Год |
Источник |
|
|
Длина |
Высота |
|||||
|
113.9±2.1** (13) |
91.8±1.7** (13) |
21.0±0.18** (13) |
8.7±0.12** (3) |
6.53±0.09** (3) |
1933 |
Конюхов и др. 2000 |
|
113.4±7.0* (8) 100.4-122.6 |
– |
– |
– |
– |
1992 |
Byrd, Williams 1993 |
|
101.3±12.0* (33) 63-120 |
– |
– |
– |
– |
1996 |
Hunter et al . 2002 |
|
111.6±13.0* (36) 86-134 |
– |
– |
– |
– |
1997 |
Hunter et al . 2002 |
|
115.8±9.3* (36) 94-137 |
– |
– |
– |
– |
1998 |
Hunter et al . 2002 |
Поведение птенцов . Подросшие птенцы встречают прилетающих родителей громким писком. Во вторую декаду июля поздним вечером (около 0 ч 30 мин) их голоса в колонии сливаются в единый громкий хор; в третью декаду июля, с началом слёта птенцов, этот хор слабеет, и к началу августа слышны голоса только единичных птенцов (Конюхов и др. 2000). За несколько дней перед слётом, как и у других видов конюг, птенцы становятся подвижными и иногда, покидая гнездовые камеры, бродят в их окрестностях. Это наблюдалось как в осыпях, так и на заросших травой прибрежных склонах (неопубл. данные автора).
Продолжительность гнездового периода . В 1993 году птенцы малой конюги, выросшие на травянистых склонах, покинули гнездовые камеры в среднем на 2 дня позже птенцов с Основной осыпи, причём у слётков с травянистых склонов достоверно больше была только длина крыла (табл. 7).
Различия в сроках пребывания птенцов в гнезде вызваны, вероятно, особенностями их поведения перед слётом, а не влиянием гнездовых биотопов на их рост и развитие. Становясь очень подвижными за несколько дней перед слётом, птенцы в осыпях легко могут покидать гнездовую камеру и перемещаться под соседние камни, что происходит при любом беспокойстве, в том числе и со стороны исследователей. При очередном осмотре гнездовая камера оказывается пустой и в результате фиксируется дата слёта птенца, хотя на самом деле тот ещё находится в гнездовой колонии. В биотопах, где у гнездовой камеры только один вход (например, травянистые склоны), птенец не может покинуть её при беспокойстве и остаётся в камере до реального слёта. Следовательно, размеры слётков, выросших в гнездовых камерах с одним входом, более близки к реальным, то есть таким, когда они не подвергаются беспокойству со стороны исследователей (Конюхов и др. 2000). Продолжительность пребывания птенцов в гнезде представлена в таблице 8.
Таблица 7. Различия веса, размеров и возраста слётков с Основной осыпи и травянистого склона Северо-западного мыса (среднее ± SE **, в скобках n) (по: Конюхов и др. 2000)
|
Параметры |
Основная осыпь |
Северо-западный мыс |
t |
P |
df |
|
Вес, г |
102.4 ± 2.85** (9) |
109.4 ± 5.02** (5) |
1.31 |
0.215 |
12 |
|
Длина крыла, мм |
98.3±0.91** (9) |
104.5 ± 0.96** (5) |
4.32 |
<0.001 |
12 |
|
Длина цевки, мм |
21.2±0.19** (9) |
21.0 ± 0.14** (5) |
0.614 |
0.55 |
12 |
|
Возраст при слёте, сут |
36.4 ± 0.67** (10) |
38.4 ± 0.98** (5) |
1.71 |
0.112 |
13 |
Таблица 8. Продолжительность периода выкармливания птенцов на острове Булдырь (среднее ± SD */ SE **, в скобках n , lim)
|
͞Продолжительность нахождения птенцов в гнездовой камере, сут |
Год |
Источник |
|
37.1±0.57**, (15), 33-41 |
1993 |
Конюхов и др. 2000 |
|
38.8±2.04*, (98), 35-46 |
1996-1998 |
Hunter et al . 2002 |
|
39.5±2.3*, (28), 36-46 |
1996 |
Hunter et al . 2002 |
|
38.6±2.0*, (34), 35-43 |
1997 |
Hunter et al . 2002 |
|
38.5±1.8*, (36), 35-43 |
1998 |
Hunter et al . 2002 |
Выживаемость птенцов от вылупления до слёта . Из всех птенцов, находившихся под наблюдением на острове Булдырь в 1990-2001 годах, слетело 73.1±12.5% (SD , n = 21 год, lim 93-47%); по другим данным в период с 1988 по 2024 год, исключая 1999 и 2020 – 82.1±13.8% (SD , n = 35 лет, lim 100-36%) (Williams et al . 1998; Warzybok 2011; Koike et al . 2025). В других исследованиях, проводившихся в 1996-1998 годах, успешно слетели 88.7±2.0% птенцов (SD , n = 3 года); успешность слёта между годами не различалась ( χ 2 = 0.3, df = 2, P = 0.9). По данным, полученным волонтёрами, успешность слёта птенцов оказалась значительно ниже (61.3%), чем показатели, полученные профессиональными орнитологами (88.8%; χ 2 = 19.4, df = 1, P = 0.0001). В период с 1990 по 1998 год слетело 72.7% птенцов ( n = 9 лет, lim 62-82%); в разные годы успешность слёта значимо не различались ( χ 2 = 9.2, df = 8, P = 0.3) (Hunter et al . 2002). На острове Касаточий в 2007 году из 4 птенцов, находившихся под наблюдением, слетело 2 (Drummond, Larned 2007), а в 2008 году из 4 только 1 (Buchheit, Ford 2008).
Гибель птенцов. В разные годы на острове Булдырь 11.2% (14 из 125) птенцов малых конюг не слетели. Из них 9 (64%) погибли в гнёздах, а 5 (36%) исчезли в возрасте 13-25 сут, то есть до самого раннего срока слё- та, который приходится на 32 сут; видимо, были съедены серокрылыми чайками. Семь из погибших птенцов, по-видимому, умерли от истощения (см. «Рост и развитие»). У оставшихся 2 птенцов, найденных мёртвыми, были множественные повреждения кожи на голове и туловище, а один птенец был ещё и растоптан. Такие повреждения могли нанести большие конюги. Маловероятно, что один из этих птенцов погиб от истощения, поскольку набирал 3.6 г/сут в течение периода линейного роста, что было близко к среднему показателю для успешно слетевших птенцов. Кроме того, у 2 вполне здоровых птенцов были обнаружены следы от клювов, характерные для нападений больших конюг. В обоих случаях повреждения зажили, и птенцы успешно слетели (Hunter et al. 2002). В 1990-1997 годах из 391 вылупившегося птенца 63 птенца исчезли из гнездовой камеры, а 26 умерли (Williams et al. 1998).
Поскольку большинство колоний морских птиц расположены на вулканических островах или вблизи вулканов, кладки и птенцы могут быть погребены под выбросами при извержениях, как это произошло на острове Касаточий в 2008 году, когда весь остров засыпало многометровым слоем тефры. Гибель птенцов стала основным прямым последствием извержения. Поскольку оно произошло 7-8 августа, то есть в конце периода размножения, большинство птенцов к этому времени уже слетело. К тому же извержение произошло днём, когда взрослые птицы и слетевшие птенцы кормились вдали от острова, что существенно снизило масштабы потерь (Williams et al . 2010).
Слётки
Состояние птенцов при слёте . Размеры и вес слётков представлены в таблице 9. Вес птенцов перед слётом снижается в среднем на 8.9% ( n = 75, lim 0-21.9%) (Hunter et al . 2002); вес птенцов составляет 90.5-92% от веса взрослых, длина крыла птенцов – 93.5-94.4% от таковой взрослых, цевки – 100-101.2%, длина клюва – 98.2%, а его высота – 89.2-91.0% (Byrd, Williams 1993; Hunter et al . 2002; неопубл. данные автора). Бо ́ ль-шая длина цевки у слётков – результат того, что на костях ног у них ещё присутствует значительное количество хряща, при окостенении которого уменьшается объём и длина кости.
Вес слётков малой конюги в разные годы исследований значительно различался (ANOVA: F = 6.19, df = 2, 75, P = 0.003), но длина крыла и цевки – нет (крыло: t = -1.84, n = 30, 35, P = 0.1; цевка: F = 2.69, df = 2, 105, P = 0.1) (Hunter et al. 2002). Вызывает недоумение сообщение о слётке малой конюги весом 127 г (Карташев 1979), добытом на рассвете 27 июля 1960 на острове Медный, поскольку вес взрослых малых конюг с Командорских островов меньше, чем у этого слётка (Конюхов 2025а). Поскольку в публикации упоминается, что он был полностью оперён и только редкие пушинки сохранились лишь на крестце и шее, то это точно был слёток. Слётка малой конюги можно спутать со слётком большой конюги, однако для последнего такой вес крайне мал, и подобные особи не способны летать (Конюхов 1993). Остаётся предположить, что произошла ошибка где-то между взвешиванием птенца и публикацией данного факта.
Таблица 9. Вес и размеры слётков малой конюги на острове Булдырь (среднее ± SD */ SE **, в скобках n , lim)
|
Вес, г |
Крыло, мм |
Цевка, мм |
Клюв, мм |
Год |
Источник |
|
|
Длина |
Высота |
|||||
|
99.0±10.1* (23) |
102.5±2.9* (23) |
20.9±0.8* (23) |
9.95±0.5* (23) |
6.5±0.3* (23) |
1999 |
Неопубликованные |
|
96-106 |
19.2-22.5 |
8.4-10.9 |
6.1-7.2 |
данные автора |
||
|
104.9±2.6** (14) |
100.5±1.05** (14) |
21.1±0.13** (14) |
9.2±0.24** (5) |
6.52±0.06** (5) |
1993 |
Конюхов и др., 2000 |
|
106±6.8* (8) 92.5-113.3 |
– |
– |
– |
– |
1992 |
Byrd, Williams 1993 |
|
101.3±11.3* (69) 69-122 |
102.2±4.3* (65) 90.0-110.7 |
21.8±0.6* (106) 20.0-23.5 |
– |
– |
1994-98 |
Hunter et al . 2002 |
|
90.7±9.5* (10) 74-103 |
– |
22.0±0.4* (32) 20.9-22.8 |
– |
– |
1996 |
Hunter et al . 2002 |
|
101.6±13.6* (31) 69-122 |
101.2±4.7* (30) 90.0-108.1 |
21.9±0.1* (38) 20.2-23.5 |
– |
– |
1997 |
Hunter et al . 2002 |
|
104.0±7.4* (35) 88-120 |
103.0±3.8* (35) 90.1-110.7 |
21.7±0.6* (36) 20.0-22.9 |
– |
– |
1998 |
Hunter et al . 2002 |
Таблица 10. Сроки слёта птенцов малой конюги (среднее ± SD */ SE **, в скобках n , lim)
|
Дата |
Медиана |
Год |
Источник |
|
Остров Булдырь |
|||
|
25.07±1.8**, (16), 17.07-12.08 |
24.07 |
1993 |
Конюхов и др. 2000 |
|
31.07±1.3*, (23), 24.07-19.08 |
29.07 |
1999 |
Неопубликованные данные автора |
|
02.08±0.9** (26) |
– |
1991 |
Hipfner, Byrd 1993 |
|
29.07±7.6*, (33), 20.07-22.08 |
– |
1996 |
Hunter et al. 2002 |
|
29.07±4.3*, (35), 20.07-12.08 |
– |
1997 |
Hunter et al . 2002 |
|
27.07±3.5*, (36), 20.07-05.08 |
– |
1998 |
Hunter et al . 2002 |
|
28.07±5.41*, (104), 20.07-12.08 |
– |
1996-98 |
Hunter et al . 2002 |
|
(5), 18.07-27.07 |
28.07 |
1990 |
Williams et al . 1998 |
|
03.08±4.2, (23), 24.07-08.08 |
04.08 |
1991 |
Williams et al . 1998 |
|
(33), 13.07-05.08 |
26.07 |
1992 |
Williams et al . 1998 |
|
(31), 15.07-12.08 |
27.07 |
1993 |
Williams et al . 1998 |
|
(44), 15.07-14.08 |
23.07 |
1994 |
Williams et al . 1998 |
|
(45), 21.07-16.08 |
30.07 |
1995 |
Williams et al. 1998 |
|
(40), 20.07-14.08 |
26.07 |
1996 |
Williams et al . 1998 |
|
30.07±5.9*, (59), 24.07-14.08 |
29.07 |
1998 |
Williams et al . 1998 |
|
31.07±9.3*, (41), 19.07-27.08 |
29.07 |
1990 |
Williams et al . 1998 |
|
Остров Медный |
|||
|
27.07 (слёт 2 птенцов) |
– |
1960 |
Карташев 1979 |
|
Остров Ионы |
|||
|
25.08 (значительная часть птенцов уже покинула гнезда) |
– |
1971 |
Нечаев, Тимофеева 1973 |
|
14.08 (плавающие у острова молодые) |
– |
1974 |
Харитонов 1980 |
Побудительные причины и поведение перед слётом . Прямых наблюдений нет. Видимо, птенцы покидают гнездовую камеру самостоятельно, без каких-либо побуждающих стимулов со стороны родителей.
Календарные сроки слёта . Время слёта птенцов малой конюги представлены в таблице 10. Птенцы на острове Булдырь в 1999 году (поздняя весна) покинули гнёзда значимо позже, чем в 1993 году (нормальный) ( t = 3.01; P = 0.0046): 31 июля ± 5.99 сут (SD , n = 23) и 25 июля ±7.2 сут (SD , n = 16), соответственно (неопубл. данные автора). Сдвиг сезона размножения был вызван экстремальными погодными условиями: предыдущая зима была многоснежной, а весна – холодной, что привело к позднему освобождению гнездовых камер от снега. До конца июля 1993 года покинули гнездовые камеры более 85% птенцов. Последний слёток, покидающий остров, был пойман на прибрежных скалах 26 августа во время предутреннего отлова молодых малых конюг (Конюхов и др. 2000). К сожалению, точные данные есть только для Булдыря, для других точек ареала – только косвенные. Два слётка ночью 21 июля 2007 прилетели на борт научно-исследовательского судна «Tiglax», стоявшего на якоре у острова Конюжий, а ещё один слёток замечен на воде 22 июля. Первый слёток у берега острова был замечен на воде 15 июля 2003 и 23 июля 2007, а 26 июля 2007 ночью встречен слёток, идущий по тропе перед осыпью (Drummond, Larned 2007). Сроки слёта птенцов на Командорских островах, по-видимому, сходны со сроками, отмеченными на острове Булдырь. К началу июля (середине июля по новому стилю) молодые малые конюги уже поднялись на крыло (Бианки 1909). В середине-конце июля молодые спускались на воду и отходили от берегов (Иогансен 1934). Полуоперившийся птенец малой конюги весом в 104 г и длиной 10-го первостепенного махового в 37 мм, найденный в узкой щели скалистого берегового обрыва близ села Преображенское на острове Медный 12 июля 1960 (Карташев 1979), должен был слететь 2-3 августа, что следует из времени, необходимого для достижения маховыми нормальной длины при известной скорости их роста. Ещё 2 слетающих птенцов наблюдали на рассвете 27 июля на Медном острове, а хорошо летающая молодая птица без всяких следов птенцового пуха села ночью 31 июля на палубу судна близ южной оконечности острова Беринга (Карташев 1979). На Курилах C.Бергман (Bergman 1935 – цит. по: Гизенко 1955) 7 августа добывал молодых, имевших полное оперение взрослых птиц, исключая неразвитые хохолки. В Охотском море сроки слёта, да и, видимо, всего сезона размножения, сдвинуты примерно на 2-3 недели по сравнению с Алеутами. На острове Ионы в конце августа 1971 года значительная часть птенцов уже покинула гнёзда и держалась на воде около острова (Нечаев, Тимофеева 1973). С.П. Харитонов (1980) наблюдал молодых птиц в акватории острова 14 августа 1974. Две молодые малые конюги, добытые на острове 22 августа и 2
сентября, были в свежем пере, но их 10-е и 9-е (в статье 1-2-е; разное направление нумерации маховых) первостепенные маховые перья ещё не достигли нормальной длины (Нечаев, Тимофеева 1973).
Время суток слёта птенцов . Птенцы на острове Булдырь, видимо, слетают только в предутренние часы, во время слёта с колонии взрослых птиц. Слёта в другое время отмечено не было (Конюхов и др. 2000). На Командорах спуск птенцов на воду был отмечен также на рассвете (Карташев 1979).
Синхронизация слёта . Слёт малых конюг в 1991 году на Булдыре был менее синхронным, чем у больших конюг или конюг-крошек, возможно, потому что они использовали более разнообразные места гнездования (Hipfner, Byrd 1993); в 1996-1998 годах число дней, в течение которых происходило вылупление, не отличалось от числа дней, в течение которых происходил слёт птенцов, то есть вылупление и слёт были одинаково асинхронны (Variance ratio test: F 104,125 = 0.72, P = 0.1) F = 0.72, df = 104 и 125, P = 0.1 (Hunter et al . 2002)
Успешность слёта . Данных по успешности слёта от гнезда до воды нет. Можно предположить, что все птенцы, слетающие рано утром в густых сумерках, успешно добираются до моря. Те птенцы, что слетают позже, подвергаются нападению серокрылых чаек, которые могут преследовать слетающих птиц несколько сотен метров.
Таблица 11. Успешность размножения малых конюг на острове Булдырь
|
Hunter et al . 2002; Bond et al . 2011 |
Koike et al . 2025 |
|||||||
|
Год |
Успешность |
n |
Год |
Успешность |
n |
Год |
Успешность |
n |
|
1990 |
0.56 |
9 |
1988 |
1 |
1 |
2007 |
0,85 |
53 |
|
1991 |
0.50 |
46 |
1989 |
0,25 |
1 |
2008 |
0,92 |
57 |
|
1992 |
0.60 |
55 |
1990 |
0,5 |
3 |
2009 |
0,85 |
75 |
|
1993 |
0.57 |
54 |
1991 |
0,51 |
18 |
2010 |
0,67 |
58 |
|
1994 |
0.74 |
57 |
1992 |
0,53 |
17 |
2011 |
0,61 |
36 |
|
1995 |
0.67 |
67 |
1993 |
0,59 |
26 |
2012 |
0,71 |
42 |
|
1996 |
0.70 |
57 |
1994 |
0,78 |
40 |
2013 |
0,64 |
47 |
|
1997 |
0.66 |
90 |
1995 |
0,72 |
44 |
2014 |
0,55 |
43 |
|
1998 |
0.53 |
78 |
1996 |
0,73 |
32 |
2015 |
0,71 |
53 |
|
2000 |
0.46 |
70 |
1997 |
0,8 |
56 |
2016 |
0,72 |
44 |
|
2001 |
0.36 |
75 |
1998 |
0,64 |
42 |
2017 |
0,78 |
58 |
|
2002 |
0.48 |
100 |
2000 |
0,49 |
29 |
2018 |
0,77 |
67 |
|
2003 |
0.55 |
44 |
2001 |
0,38 |
24 |
2019 |
0,85 |
82 |
|
2004 |
0.53 |
66 |
2002 |
0,58 |
47 |
2021 |
0,86 |
89 |
|
2005 |
0.76 |
70 |
2003 |
0,63 |
24 |
2022 |
0,83 |
85 |
|
2006 |
0.62 |
68 |
2004 |
0,63 |
35 |
2023 |
0,71 |
87 |
|
2007 |
0.85 |
67 |
2005 |
0,8 |
48 |
2024 |
0,33 |
31 |
|
2008 |
0.84 |
68 |
2006 |
0,64 |
39 |
|||
Общая успешность размножения. На острове Булдырь успешность размножения малых конюг в период с 1990 года по 2011 год составила 62.2±13.1% (SD, n = 21 год, lim 85-36%); по другим данным, охватываю- щим период с 1988 по 2024 год, исключая 1999 и 2020 – 67.3±13.8% (SD, n = 35 лет, lim 100-25%) (Williams et al. 1998; Warzybok 2011; Koike et al. 2025). В таблице 11 представлены показатели успешности размножения малых конюг. Несмотря на совпадение некоторых лет сбора данных мониторинг в каждом случае проводился в разных гнёздах.
Успешность размножения малых конюг на Булдыре положительно коррелировала с температурой поверхности моря с марта по май в западной части Берингова моря (рис. 10), а также отрицательно коррелировала с низким давлением над уровнем моря в восточной части Тихого океана (Jones et al . 2007; Bond et al . 2011).
Рис. 10. Корреляция между продуктивностью малых конюг
(число слётков на 1 отложенное яйцо) и средней температурой поверхности моря (SST), окружающего остров Булдырь в 1992-2003 годы, за предшествующий август-апрель.
Продуктивность = 0.351×SST -1.45, r 2 = 0.584, P = 0.007 (по: Jones et al . 2007)
Продолжительность сезона размножения . Поскольку время начала откладки яиц остаётся неизвестным, то можно предположить, что сезон размножения малых конюг на острове Булдырь длится с середины мая (Williams et al . 1994) до середины августа.
Послегнездовой период жизни слётков
Забота родителей о птенцах после их слёта . После того как птенец покидает гнездо, он становится самостоятельным.
Активность молодых на суше (в колонии) после слёта. Малые ко-нюги уникальны среди чистиковых: повторный отлов окольцованных молодых птиц доказал, что сразу же после слёта они начинают возвращаться на остров на ночёвку (Конюхов, Зубакин 1994). Молодые птицы были встречены во всех гнездовых биотопах, а также на глинистых обрывах, где пригодных для гнездования щелей нет. Число молодых ко-нюг на берегу день ото дня может значительно флуктуировать, причём пики численности отмечались каждые 5-8 дней (рис. 11). На птиц оказывала влияние погода и пики численности не совпадали с лунными, ветреными или дождливыми ночами. Те немногочисленные птицы, которые присутствовали на берегу в штормовые ночи, прятались от ветра в различных укрытиях с подветренной стороны скал.
Рис. 11. Число молодых малых конюг, встреченных на постоянном маршруте (4, 5, 8, 13, 19, и 24 августа учёты не проводились) (по: Зубакин, Конюхов 2001)
В отличие от больших конюг и конюг-крошек, начинающих принимать участие в социальной активности колонии только в возрасте около 2 лет (Bedard, Sealy 1984; Конюхов 1993; неопубл. данные автора), молодые малые конюги не только прилетают на остров, но и посещают гнездовые биотопы, где вместе со взрослыми птицами участвуют в групповой вокализации. Они в акустическом взаимодействии используют не птенцовый писк, а трельки, звучащие почти как у взрослых особей (Конюхов, Зубакин 1994; Зубакин, Конюхов 2001).
Слётки, возвратившиеся на ночлег на скалах, находящихся за сеткой, в которую ловили прилетающих в колонию с моря взрослых малых конюг, на следующий год были пойманы той же самой сеткой (Jones et al . 2007), что подтверждает предположение, что у слётков вскоре после слёта происходит запечатление мест их будущего гнездования (Зубакин, Конюхов 2001). Л.Стейнегер (1885) предполагал, что слётки, пока находятся вблизи суши, проводят ночи в норах.
Рост и развитие после слёта (начало самостоятельной жизни). У молодых, покинувших гнездовые камеры, разные части тела продолжают расти с разной скоростью. Примерно через месяц после слёта цевка остаётся практически такой же, как у слётков, а клюв и крыло продолжают расти (табл. 12). Увеличение длины крыла происходит главным обра- зом за счёт роста 10-го первостепенного махового пера, длина которого увеличивается с 91.5 до 98.6% от дефинитивной; 1-е первостепенное практически не растёт, увеличиваясь с 97.3 до 99.4%. Вершина крыла постепенно переходит от 9-го к 10-му первостепенному маховому и крыло по форме приближается к крылу взрослых птиц; это говорит о том, что способности молодых особей к полёту приближаются к таковым взрослых птиц. У некоторых молодых особей начинают расти перья косиц, а у многих из них также и хохолок (Зубакин, Конюхов 2001).
Таблица 12. Различия между молодыми птицами, пойманными в разное время послегнездового периода ( n = 35) (по: Зубакин, Конюхов 2001)
|
Показатели |
Август |
t |
P |
df |
|
|
Начало |
Конец |
||||
|
Вес, г |
97.6±1.18 |
107.0±1.4 |
5.19 |
<0.001 |
68 |
|
Длина цевки, мм |
21.3±0.12 |
21.2±0.12 |
0.83 |
0.41 |
68 |
|
Длина клюва, мм |
9.1±0.07 |
9.5±0.06 |
4.5 |
<0.001 |
68 |
|
Высота клюва, мм |
6.5±0.04 |
6.7±0.05 |
2.62 |
0.011 |
68 |
|
Длина крыла, мм |
106.3±0.46 |
107.4±0.24 |
2.13 |
0.036 |
68 |
|
Длина 1-го махового, мм |
48.7±0.41 |
48.7±0.24 |
0.13 |
0.89 |
68 |
|
Длина 10-го махового, мм |
60.8±0.47 |
63.2±0.48 |
3.55 |
0.0007 |
68 |
Таблица 13. Размеры молодых малых конюг вскоре после слёта на острове Егг в 1992-1993 годах (среднее ± SD *, в скобках n) (по: Pitocchelli et al . 2003)
|
Пол |
Вес, г |
Крыло, мм |
Цевка, мм |
Клюв, мм |
|
|
Длина |
Высота |
||||
♂♂ 109.1±8.24* (10) 109.8±2.49* (10) 21.27±0.81* (10) 10.0±0.79* (10) 6.5±0.39* (10)
♀♀ 101.5±8.89* (4) 106.6±2.61* (5) 21.18±0.76* (5) 9.96±0.81* (5) 6.32±0.13* (5)
Исследования на острове Егг проводились с 10 по 25 августа, когда большинство птенцов слетело и отлавливались молодые птицы, возвращающихся в колонию на ночёвку. Возраст определялся по внешнему виду птиц. Молодые малые конюги отличалась от взрослых некоторыми морфометрическими параметрами и весом. У них отсутствовала как линька маховых, так и украшающие образования, а также клюв имел тёмную окраску, но размер скелетные элементов очень схож (Pitocchelli et al . 2003). Промеры этих птиц представлены в таблице 13.
Вес тела молодых птиц в течение первого месяца после слёта первоначально снижается, а затем возрастает, что, вероятно, связано со стрессом, связанным с переходом к самостоятельной жизни в море (Зубакин, Конюхов 2001).
Верность слётков гнездовой колонии . Есть только косвенные данные. Из 154 молодых птиц, пойманных на ночёвке на берегу в августе 1993 года, в июле 1999 года на том же маршруте повторно были пойманы 4
птицы, причём одна из них на расстоянии около 50 м от места её поимки в 1993 году. Отмечу, что длина всего маршрута составляла около 300 м (неопубл. данные автора).
Разлёт слётков . Данных нет.
Перемещение между колониями . Мечение малых конюг геологгера-ми показало, что птицы, гнездящиеся на Булдыре, в течение всего года находятся в акватории этого острова (Schacter, Jones 2018). Такой образ жизни характерен для колоний на западных Алеутских островах, поскольку плотность малых конюг в акватории этих островов летом и зимой примерно одинакова (Renner et al . 2008). На самом востоке ареала явно происходят сезонные перемещения малых конюг. Зимой в акватории Андреяновских островов численность этих птиц значительно возрастает, тогда как в районе Четырёхсопочных островов – снижается (Byrd, Gibson 1980).
Послегнездовой период взрослых малых конюг
В послегнездовой период (конец июля – август и, видимо, позже) малые конюги посещают те же прибрежные глинистые и скальные обрывы, что и во время сезона размножения. Посещаются также гнездовые камеры – как те, где размножение было успешным, так и те, где потомство птиц погибло. Помимо этого, малые конюги посещают и гнездовые камеры, где они в данном сезоне не размножались, поскольку те были заняты белобрюшками. В тоже время после окончания сезона размножения посещаются далеко не все гнездовые камеры малых конюг. В прибрежных осыпях в первую половину августа из 15 гнездовых камер каждую ночь посещались 2, не каждую ночь – 6, а 7 камер не посещались ни разу. В конце августа птицы за ночь посещали не более 3-4 камер. На задернованном склоне в первую половину августа за ночь посещались в среднем 61% (44-78%) гнездовых камер, в конце августа – только 19.8% (0-44%; n = 9) тех же камер. В целом чаще посещаются гнездовые камеры в центральной части колоний, чем на их периферии. Остаётся открытым вопрос, кто посещает все эти гнездовые камеры: бывшие хозяева или же пришлые особи (Зубакин, Конюхов 1994).
Увеличение уровня вокализации малых конюг в колонии в начале послегнездового периода (Конюхов 2025б) примерно в 2 раза по сравнению с периодами насиживания и выкармливания птенцов свидетельствует о том, что у взрослых птиц происходит активный поиск мест для будущего гнездования и перераспределение занятых гнездовых камер; к концу августа интенсивность вокализации резко снижается (Зубакин, Конюхов 1994).
Конец сезона размножения (оставление колоний). Сроки, в которые малые конюги покидают гнездовые биотопы, остаются неизвестными. В той части ареала, где эти птицы оседлы, они продолжают посещать ко- лонии и зимой. Там же, где море покрывается льдом, они, видимо, покидают окрестности колоний после начала ледостава либо когда замерзают все трещины и полыньи.
Я искренне благодарен сотрудникам Аляскинского морского национального природного заказника (Alaska Maritime National Wildlife Refuge) за предоставленную возможность работы на острове Булдырь, а также за техническую поддержку исследований. Автор крайне признателен Ю.Б.Артюхину, В.А.Зубакину, К.Е.Литвину и С.П.Харитонову за фотографии, замечания и ценные советы во время написания данной публикации.
