Пути включения миндалины и поясной извилины в центральные механизмы регуляции дыхания

Автор: Ведясова О.А., Романова И.Д., Ковалв А.М.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экологическая физиология и биохимия
Статья в выпуске: 1-4 т.11, 2009 года.
Бесплатный доступ
В работе анализируются центральные пути, опосредующие передачу влияний ядерных групп миндалевидного комплекса и топографических полей передней области поясной извилины на функционально различные структуры дыхательного центра. Предложены концептуальные блок-схемы реализации эффектов изучаемых лимбических структур на дыхание.
Миндалина, поясничная извилина, дыхание
Короткий адрес: https://sciup.org/148198430
IDR: 148198430 | УДК: 612.28+612.222.8
Pathways of the amygdala and the gyrus cinguli participation in the сentral mechanisms of the respiratory control
In the article the projections of amygdaloid complex nuclei and limbic cortex topographic fields on the structures of respiratory center have discussed. The conceptual models of the central pathways responsible for realization of the limbic structures respiratory effects are presented.
Текст научной статьи Пути включения миндалины и поясной извилины в центральные механизмы регуляции дыхания
Подчиненность системы дыхания супрабуль-барным влияниям во многом обусловлена особенностями распределения проекций от различных структур ЦНС к респираторным нейронам, вовлеченным в процессы инициации вдоха и выдоха. Например, установлена способность коры больших полушарий, подкорковых ядер, структур промежуточного, среднего и заднего мозга контролировать активность премоторных нейронов дыхательного центра (ДЦ), диафрагмальных и интеркостальных мотонейронов, и, как результат, параметров внешнего дыхания [6, 8, 9, 13,17, 21, 22, 23]. Из этого следует, что пути, нисходящие к ДЦ, имеют многоканальную организацию, благодаря чему достигается параллельный супрабульбарный контроль за локальными медуллярными и спинальными сетями респираторных нейронов. Такое устройство системы иерархических взаимодействий создает дополнительный фактор устойчивости дыхания при флуктуациях функционального состояния ЦНС или самого ДЦ в норме и при патологии.
Понимание биологического смысла респираторных эффектов, наблюдаемых при изменении морфофункционального состояния структур ЦНС, невозможно без анализа связей между высшими уровнями мозга и ДЦ. Это, в свою очередь, ставит вопрос о необходимости моделирования иерархической организации системы регуляции дыхания, что позволяет максимально приблизиться к воссозданию целостной картины центральных механизмов управления ритмикой и паттерном дыхания. Именно в этом плане выполнена настоящая работа, задача которой заключалась в анализе изменений внешнего дыхания и биоэлектрической активности инспираторных мышц у наркотизированных крыс при электростимуляции отдельных структур лимбической системы (ядра миндалевидного комплекса, поля 24 и 25 передней области поясной извили-
Ведясова Ольга Александровна , доктор биологических наук, профессор кафедры физиологии человека и животных. Романова Ирина Дмитриевна , кандидат биологических наук, доцент той же кафедры. Ковалев Александр Михайлович , аспирант той же кафедры.
ны) с позиций архитектоники их морфофункциональных взаимодействий с ДЦ.
Респираторные эффекты раздражения миндалины и поясной извилины. В ходе экспериментальной части нашего исследования установлено, что изучаемые структуры лимбики могут оказывать различные (стимулирующие и тормозящие) влияния на паттерн внешнего дыхания и активность дыхательных мышц. В частности, филогенетически древние кортикомедиальные (центральное, медиальное и кортикальное) ядра миндалины оказывают преимущественно стимулирующее действие на дыхание, тогда как филогенетически молодые базолатеральные (латеральное, базальное) ядра, напротив, тормозят эту функцию [16, 17]. Что касается поясной извилины, то доминирующим эффектом электрической стимуляции её ростро-дорсальных участков (поле 24) является угнетение деятельности ДЦ, в то время как при раздражении ростровентральных областей (поле 25) преобладают реакции активации дыхания [7, 8]. Такой характер респираторных эффектов может быть обусловлен целым рядом факторов – цитоархитектоникой и топографией изучаемых областей лимбической системы [1, 4, 5], уровнем эволюционного развития [10], особенностями нейрохимии [7, 11, 16, 24] и морфофункциональной спецификой их взаимодействий [2-4, 15, 23].
Пути передачи влияний миндалины на структуры ДЦ. Объясняя респираторные эффекты, наблюдаемые при раздражении миндалины, можно предположить, что облегчающие и угнетающие влияния её ядер на ДЦ осуществляются как через амигдало-гипоталамические проводящие пути, так и через системы прямых амигдало-бульбарных связей.
Особое значение, вероятно, играют двусторонние связи миндалины с дорсальным ядром вагуса [1, 3] и ядром солитарного тракта [31], которые обусловливают приток информации о состоянии внутренней среды организма и висцеральное подкрепление адаптивного поведения [10, 25, 26, 30]. В этом плане интересен установленный нами факт, что на фоне электростимуляции кортикомедиальных ядер миндалины у крыс происходит значи- тельное укорочение нормализованной длительности выдоха, что свидетельствует об угнетающем влиянии данных ядер на инспираторно-тормозящий рефлекс Геринга-Брейера [19], который, как известно, реализуется через структуры ваго-солитарного комплекса, расположенного в пределах ДЦ. Источником отмеченных модулирующих влияний является центральное ядро миндалины.
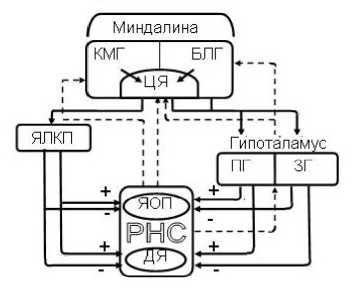
Рис 1. Блок-схема механизмов реализации влияний ядер миндалевидного комплекса на структуры респираторной нейронной сети. Обозначения: БЛГ, КМГ-базолатеральная и кортикомедиальная ядерные группы миндалины; ЦЯ - центральное ядро миндалины; ЯЛКП -ядро ложа конечной полоски; ПГ, ЗГ - передняя и задняя области гипоталамуса; РНС - респираторная нейросеть продолговатого мозга (ДЯ - двойное ядро, ЯОП - ядро одиночного пучка). Сплошные линии со стрелками - эфферентные облегчающие (+) и тормозные (-) проекции к РНС, штриховые линии - обратные связи
Опираясь на ранее полученные нами результаты экспериментов с химическим выключением ряда подкорковых структур мозга [16], допустимо считать, что для реализации модулирующего действия миндалины на ДЦ необходима целостность ядра ложа конечной полоски, входящего в состав дорсальной амигдалофугальной проекционной системы [1]. Последняя, в свою очередь, обеспечивает как прямые связи миндалины с ДЦ, так и опосредованные, в том числе, проходящие через ядра гипоталамуса и структуры лимбико-ретикулярного комплекса среднего мозга [3, 4]. Существенную роль в организации взаимодействий миндалины и ДЦ играет также вентральный амигдалофугальный путь [1, 3]. С учетом проведённого анализа экспериментальных данных нами была построена принципиальная блок-схема, в которой представлены возможные пути, опосредующие реализацию влияний ядер миндалевидного комплекса на структуры ДЦ продолговатого мозга (рис. 1).
Пути передачи влияний поясной извилины на структуры ДЦ. Исходя из данных классической физиологии [5, 20], следует считать, что регуляция дыхания лимбической корой может осуществляться за счет прямой и опосредованной передачи активирующих и тормозящих влияний полей поясной извилины к нейронам локальных нейросетей продолговатого и спинного мозга.
Говоря о путях поступления управляющих сигналов от поясной извилины непосредственно к ДЦ, следует особо выделять лимбико-бульбарные проекции, причем как прямые, так и опосредованные синаптическими переключениями в подкорке и стволе мозга [2, 4, 23]. При этом узловым пунктом в лимбико-респираторных взаимоотношениях следует считать вагосолитарный комплекс [2, 15, 27], различные ядра которого реципрокно взаимодействуют со многими клеточными группами ЦНС, посылающими прямые входы к симпатическим и вагальным преганглионарным нейронам [3, 4, 27]. Особо значима вентролатеральная часть ядра солитарного тракта, которая, являясь объектом модулирующих влияний со стороны лимбической области мозга, непосредственно участвует в регуляции дыхания [2, 3, 8, 19].
Несомненную роль в реализации респираторных эффектов поясной извилины играют ее связи с ядрами пневмотаксического комплекса варолиева моста - медиальным парабрахиальным ядром и ядром Келликера-Пьюза, целостность которых необходима для осуществления нормального дыхания типа эйпноэ, особенно при децеребрации или массивном повреждении супрабульбарных отделов мозга [32]. В ряде электрофизиологических исследований продемонстрированы нисходящие влияния указанных ядер на ДЦ [21, 22].
Влияние поясной извилины на ДЦ может осуществляться опосредованно через вегетативные [2, 3] и неспецифические [11] ядра промежуточного мозга. Субстратом этих влияний являются полиси-наптические пути, проходящие через моторные и премоторные зоны неокортекса и далее, возможно, через гипоталамус [4, 14], часть проекций которого в ДЦ имеет вазопрессин- [24] и окситоцинергиче-скую [28] природу. Специфическими мишенями, опосредующими влияния поясной извилины на ДЦ, также могут быть нейроны вентрального покрышечного поля [29] и миндалевидный комплекс [17], который следует рассматривать как подкорковый блок, параллельно подключенный к эфферентным коммуникациям лимбической области и регулирующий передачу цингулофугальных сигналов к ДЦ [12].
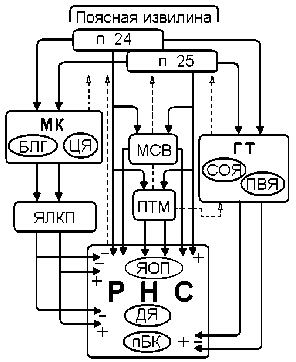
Рис. 2. Концептуальная схема механизмов реализации влияний полей передней области лимбической коры на структуры респираторной нейронной сети. Обозначения: МК – миндалевидный комплекс (БЛГ– базолатеральная группа ядер, ЦЯ – центральное ядро); ЯЛКП – ядро ложа конечной полоски; МСВ – мезенцефалическое серое вещество; ТМ – пневмотаксический механизм варолиева моста; ГТ – гипоталамус (СОЯ – супраоптическое ядро, ПВЯ – паравентрикулярное ядро); РНС – респираторная нейросеть продолговатого мозга (пБК – пре-Бетцингера комплекс, ДЯ – двойное ядро, ЯОП – ядро одиночного пучка). Сплошные линии со стрелками – эфферентные облегчающие (+) и тормозные (–) проекции к РНС, штриховые линии – обратные связи.
Анализ результатов проведенного нами исследования с позиций организации внутрицентраль-ных связей лимбики с ДЦ позволил нам разработать концептуальную модель включения дорсального и вентрального полей поясной извилины в управление бульбарной респираторной нейросетью (рис. 2).
Таким образом, внутрицентральные связи лимбической коры и миндалевидного комплекса в значительной мере обусловливают ведущее место передней области поясной извилины и центрального ядра миндалины в сложной иерархии супрабуль-барных структур, контролирующих дыхание. При этом, являясь своего рода экраном для интерференционного взаимодействия мотиваций и подкреплений, изученные лимбические структуры, способные усиливать и тормозить деятельность ДЦ, выступают в роли акцептора результата действия в процессе адаптивного поведения функциональной дыхательной системы [18]. На основании собственных и литературных данных о механизмах и путях реализации супрабульбарных влияний на висцеральные функции [5, 8, 13, 14, 15, 20], мы считаем возможным говорить о вероятностном характере объединения ДЦ с высшими отделами мозга, что является основой функциональной пластичности иерархической системы регуляции дыхания.
Список литературы Пути включения миндалины и поясной извилины в центральные механизмы регуляции дыхания
- Акмаев И.Г., Калимуллина Л.Б. Миндалевидный комплекс мозга: функциональная морфология и нейроэндокринология. М.: Наука, 1993. 264 с.
- Багаев В.А., Пантелеев С.С. Эффекты стимуляции лимбической коры на ответы нейронов ядер вагосолитарного комплекса, вызванные раздражением блуждающих нервов//Докл. РАН. 1995. Т. 340. № 44. С. 555-558.
- Баклаваджян О.Г., Аветисян Э.А., Багдасарян К.Г и др. Нейронная организация амигдало -висцеральной рефлектор дуги//Усп. физиол. наук. 1996. Т. 27. № 4. С. 51-77.
- Баклаваджян О.Г., Нерсесян Л.Б., Аветисян Э.А. и др. Нейронная организация лимбико-(цингуло)-висцеральной рефлекторной дуги//Усп. физиол. наук. 2000. Т. 31. № 4. С. 11-23.
- Беллер Н.Н. Организация и механизмы центральных эфферентных влияний на висцеральные функции. Л.: Наука, 1983. 34 с.
- Беляков В.И., Меркулова Н.А., Инюшкин А.Н. Респираторные влияния сенсомоторной коры мозга и механизмы их реализации//Бюлл. эксперим. биол. и мед. 2002. Т. 133. № 4. С. 314-317.
- Ведясова О.А. Респираторные эффекты раздражения лимбической коры и их модуляция серотонином у крыс//Бюлл. эксперим. биол. и мед. 2005. Т. 140. № 9. С. 244-246.
- Ведясова О.А., Еськов В.М., Филатова О.Е. Системный компартментно-кластерный анализ механизмов устойчивости дыхательной ритмики млекопитающих: Монография/Под ред. В.М. Еськова. Самара: ООО «Офорт», 2005. 215 с.
- Гордиевская Н.А., Буракова А.В. Конвергенция импульсов от соматических и висцеральных афферентов и ядер гипоталамуса на нейроны ядра одиночного пучка//Регуляция автономных функций: Сборник статей. Самара: Самарский университет, 1998. С. 121-130.
- Калимуллина Л.Б., Ахмадеев А.В., Минибаева З.Р., Муталова Л.П. Структурная организация миндалевидного комплекса мозга крыс//Росс. физиол. журн. им. И. М. Сеченова. 2003. Т. 89. № 1. С. 8-14.
- Кратин Ю.Г., Сотниченко Т.С. Неспецифические системы мозга. Л.: Наука, 1987. 159 с.
- Макаров Ф.Н., Варламова Т.И., Гранстрем Э.Э. Пространственная организация афферентных входов лимбической коры крыс и кошек//Росс. физиол. журн. им. И.М. Сеченова. 1997. Т. 83. № 1-2. С. 146-154.
- Меркулова Н.А., Инюшкин А.Н., Зайнулин Р.А., Кретова И.Г. Особенности и механизмы реализации респираторных влияний структур экстрапирамидной системы//Усп. физиол. наук. 2004. Т. 35. № 2. С. 22-34.
- Нерсесян Л.Б. Влияние лимбической коры и гипоталамуса на активность медуллярных дыхательных нейронов//Физиол. журн. СССР им. И.М. Сеченова. 1985. Т. 71. № 3. С. 304-309.
- Пантелеев С.С., Багаев В.А., Любашина О.А. Анализ возможных механизмов влияния передней лимбической коры на активность нейронов ваго-солитарного комплекса//Росс. физиол. журн. им. И.М. Сеченова. 1997. Т. 83. № 4. С. 33-44.
- Романова И.Д. Участие латерального ядра миндалины в регуляции активности нейронов дыхательного центра крыс//Бюлл. сибирск. мед. 2005. Т.4. Приложение 1. С. 46.
- Романова И.Д. Участие ядер миндалевидного комплекса в регуляции дыхания крыс//Нейронауки: теоретические и клинические аспекты. 2005. Т. 1. № 1. Приложение. С. 103-104.
- Судаков К.В. Голографический принцип системной организации деятельности мозга//Материалы XVII съезда физиологов России. Ростов-на-Дону, 1998. С. 365.
- Федорченко И.Д., Меркулова Н.А., Инюшкин А.Н. Амигдалофугальная модуляция инспираторно-тормозящего рефлекса Геринга-Брейера//Бюлл. эксперим. биол. и мед. 2002. Т. 133. № 4. С. 371-373.
- Черниговский В.Н. Физиологические и структурные основы кортико-висцеральных взаимоотношений//В.Н. Черниговский. Избранные труды. С-Пб.: Наука, 2007. С. 348-368.
- Якунин В.Е., Алифанов А.В., Якунина С.В. Нейрофизиологические связи субъядер Келликера-Фузе с ретикулярными и дыхательными нейронами дыхательного центра продолговатого мозга//Современные проблемы физиологии вегетативных функций: Сборник статей. Самара: Самарский университет, 2001. С. 127-139.
- Dutschmann M., Morschel M., Kron M., et al. Development of adaptive behavior of the respiratory network: implications for the pontine Kolliker-Fuse nucleus//Respir. Physiol. Neurobiol. 2004. V. 143. № 2-3. P. 155-165.
- Huang Z.-G., Subramаnian S.H., Bainave R.J., et al. Role of periaqueductal gray and nucleus tractus solitarius in cardiorespiratory function in the rat brainstem//Respir. Physiol. 2000. V. 120. No 3. P. 185-195.
- Kc P., Haxhiu M.A., Tolentino-Silva F.P., et al. Paraventricular vasopressin-containing neurons project to brain stem and spinal cord respiratory-related sites//Respir. Physiol. Neurobiol. 2002. V. 23. № 133 (1-2). P. 75-78.
- Krettek J.E., Price J.L. Amygdaloid projections to subcortical structures within the basal forebrain and brainstem in the rat and cat//J. Comp. Neurol. 1978. V.15. № 2. P. 225-254.
- LeDoux J.E., Farb C.R., Milner T.A. Ultrastructure and synaptic associations of auditory thalamo-amygdala projections in the rat//Exp. Brain. Res. 1991. V. 85. № 3. P. 577-586.
- Loewy A.D., Burton H. Nucleus of the solitary tract: efferent projections on the lower brain stem and spinal cord of the cat//J. Compar. Neurol. 1978. V. 181. P. 421-450.
- Mack S.O., Kc P., Wu M., et al. Paraventricular oxytocin neurons are involved in neuronal modulation of breathing//J. Appl. Physiol. 2002. V. 92. № 2. P. 826-834.
- Miller E.K., Cohen J.D. An integrative theory of prefrontal cortex function//Annu. Rev. Neurosci. 2001. V. 24. P. 167-202.
- Pitkanen A., Savander V., LeDoux J. E. Oraganization of intra-amygdaloid circuitries in the rat: in emerging feramefork for understanding function of the amygdala//Trends Neurosci. 1997. V. 20. № 11. P. 517-523.
- Saha S. Role of the central nucleus of the amygdala in the control of blood pressure: descending pathways to medullary cardiovascular nuclei//Clin. Exp. Pharmacol. Physiol. 2005. V. 32. № 5-6. P. 450-456.
- St-Jacques R., St-John W.M. Transient, reversible apnoea following ablation of the pre-Botzinger complex in rats//J. Physiol. 1999. V. 520. № 1. P. 303-314.
