Птицы Западных Гималаев (по итогам экспедиций Санкт-Петербургского союза учёных, 2011-2015 годы)
")
Автор: А.В. Андреев
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 2571 т.34, 2025 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140312629
IDR: 140312629
The birds of Western Himalaya as observed during expeditions of the St. Petersburg Association of Scientists & Scholars in 2011-2015
Текст статьи Птицы Западных Гималаев (по итогам экспедиций Санкт-Петербургского союза учёных, 2011-2015 годы)
Александр Владимирович Андреев . Институт биологических проблем Севера ДВО РАН, Магадан, Россия
Второе издание. Первая публикация в 201 7*
На северо-западной окраине Гималаев сходятся границы нескольких крупных естественно-исторических областей Азии. Насыщенное многообразие климатических, орографических и ландшафтно-растительных условий позволило сформироваться здесь одному из очагов биоразнообразия планетарной значимости (Mani 1974). Фауна птиц – его неотъемлемая и весьма заметная часть. Своими истоками гималайская орнитология восходит к середине XIX века, когда на волне европейского естественно-исторического энтузиазма и британской экспансии в Азию были собраны основополагающие научные коллекции птиц Непала и северозападных провинций Индии (Adams 1859; Stoliczka 1866; von Pelzeln 1868).
Хотя первые сводки по авифауне Гималаев появились в конце XIX века, фаунистический этап этой многотрудной работы продолжался ещё в течение многих десятилетий (Osmaston 1925; Whistler 1925; Hellmayr 1929; Koelz 1940) и не завершился поныне. Он послужил основой к развитию современных исследований по истории восточноазиатских авифаун (Vaurie 1972), эволюции отдельных таксонов (Ruan et al . 2005; Tietze et al . 2006; Drovetski et al . 2013) и сохранению биоразнообразия (Stattes-field et al . 1998).
К настоящему времени гималайская фаунистика сделалась частью процветающего туристического бизнеса и фотоискусства (Ali 1996; Grim-met, Inskip 2003). Однако коммерция XXI века не отменяет научной ценности фаунистических наблюдений: в условиях постоянной трансформации и утраты естественных местообитаний эти данные играют важную роль в осуществлении природоохранной политики (Collar et al . 2001; BirdLife International 2004).
Российское участие в гималайских исследованиях британского периода ограничено краткими заметками В.Ф.Новицкого (1903) на пути из Кашмира в Фергану. В течение большей части ХХ века по причинам, от вопросов естественной истории далёким, интерес отечественных зоологов к их постановке и решению ограничивался изучением коллекций Зоологического института АН СССР в Ленинграде (Штегман 1938; Козлова 1975), Зоологического музея Московского университета (Бёме 1975), а также полевыми исследованиями на южных рубежах среднеазиатских республик (Потапов 1968; Ковшарь 1981), в горах южной Сибири (Сушкин 1938) и Дальнего Востока (Назаренко 1985). Лишь в середине 1990-х годов у российских учёных появилась возможность своими глазами увидеть самобытную фауну Гималайских гор и получить существенные научные результаты (Коблик и др. 2000; Андреев 2012; Романов и др. 2016).
С 2011 года в развитии гималайской тематики принял участие Санкт-Петербургский союз учёных (СПбСУ). В сентябре-октябре 2011, мае-июне 2013 и июне-июле 2015 года автор принял участие в трёх экспедициях Центра гималайских научных исследований СПбСУ на северо-запад Индии (штат Химачал-Прадеш и штат Джамму и Кашмир). Маршруты ещё одной экспедиции, состоявшейся в феврале 2014 года, лежали в пределах Индо-Гангской равнины (штаты Харьяна, Гуджарат, Раджастан). Эта поездка дала возможность познакомиться с «фоновым» состоянием авифауны Индостана и увидеть её связь с фауной гималайских предгорий, удалённых почти на 1200 км к северо-востоку.
Маршрутами наших гималайских поездок (см.: Боркин 2014; Боркин, Ганнибал 2016) были охвачены три основных экорегиона северо-западной части Гималаев: «предгорный» (500-1800 м над уровнем моря), «склоновый» (1800-3700 м) и «нагорный» (3000-5000 м). На пространстве между 30-35° с.ш. и 74-79° в.д. удалось осуществить три пересечения горных цепей, отделяющих Индо-Гангскую равнину от нагорий Малого Тибета и Ладака (долина реки Сатледж, перевалы Рохтанг и Зоджи-Ла). Общая протяжённость маршрутов составила около 3.5 тыс. км, площадь «освоенного» пространства оказалась близка к 150 тыс. км2. Хотя при избранном темпе перемещений на экскурсии и изучение птиц оставалось лишь 3-4 ч в день, такая «придорожная фаунистика», сопровождаемая регистрацией всех встреченных видов птиц на фотокамеру, позволила получить представление о составе и современном состоянии «верхнего среза» гималайской фауны птиц.
Всего зарегистрировано 759 встреч с птицами 188 видов («встречей» считали регистрацию одной или нескольких особей одного вида в течение одной экскурсии), что составляет около 50% регионального списка видов (Grimmet, Inskip 2003).
На южном макросклоне Гималаев в садовых и лесных местообитаниях с преобладанием деодара, ели Смита и голубой сосны (окрестности селения Ракчам в долине Баспы, города Наггар в долине Куллу в штате Химачал-Прадеш, Гульмарг, Сонамарг в Кашмире и другие; всего 30 экскурсий) отмечены 45 видов. Наиболее обычными были серый чекан Saxicola ferrea , чёрный турач Francolinus francolinus и большой борода-стик Megalaima virens . В прибрежных (ручьевых) биотопах – ручьевая Rhyacornis fuliginosus и водяная Chaimarrornis leucocephalus горихвостки, а также бурая оляпка Cinclus pallasi . Среди гималайских эндемиков наиболее заметными были дендрофильные виды: желтоклювая голубая сорока Urocissa flavirostris , рыжешейная синица Periparus rufo-nuchalis и рыжехвостая мухоловка Ficedula ruficauda .
В предгорных и склоновых ландшафтах отмечено 17 общих видов (коэффициент сходства 24%). Среди последних наиболее обыкновенными были гималайские эндемики: исчерченная кустарница Garrulax linea-tus и белощёкий бюльбюль Pycnonotus leucogenys .
В нагорных местностях Ладака и долины Спити на севере Химачал-Прадеша (каменистые пустыни, ивняки, заросли караганы, эфедры, тамариска; 39 экскурсий) встречено 57 видов птиц. Среди обитателей открытых местообитаний наиболее обычными были горихвостка-чернушка Phoenicurus ochruros , клушица Pyrrhocorax pyrrhocorax и кеклик Alectoris chukar; из околоводных видов – огарь Tadorna ferruginea , горный гусь Anser indicus и травник Tringa totanus , а среди эндемичных форм рыжегорлая завирушка Prunella rubeculoides и горные вьюрки – гималайский Leucosticte nemoricola и жемчужный Leucosticte brandti .
Коэффициент сходства между фаунами южного макросклона Больших Гималаев и нагорий составил 20%. Среди 17 общих видов наиболее часто встречались синяя птица Myophonus caeruleus , серая синица Parus cinereus и желтоголовая трясогузка Motacilla citreola .
Оригинальные наблюдения далеки пока от желаемой полноты, но позволяют, тем не менее, наметить положение зоогеографической границы высокого ранга между северными (палеарктическими) и южными (преимущественно ориентальными) фаунами птиц в этой части Гималаев. Путём ареалогического анализа (карты ареалов заимствованы с сайта в каждой из названных ландшатно-высотных групп можно выделить подгруппы эндемичных, палеарктиче- ских и тропических («ориентальных») видов. По их численному соотношению (см. рисунок) можно видеть, что уровень эндемизма наиболее высок среди птиц горнолесных местообитаний гималайского макросклона (до 40%). В большинстве своём это лесные виды восточноазиатского, чаще индо-малайского происхождения, обособившиеся в горной тайге Гималаев. Их ареалы имеют дугообразные очертания с площадью, превышающей 150 тыс. км2.
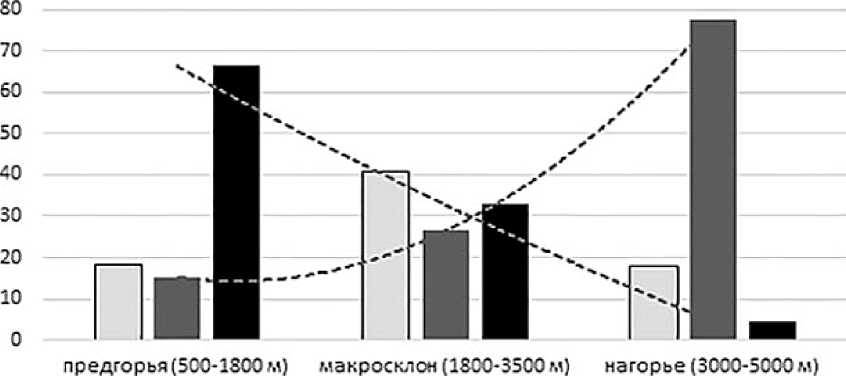
□ эндемичные ■ палеарктические ■палеотропические
Доля видов птиц разного генезиса (по вертикали, в % от числа видов, отмеченных в данном поясе) в разных высотных поясах северо-западных Гималаев (по горизонтали)
В гималайских предгорьях доля эндемичных видов примерно вполовину ниже (18%), а виды тропического генезиса абсолютно преобладают (67%). В лесной полосе макросклона соотношение между южными и северными видами выравнивается (33% и 26% соответственно). В нагорных фаунах абсолютно преобладают палеарктические виды (77%), доля эндемичных видов также относительно высока (18%), при том, что все они произошли от палеарктических групп, населяющих горы Центральной Азии.
Таким образом, нижние ступени гималайской фаунистической «лестницы» заняты тропическими видами, расселявшимися с востока и юга, а верхние – палеарктическими видами, чьи ареалы лежат западнее и севернее Гималаев. Заметим также, что палеарктические виды проникают в нижние части гор и на равнины относительно дальше, чем ориентальные виды в горы. По нашим оценкам, численное равновесие между палеарктическими и палеотропическими видами лежит в срединной части лесного пояса Гималаев в полосе абсолютных высот между 2800 и 3500 м н.у.м. Как и в других изученных случаях, «южная граница» Палеарктики в северо-западных Гималаях представляет собой мозаичную, пульсирующую и относительно широкую полосу переходного состояния.
