Радиационно-индуцируемое изменение массы наземного моллюска F. fruticum M. двух возрастных групп

Автор: Черкасова Е.Е., Лаврентьева Г.В., Сынзыныс Б.И., Мирзеабасов О.А.
Рубрика: Научные статьи
Статья в выпуске: 4 т.32, 2023 года.
Бесплатный доступ
В связи с расширением принципов антропоцентрической стратегии радиационной защиты существует проблема дефицита научных данных о радиационно-индуцируемых эффектах у представителей биоты. При этом международными организациями по вопросам радиационной защиты поддерживается инициатива изучения воздействия ионизирующего излучения не только на референтные организмы, но и на других представителей биоты. Цель данной работы - изучение влияния острого g-облучения на биомассу наземного моллюска F. fruticum M. двух возрастных групп для расширения баз данных о радиоционно-индуцируемых эффектах биоты. Моллюски широко применяются при биологическом контроле радиоактивного загрязнения, а изменение массы моллюсков можно интерпретировать как релевантный показатель «заболеваемость». Возраст моллюсков определялся по количеству оборотов раковины. Облучение моллюсков проводили в диапазоне от 10 до 300 Гр с шагом 10 Гр. Изучение показателя биомассы осуществляли на протяжении 210 сут после облучения. Полученные результаты лабораторного исследования для моллюсков второй и третьей возрастных групп являются частью эксперимента, направленного на выявление особенностей радиочувствительности представителей наземной малакофауны. На протяжении всего эксперимента в контрольной группе (необлучённые особи) отмечается динамика массы моллюсков, характерная для сезонных изменений в естественных условиях. При этом наблюдается абсолютная выживаемость животных в подобранных лабораторных условиях. Выявлены дозовые диапазоны с различным проявлением изменения массы моллюсков. Дозы облучения моллюсков менее 100 Гр (вторая группа) и 120 Гр (третья группа) не индуцируют изменение изучаемого показателя. Облучение дозами 110-150 Гр и 130-150 Гр инициирует снижение массы моллюсков второй и третьей группы соответственно в период анабиоза. В дозовых диапазонах более 150 Гр установлена более сложная динамика изменения показателя в зависимости от времени после облучения и от возрастной группы моллюсков, что обусловлено особенностями проявления радиочувствительности у животных разного возраста.
Наземный моллюск, g-облучение, острое облучение, биомасса, лабораторный эксперимент, смертность, возрастные группы, релевантный показатель, радиационно-индуцируемый эффект, радиочувствительность
Короткий адрес: https://sciup.org/170201862
IDR: 170201862 | УДК: 594:612.014.482.4:57.084.1 | DOI: 10.21870/0131-3878-2023-32-4-134-146
Change in the mass of the terrestrial mollusk F. fruticum M. of two age groups after g-irradiation
There is a problem of a shortage of scientific data on radiation-induced effects in representatives of biota in connection with the expansion of the principles of the anthropocentric strategy of radiation protection. At the same time, international organizations on radiation protection issues support the initiative to study the effects of ionizing radiation not only on reference organisms, but also on other representatives of the biota. The purpose of this work is to study the effect of acute g-irradiation on the biomass of the terrestrial mollusk F. fruticum M. two age groups to expand databases on radio-induced effects of biota. Mollusks are widely used in biological control of radioactive contamination, and the change in the mass of mollusks can be interpreted as a relevant indicator of "morbidity". The age of the mollusks was determined by the number of turns of the shell. Irradiation of mollusks was carried out in the range from 10 to 300 Gy in increments of 10 Gy. The study of the biomass index was carried out for 210 days after irradiation. The obtained results of a laboratory study for mollusks of the second and third age groups are part of an experiment aimed at identifying the features of radio sensitivity of representatives of terrestrial malacofauna. Throughout the experiment, in the control group (non-irradiated individuals), the dynamics of the mass of mollusks, characteristic of seasonal changes in natural conditions, is noted. At the same time, the absolute survival of animals in selected laboratory conditions is observed. Dose ranges with different manifestations of changes in the mass of mollusks were revealed. Radiation doses less than 100 Gy (second group) and 120 Gy (third group) do not induce a change in the studied indicator. Irradiation with doses of 110-150 Gy and 130-150 Gy initiates a decrease in the mass of mollusks of the second and third groups, respectively, during suspended animation. In the dose ranges of more than 150 Gy, a more complex dynamics of changes in the indicator was established depending on the time after irradiation and on the age group of mollusks. It is due to the peculiarities of the manifestation of radio sensitivity in animals of different ages.
Текст научной статьи Радиационно-индуцируемое изменение массы наземного моллюска F. fruticum M. двух возрастных групп
В настоящее время международными организациями (МАГАТЭ, МКРЗ, НКДАР ООН) по вопросам радиационной защиты инициируется смена антропоцентрической парадигмы («защищён человек – защищена и биота») экоцентрическим подходом, в основе которого заложена концепция референтных животных и растений (RAPs – reference animals and plants) [1-3]. Данная концепция направлена на разработку методов оценки последствий на популяционном и экосистемном уровне при выявлении радиационно-индуцированных эффектов у представителей RAPs. Одним из актуальных направлений исследований в области радиационной защиты биоты является обоснование новых видов для расширения набора RAPs, который включает небольшое число референтных организмов, а именно, представителей водных экосистем (краб, бурая водоросль, форель, камбала, лягушка, утка), наземных экосистем (злаковое растение, сосна, дождевой
червь, крыса, олень, пчела). Данная работа направлена на расширение баз данных о радиационно-индуцированных эффектах у представителей биоты и содержит результаты изучения влияния Y -облучения на изменение массы представителя наземной малакофауны - моллюска Fruticicola fruticum M. Изменение массы животных является удобным показателем при оценке ответной реакции организма на воздействие различных факторов окружающей среды. При этом изменение массы организма при воздействии ионизирующего излучения может выступать индикатором заболеваемости, что является релевантным показателем, предложенным МКРЗ [1].
Следует отметить, что на фоне широкого применения представителей водной малакофа-уны в методах биологического контроля радиоактивного загрязнения водных объектов [4], изучению наземных моллюсков в контексте биоиндикации радиоактивного загрязнения наземных экосистем практически не уделяется внимания. Наземные моллюски не включены в набор RAPs, но при этом соответствуют требованиям, предъявляемым к референтным организмам: простота идентификации, широкая распространённость, короткий жизненный цикл, высокие коэффициенты накопления тяжёлых металлов и радионуклидов.
Целью данной работы является изучение влияния острого Y -облучения на биомассу наземного моллюска F. fruticum M. двух возрастных групп для расширения баз данных о радиоционно-индуцируемых эффектах биоты. Экспериментальное исследование является частью цикла работ по изучению воздействия острого и хронического у - и в -облучения на релевантные показатели наземных моллюсков [5, 6].
Материал и методы
Объектом исследований является наземный моллюск F. fruticum M . Пробоотбор особей осуществлялся на территории сопредельной с национальным парком «Угра» в Калужской области вдали от населённых пунктов, предприятий и автомобильных дорог. Отбор моллюсков осуществляли ручным способом с растительности (преимущественно с крапивы) и с почвы под растительностью, время отбора - конец лета (август). Для эксперимента отбирали особей двух возрастных групп (половозрелые особи второй и третьей группы) без видимых внешних повреждений. Возраст моллюсков устанавливали по количеству оборотов раковины [7]. На основании литературных данных были подобраны и апробированы условия содержания моллюсков в лаборатории. Методику содержания моллюсков отрабатывали в течение трёх лет перед выполнением эксперимента по облучению животных [6]. Эксперимент проводили в период с сентября по апрель, учитывая биологические ритмы животных и этапы годовой активности моллюсков (активные период перед анабиозом, анабиоз, выход из анабиоза, активный период перед размножением).
Облучение моллюсков. Облучение моллюсков проводили на Y -установке ГУР-120 в диапазоне поглощённых доз 10-300 Гр с шаговым отличием 10 Гр. Мощность поглощённой дозы составляла 30 Гр/ч. Всего было облучено 60 проб, по 15 особей моллюсков в каждой пробе. В качестве контроля выступает проба моллюсков, не подвергнутых облучению.
Определение биомассы моллюсков. Перед измерением массы моллюски промывались под проточной водой для удаления слизи, субстрата, а также для выведения особей из спящего состояния и высушивали фильтровальной бумагой перед взвешиванием. Моллюсков взвешивали на аналитических весах с точностью до 0,0002 мг. Взвешивание проб проводили каждые 15 сут после облучения в течение 210 сут. Параллельно с определением биомассы проведён анализ смертности моллюсков и рассчитан показатель ЛД 50/60 , который составляет для второй возрастной группы 115,6 ± 33,8 Гр, для третьей - 141,4 ± 26,3 Гр [5, 6].
Статистическая обработка данных. Для расчёта достоверного отличия от контроля изменение массы переводили в проценты, так как сравнение массы в граммах не отражает динамику изменения контрольной группы и групп моллюсков после облучения. Первоначальная масса принималась за 100%. Дальнейшие изменения показаны из расчёта превышения или уменьшения первоначальной массы, выраженной в процентах. Достоверность отличия значений, полученных в группах после облучения, сравнивали с такими же значениями в контрольной группе в соответствующий временной промежуток.
Экспериментальные данные обработаны с помощью программного пакета Microsoft Excel 2019. На графиках представлены средние значения и стандартная ошибка среднего. Для определения значимости различий между средними значениями изучаемых показателей и контролем использовался t-критерий Стьюдента.
Результаты и обсуждение
Изменение массы моллюсков в контрольной группе. Масса моллюсков в контрольных группах изменяется в диапазонах 0,24-0,35 г и 0,48-0,67 г во второй и третьей возрастных группах соответственно. При этом масса особей второй возрастной группы в среднем в 2 раза меньше массы животных третьей группы. Динамика изменения массы двух возрастных групп имеет схожую тенденцию в соответствии с биологическими ритмами животных (рис. 1).
га
0,8
0,7
0,6
0,5 0,4
0,3
0,2
0,1
до облучения
30 суток 60 суток 90 суток 120 суток 150 суток 180 суток 210 суток
(октябрь) (ноябрь) (декабрь) (январь) (февраль) (март) (апрель)
Время после облучения, сут (календарный месяц)
----Третья возрастная трупа ----Вторая возрастная группа
Рис. 1. Изменение массы моллюсков в контрольных группах.
Через 30 сут эксперимента масса моллюсков двух возрастных групп достоверно увеличивается в 1,1 раза по сравнению с первоначальными значениями. Во временной период наблюдений 30-150 сут отмечается снижение массы моллюсков в среднем в 1,3 раза относительно первоначального показателя, что может быть обусловлено наступлением периода анабиоза. Период 150-180 сут характеризуется достоверным повышением массы моллюсков двух возрастных групп. При этом к концу марта показатель увеличивается в 1,4 раза по сравнению с предыдущим временным периодом наблюдения и в 1,1 раза относительно первоначальной массы. Увеличение изучаемого показателя возможно связано с периодом выхода животных из анабиоза и активным потреблением пищи. До конца апреля (через 210 сут наблюдений) отмечается период достоверного снижения массы в 1,4 и 1,2 раза для второй и третьей возрастных групп соответственно.
Анализ изменения массы моллюсков (в процентном соотношении) на протяжении 210 сут эксперимента относительно первоначальной массы особей (рис. 2) позволил установить, что динамика изменения массы моллюсков второй и третьей возрастных групп имеет одинаковую тенденцию (r=0,9). На момент окончания периода наблюдения (210 сут) отмечается достоверное (p<0,05) снижение массы моллюсков в 1,3 и 1,1 раза для второй и третьей возрастных групп соответственно.
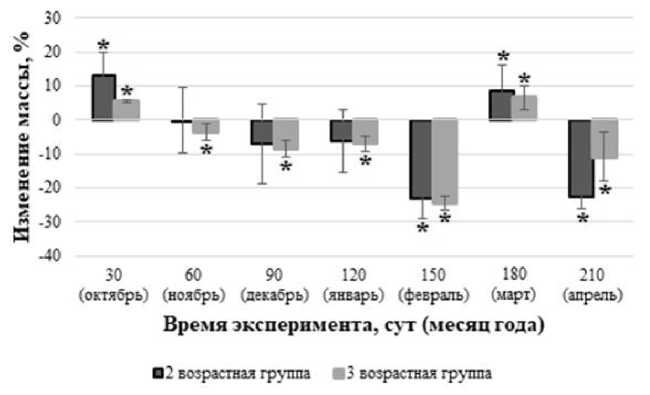
Рис. 2. Изменение массы моллюсков в контрольных группах по отношению к первоначальной массе.
Следует отметить, что на протяжении всего эксперимента отмечается абсолютная выживаемость моллюсков в контрольной группе, что обосновано адекватностью подобранных условий содержания моллюсков в лабораторных условиях. При этом динамика изменения массы в контрольной группе связана с сезонными изменениями, характерными для наземных моллюсков в естественных условиях.
Изменение массы моллюсков контрольной группы обосновано естественными биологическими закономерностями. Первоначальное повышение массы моллюсков обусловлено разнообразием пищи в лабораторных условиях, а также дополнительным внесением кальциевой подкормки. В естественной среде обитания в конце сентября разнообразие пиши, богатой кальцием, недостаточное. Избыток пищи моллюсков, обеспеченный в лабораторных условиях, приводит к увеличению потребления пищи и, соответственно, приросту массы животных. Подобный эффект отмечается в исследовании [8], где показано, что в лабораторных условиях увеличение рациона пищи у гребневика Mnemiopsis Mccradyi приводит к более активному потреблению пищи.
Известно, что в естественных условиях период анабиоза приходится на ноябрь-март [9]. В лабораторном эксперименте также установлено снижение массы животных в этот период (60-150 дней эксперимента). Период зимовки моллюсками определяется не температурным фактором, а продолжительностью светового дня, что обосновывает состояние покоя животных в период с 60 по 150 сут эксперимента (ноябрь-февраль).
После анабиоза моллюски активно набирают массу, что соответствует повышению массы на 180 сут (март) эксперимента. В естественной среде, например, активный набор массы после анабиоза наблюдается у моллюсков вида Helix lucorum Linnaeus и Helix albescens Rossmassler [9, 10], в апреле-мае наблюдается активный рост Caucasotachea vindobonensis [11].
В последующем (через 210 сут после облучения) после периода активного роста моллюсков наблюдается период замедления роста, как, например, у С. Vindobonensis [11]. Также подобное изменение ростового показателя может быть связано с началом периода размножения. Например, в естественных условиях откладка яиц Bithynia tentaculata осуществляется преимущественно с середины мая по конец июня [12]. В работе Kuźnik-Kowalska с соавт. [13] отмечается, что моллюски вида B. fruticum используют стратегию, которая предполагает вложение всех ресурсов в рост, а как только особь достигает своего предельного размера, она перестаёт расти и вкладывает все резервы в воспроизводство. Возможно, данный процесс наблюдается у всех половозрелых особей.
Изменение массы моллюска после облучения. Анализ данных был проведён с учётом изменения массы моллюсков после облучения относительно контрольных групп и между двумя возрастными группами. Выявлено закономерное изменение массы облучённых моллюсков двух возрастных групп относительно контроля, которое проявляется в четырёх дозовых диапазонах: первый диапазон – 10-100 Гр и 10-120 Гр; второй – 110-150 Гр и 120-150 Гр; третий – 160-250 Гр и 160-260 Гр; четвёртый – 260-300 Гр и 270-300 Гр для второй и третьей возрастных групп соответственно.
Облучение моллюсков дозами первого диапазона не приводит к достоверному изменению массы животных относительно контрольных групп (p<0,05). При облучении моллюсков дозами второго диапазона отмечается уменьшение массы животных через 90-120 сут (рис. 3).
Вторая возрастная группа
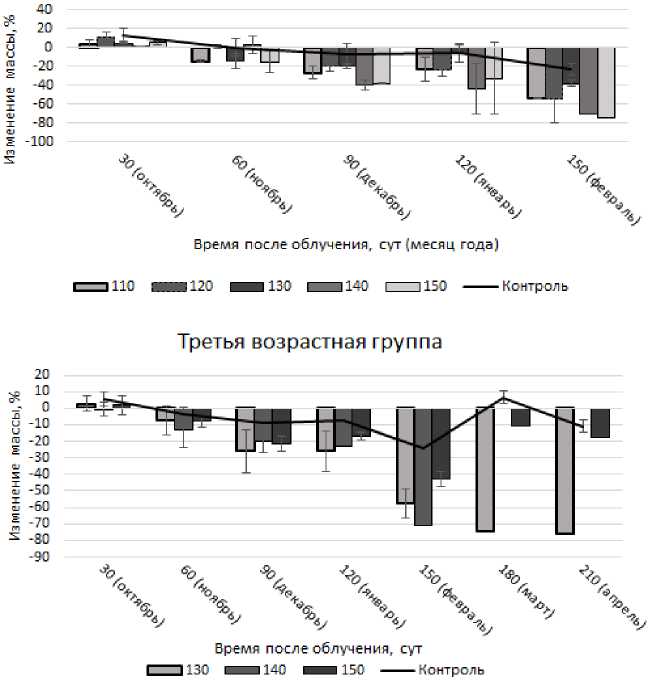
Рис. 3. Изменение массы моллюсков при облучении дозами второго диапазона.
В этот период масса моллюсков второй возрастной группы достоверно (p<0,05) снижается в 4,1 раза, а масса третьей группы – в 3,2 раза относительно контроля. Через 150 сут после облучения масса моллюсков возвращается к значениям массы контрольной группы (p<0,05). Дозы облучения третьего диапазона приводят к снижению массы (p<0,05) моллюсков через 90 сут (рис. 4), что соответствует периоду начала анабиоза. При этом масса моллюсков второй возрастной группы в среднем меньше контроля в 26,9 раза, третьей группы – в 3,3 раза.
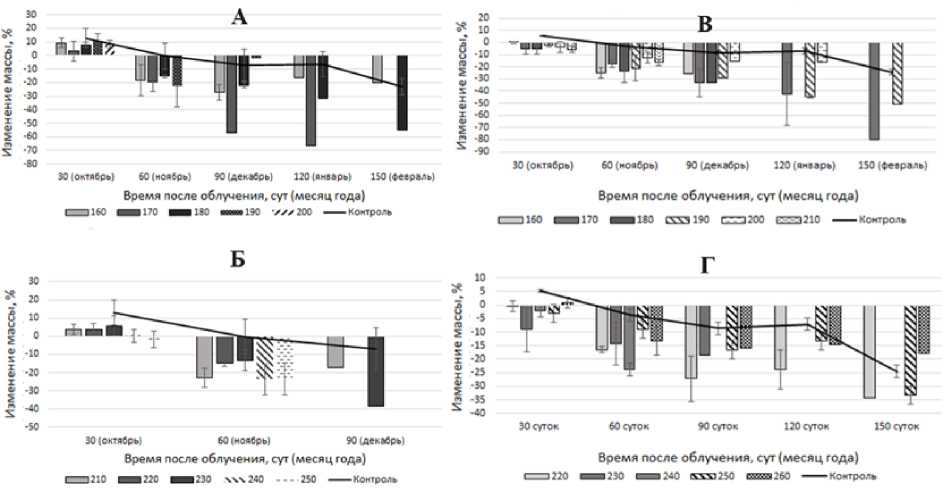
Рис. 4. Изменение массы моллюсков второй (А, Б) и третьей (В, Г) возрастных групп при облучении дозами третьего диапазона.
При облучении моллюсков дозами четвёртого диапазона отмечается снижение массы через 60 сут после облучения (рис. 5). При этом к моменту достижения 100% смертности масса второй возрастной группы животных уменьшилась в 52,3 раза, третьей группы – в 3 раза относительно контроля (p<0,05).
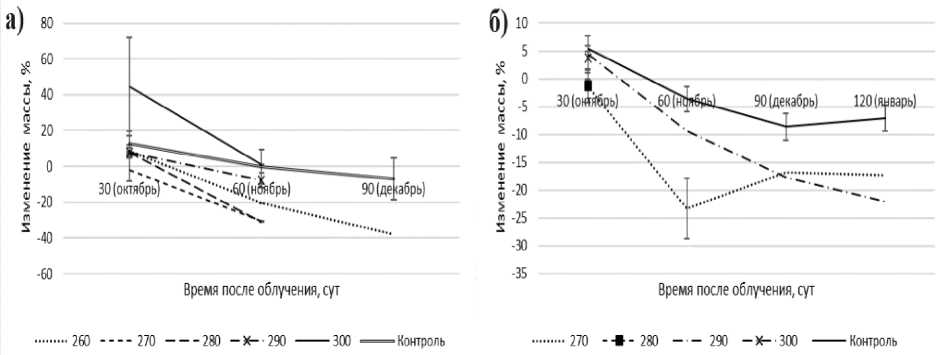
Рис. 5. Изменение массы моллюсков второй (а) и третьей (б) возрастных групп при облучении дозами четвёртого диапазона.
При облучении дозами свыше 260 Гр через 60 сут и 30-120 сут наступает 100%-ая смертность моллюсков второй и третьей возрастных групп соответственно. При этом масса моллюсков второй и третьей возрастных групп снижается с момента облучения до времени наступления абсолютной смертности в 2 раза и в 4 раза соответственно.
При обработке данных эксперимента также выявлены закономерности динамики массы жи-
вотных двух возрастных групп, учитывая установленные ранее три дозовых диапазона изменения показателя смертности моллюсков через 210 сут после облучения [6]. При этом первый дозонезависимый диапазон (10-70 Гр) характеризуется выживаемостью более 50% особей моллюсков. Второй дозозависимый диапазон (80-120 Гр и 80-140 Гр для второй и третьей возрастных групп соответственно характеризуется 50%-ой смертностью моллюсков. Третий дозонезависимый диапазон 130-300 Гр (для второй группы) и 150-300 Гр (для третьей группы) соответствует абсолютной смертности моллюсков.
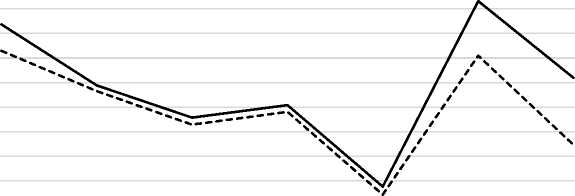
При облучении моллюсков дозами в диапазоне от 10 до 70 Гр (рис. 6) отмечается изменение массы моллюсков двух возрастных групп. До 150 сут после облучения различие в изучаемом показателе между двумя возрастными группами составляет в среднем 2,3%. Начиная со 150 сут эксперимента и до его окончания, изменения масса моллюсков второй группы на 11-13% больше, чем в третьей группе.
-5
-10
-20
-30
60 90
(ноябрь) (декабрь)
120 150 180 (март) 210
(январь) (февраль) (апрель)
30 (октябрь)
Время после облучения, сут (месяц года)
Третья возрастная группа
Вторая возрастная группа
Рис. 6. Изменение массы моллюсков при облучении дозами первого диапазона.
Облучение моллюсков дозами второго диапазона (рис. 7а) приводит к снижению массы особей второй и третьей возрастных групп на 21% и 23% соответственно, в период с начала эксперимента и до 150 сут. При этом изменение массы второй возрастной группы на 12,8% меньше, чем третьей. Начиная со 150 сут и до окончания эксперимента, это процентное соотношение увеличивается до 22%.
Облучение моллюсков дозами третьего диапазона (рис. 7б) приводит к 100% смертности к 150 сут эксперимента. Первоначальная масса моллюсков второй возрастной группы снижается на 51,6%, а третьей - на 43,3%. При этом в период с 60 сут отмечается изменение тенденции снижения массы моллюсков: масса второй группы уменьшается интенсивнее, чем третьей на 8%.
Установлены особенности радиационно-индуцированного изменения массы моллюсков. При этом облучение моллюсков двух возрастных групп дозами ниже 100 и 120 Гр не приводит к изменению массы, учитывая значимое отличие от контроля (p<0,05). Это может быть связанно с отсутствием необратимых изменений в организме животных, а также из-за достаточной выработ- ки мелатонина в период анабиоза, что может обеспечивать радиорезистентность в отдалённый период после облучения [14]. Такова рабочая гипотеза данных исследований, что требует дополнительных экспериментов с включением цикла работ по определению уровня мелатонина. Следует отметить, что прирост массы после анабиоза во второй возрастной группе выше, чем в третьей возрастной группе. При этом известно, что с возрастом происходит снижение уровня эндогенного мелатонина и, соответственно, устойчивость более молодых особей к облучению будет выше [15].
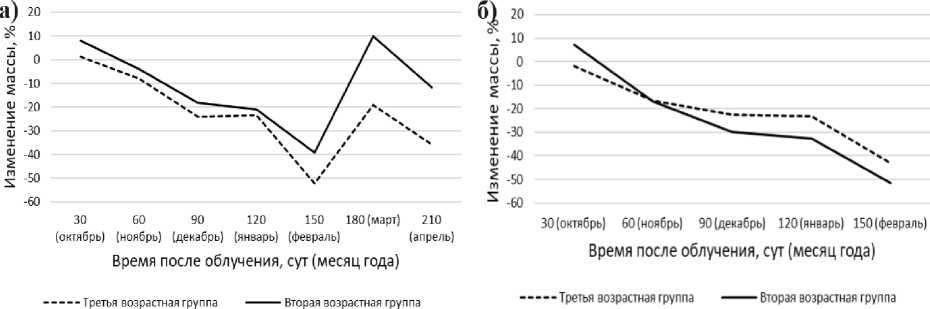
Рис. 7. Изменение массы моллюсков при облучении дозами второго (а) и третьего (б) диапазонов.
Облучение моллюсков дозами 110-150 Гр (вторая возрастная группа) и 120-150 Гр (третья возрастная группа) приводит к снижению массы (через 60-150 дней после облучения), что приходится на период анабиоза животных. В данный период у моллюсков отмечается снижение жизненных функций, а также возможно происходит снижение регенеративных способностей [14]. При этом отмечается более интенсивное снижение массы моллюсков второй возрастной группы. Однако данная тенденция меняется при выходе животных из анабиоза и наблюдается более интенсивное увеличение массы второй группы по сравнению с третьей возрастной группой. В исследовании [16] отмечено, что с увеличением возраста меняется процесс адаптивного ответа на различные факторы и происходят «необратимые» изменения в регуляции метаболизма, которые закрепляются в эпигенотипе. При этом в работе [17] установлено, что высокие дозы облучения приводят к элиминации менее устойчивых особей моллюсков L. Stagnalis в первые годы после аварии на Чернобыльской АЭС, однако, выжившая часть популяции вышла на более высокий уровень резистентности.
Дозы облучения моллюсков 150 Гр и выше приводят к снижению массы моллюсков двух возрастных групп на протяжении всего периода исследования. Подобные радиационно-индуци-руемые эффекты у животных наблюдаются многими исследователями. Снижение массы характерно для острой лучевой болезни, что, например, отражено в работе [18]: масса крыс снижается на 3,5-9% в течение 14 дней после острого облучения. В исследовании [19] γ -облучение мышей в дозах 9,5-10,0 Гр вызывало снижением массы тела на 24%. Стоит отметить, что снижение массы моллюсков второй возрастной группы происходит более интенсивно, чем третьей, что согласуется с данными об увеличении радиорезистентности с возрастом [20].
Заключение
Полученные научно обоснованные результаты лабораторного эксперимента расширяют базы немногочисленных (в отличие от водных моллюсков) данных о радиационно-индуцируемых эффектах представителей наземной малакофауны, что в свою очередь, вносит вклад в решение задач, сформулированных МКРЗ для развития экоцентрической концепции радиационной защиты. При этом выявлены дозовые диапазоны острого γ -облучения наземного моллюска с различным проявлением изменения биомассы животных двух возрастных групп. Дозы облучения моллюсков второй и третьей возрастных групп ниже 100 и 120 Гр соответственно не индуцируют изменение изучаемого показателя.
При облучении моллюсков дозами 110-150 Гр и 120-50 Гр (второй и третьей группы соответственно) установлено снижение массы животных в период анабиоза. Следует отметить, что масса необлучённых животных также снижается, однако, уменьшение массы облучённых моллюсков происходит интенсивнее в 4,1 раза (вторая возрастная группа) и в 3,2 раза (третья возрастная группа), чем в контроле.
Дозы облучения 150-200 Гр приводят к снижению массы моллюсков обеих возрастных групп, начиная с 90 сут после облучения, а дозы облучения свыше 200 Гр приводят к снижению, начиная с 30 сут. При этом у моллюсков второй возрастной группы наблюдается более интенсивное снижение массы (в среднем на 8%), чем у особей третьей группы в период с 60 сут эксперимента, что свидетельствует об увеличении радиорезистентности с возрастом. Однако, в отдалённый период после облучения (через 180 дней при облучении дозами ниже 120 Гр) у моллюсков второй возрастной группы отмечается увеличение массы, что может быть связано с более активным восстановлением после облучения.
Следует также отметить, что полученные экспериментальные результаты с учётом уже накопленных данных о воздействии острого и хронического γ - и β -облучения на релевантные показатели F. fruticum M. расширяют возможности рассмотрения моллюска как претендента на включение в набор RAPs.
Список литературы Радиационно-индуцируемое изменение массы наземного моллюска F. fruticum M. двух возрастных групп
- ICRP, 2008. Environmental protection: the concept and use of reference animals and plants. ICRP Publication 108 //Ann. ICRP. 2008. V. 38, N 4-6. P. 1-234.
- ICRP, 2009. Environmental protection: transfer parameters for reference animals and plants. ICRP Publication 114 //Ann. ICRP. 2009. V. 39, N 16. P. 1-111.
- ICRP, 2014. Protection of the environment under different exposure situations. ICRP Publication 124 //Ann. ICRP. 2014. V. 43, N 1. P. 1-59.
- Гудков Д.И., Назаров А.Б., Дзюбенко Е.В., Каглян А.Е., Кленус В.Г. Радиоэкологические исследования пресноводных моллюсков в Чернобыльской зоне отчуждения //Радиационная биология. Радиоэкология. 2009. Т. 49, № 6. С. 703-713.
- Лаврентьева Г.В., Черкасова Е.Е., Мирзеабасов О.А., Шошина Р.Р., Сынзыныс Б.И. Линейно-по-роговые эффекты гамма-облучения наземного моллюска F. fruticum M. третьей возрастной группы //Радиация и риск. 2023. Т. 32, № 3. С. 84-96.
- Черкасова Е.Е., Лаврентьева Г.В., Сынзыныс Б.И., Мирзеабасов О.А., Павлов А.Н. Влияние -облучения на смертность и биомассу наземного моллюска F. fruticum M. //Радиационная биология. Радиоэкология. 2023. T. 63, № 3. С. 285-299.
- Гребенников М.Е., Хохуткин И.М. Содержание тяжёлых металлов в наземных моллюсках в районе Среднеуральского медеплавильного завода //Экологические основы стабильного развития Прикамья: материалы научно-практической конференции. Пермь, 2000. С. 141-144.
- Сергеева Н.Г., Заика В.Е., Михайлова Т.В. Питание гребневика Mnemiopsis mccradyi в условиях Чёр-ного моря //Экология моря. 1990. Т. 35. С. 18-22.
- Светлакова Е.В., Коломысова Н.М., Скакунов Д.С. Биологические особенности, экология и значение виноградной улитки //Лучшая научная статья 2019: сборник статей XXV Международного научно-ис-следовательского конкурса, 30 мая 2019 г. Пенза: Наука и Просвещение, 2019. С. 26-29.
- Резник Е.П., Попов В.Н. Разведение улиток Helix lucorum и Helix albescens (Helicidae) в условиях фермерского хозяйства в Крыму //Экосистемы, их оптимизация и охрана. 2010. № 2(21). С. 153-157.
- Снегин Э.А., Тищенко А.Ю. Многолетняя динамика морфогенетических показателей наземного моллюска Cepaea vindobonensis (Gastropoda, Pulmonata, Helicidae) в памятнике природы «Бекарюковский бор» (Россия) //Заповедная наука. 2021. № 6(3). С. 58-72.
- Козминский E.В. Сезонная динамика размножения и репродуктивные показатели Bithynia tentaculata (Gastropoda, Prosobranchia) //Зоологический журнал. 2003 Т. 82, № 3. С. 325-331.
- Kuźnik-Kowalska E., Lewandowska M., Pokryszko B.M., Proćków M. Reproduction, growth and circadian activity of the snail Bradybaena fruticum (O.F. Müller, 1774) (Gastropoda: Pulmonata: Bradybaenidae) in the laboratory //Cent. Eur. J. Biol. 2013. V. 8, N 7. P. 693-700.
- Темурьянц Н.А., Демцун Н.А., Костюк А.С., Ярмолюк Н.С. Особенности регенерации планарий Dugesia tigrina и ноцицепции моллюсков Helix albescens в условиях слабого электромагнитного экранирования //Геофизические процессы и биосфера. 2011. Т. 10, № 4. С. 66-80.
- Анисимов В.Н. Эпифиз, биоритмы и старение организма //Успехи физиологических наук. 2008. Т. 39, № 4. С. 40-65.
- Божков А.И., Длубовская В.Л., Малеев В.А., Белецкая Л.Б., Дмитриев Ю.В. Сдерживающая рост диета вызывает различные стратегии адаптации организма у молодых и взрослых животных //Успехи геронтологии. 2006. № 19. С. 36-43.
- Голубев А.П. Динамика процессов радиоадаптации в популяциях моллюсков из водоемов Белорусского сектора зоны загрязнения ЧАЭС //Экологический вестник. 2012. № 2. С. 44-57.
- Дежаткина С.В., Тушина А.Д. Видовые особенности лучевой болезни животных //Инновационная деятельность в модернизации АПК: материалы Международной научно-практической конференции студентов, аспирантов и молодых ученых. В 3 частях, Курск, 7-9 декабря 2016 г. Часть 3. Курск: Курская государственная сельскохозяйственная академия им. проф. И.И. Иванова, 2017. С. 114-117.
- Васин М.В., Ушаков И.Б., Ковтун В.Ю., Семенова Л.А., Королева Л.В., Галкин А.А., Афанасьев Р.В. Терапевтический эффект длительного применения мелатонина на течение и летальный исход экспериментальной острой лучевой болезни //Бюллетень экспериментальной биологии и медицины. 2013. Т. 156, № 12. С. 738-740.
- Лазаревич Н.В., Сергеева И.И., Лазаревич С.С. Радиобиология: курс лекций. В 4 ч. Ч. 3. Радиобиология животных и человека. Горки: БГСХА, 2012. 103 с.
