Радиальный прирост желтопыльниковой и краснопыльниковой форм сосны (Pinus sylvestris L.) в условиях избыточного увлажнения почв северной тайги
 в условиях избыточного увлажнения почв северной тайги")
Автор: Тарханов Сергей Николаевич, Пинаевская Екатерина Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Лесные ресурсы
Статья в выпуске: 6-1 т.17, 2015 года.
Бесплатный доступ
Изучена изменчивость радиального прироста у желтопыльниковой и краснопыльниковой форм сосны в сосняках кустарничково-сфагновых на болотных верховых почвах северной тайги. Выявлено преимущество в средней величине годичного прироста в спелом возрасте у сосны с желтой окраской пыльников по сравнению с краснопыльниковой сосной. Проанализирована временная динамика ширины годичных колец у этих форм.
Сосна, желтопыльниковая и краснопыльниковая формы, радиальный прирост, избыточное увлажнение, почва, северная тайга
Короткий адрес: https://sciup.org/148204232
IDR: 148204232 | УДК: 631.524:582.475.4
Radial growth of F (var) sulfuranthera Kozubow and F. (var.) erythranthera sanio pine forms (Pinus sylvestris L.) in the conditions of northern taiga soils excess moistening
Variability of radial growth of (f. (var.) sulfuranthera Kozubow) and (f. (var.) erythranthera Sanio) forms of pine in pine forests of shrub-sphagnum bog on upland soils of northern taiga is studied. Advantage in the average size of a year growth at ripe age of pine with yellow coloring of boots in comparison with red coloring of boots at pine is revealed. Temporary dynamics of width of year rings at these forms is analyzed.
Текст научной статьи Радиальный прирост желтопыльниковой и краснопыльниковой форм сосны (Pinus sylvestris L.) в условиях избыточного увлажнения почв северной тайги
Проблема изучения радиального прироста как индикатора продуктивности хвойных насаждений при экологическом мониторинге включает вопросы изменчивости этого показателя под влиянием внешних и внутренних факторов [2, 6, 13, 14]. В пределах своего ареала сосна обыкновенная произрастает в различных экологических условиях и различается по морфологическим формам [9]. Радиальный прирост древесных видов зависит как от естественных и антропогенных факторов среды, так и обусловлен наследственными свойствами [1, 10, 16]. Ширина годичных колец изменяется под влиянием экологических факторов, но деревья разных морфологических форм имеют и генетические различия, которые в той или иной степени оказывают влияние на динамику радиального прироста. Изменчивость радиального прироста морфологических форм сосны на Европейском Севере изучена слабо. Известны лишь работы [15, 16], касающиеся радиального роста ширококронной и узкокронной форм сосны.
Окраска микростробилов у деревьев сосны является устойчивым генетическим признаком, который не изменяется в период онтогенеза. По окраске мужских стробилов выделяют формы: желтопыльниковая (f. (var.) sulfuranthera Kozu-bow) и краснопыльниковая (f. (var.) erythranthera Sanio) [3-5]. Краснопыльниковая форма отличается повышенным содержанием антоцианов, что способствует лучшему прогреванию ее солнечными лучами. Это явление особенно важно для сосен, произрастающих в
экстремальных условиях [4]. В сосняках сфагновой группы форма отличается повышенным содержанием антоцианов, что способствует лучшему прогреванию ее солнечными лучами. Это явление особенно важно для сосен, произрастающих в экстремальных условиях [4]. В сосняках сфагновой группы долевое участие краснопыльниковой формы может быть сопоставимо с численностью желтопыльниковой формы. С улучшением условий произрастания частота встречаемости в насаждениях краснопыльниковой формы снижается до 10 – 20 % от общего числа деревьев, а желтопыльниковой – увеличивается до 80 – 90 %. Краснопыльниковой сосне присущи: куполообразная крона рыхлого сложения, деформированность ствола и кроны, часто приобретающих уродливую форму, с сильно выдающимися толстыми, длинными и сильно искривленными сучьями, ажурность. Особенно это характерно для насаждений сфагновой группы типов леса, произрастающих на верховых торфяных почвах [11, 12]. В сосняках сфагновых желтопыльниковая сосна чаще имеет конусообразную, узкоовальную или с неопределенными геометрическими пропорциями крону с предельной продолжительностью жизни хвои от 3 до 8 лет [11, 12].
Цель исследований: выявление различий в динамике радиального прироста у морфологических форм сосны обыкновенной, различающихся окраской микростробилов (пыльников) на избыточно-увлажненных почвах северной тайги.
Методика исследования. Исследования проводили в низкопродуктивных (Va класса бонитета), низкополнотных (0,4-0,5) и чистых по составу сосняках кустарничково-сфагновых, произрастающих на болотных верховых почвах устья Северной Двины (северная тайга), на 3 постоянных пробных площадях, заложенных стандартными методами (ОСТ 56-69-83) [8]. Был проведен отбор кернов древесины ствола в направлениях с севера на юг и с востока на запад приростным буравом (на высоте 1,3 м) у форм деревьев с разным цветом пыльников. На каждой пробной площади объем выборки составлял 24 дерева каждой формы. Исходными данными для дендрохронологического анализа радиального прироста различных форм сосны послужили средние значения прироста, рассчитанные по измерениям ширины годичных слоев в двух взаимно перпендикулярных направлениях. Измерения ширины годичных слоев проводили методом световой микроскопии с помощью окуляр-микрометра с точностью ±0,05 мм [17]. Колебание длины полученных рядов прироста не превышали одного класса возраста. Выявление закономерностей радиального прироста в полученных временных рядах осуществляли дифференцированно для разных форм. Методом 5-летнего скользящего сглаживания определены относительные индексы прироста [1]:
1= i ф / i сгл • 100 %, (1)
где I - относительный индекс, %; i ф - фактическая ширина годичного кольца; i^ - сглаженная ширина годичного кольца (или норма прироста данного года рассчитанная любым конкретным способом). Данный способ расчетов обеспечивает удаление возрастного тренда.
Рассчитывали показатель чувствительности дерева (%) к условиям внешней среды по формуле [1]:
K s = 5! М / 5 i ; • 100 %, (2)
где 2! Ai I — абсолютная разность ширины соседних годичных колец деревьев по модулю; 2 i — сумма годичных колец деревьев за весь исследуемый период.
Уровни изменчивости оценивали по шкале С.А. Мамаева [7]. При сравнении одноименных признаков использовали t -критерий Стьюдента. Для выявления цикличности радиального прироста разных форм сосны использовали календарные даты наступления экстремальных приростов и значений нормы годичного прироста, рассчитанных по 5-летним скользящим кривым.
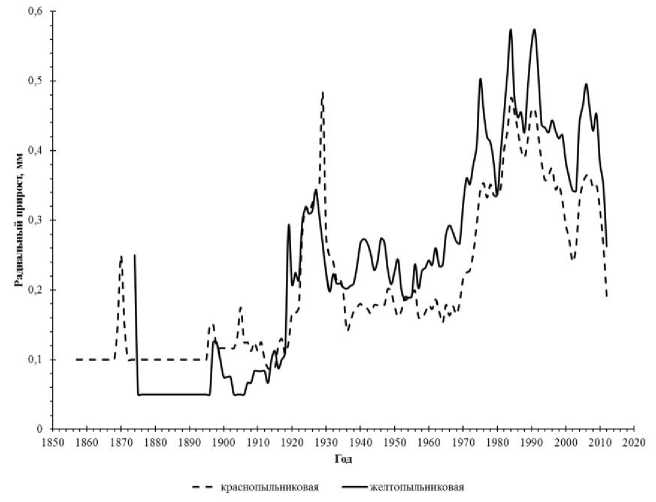
Рис. 1. Хронологическая динамика средних значений радиального прироста у форм сосны
Экспериментальная часть, анализ, обобщение и разъяснение полученных данных. Анализ полученных данных показал, что среднее значение годичного радиального прироста у сосны с красным цветом пыльников ниже (0,38±0,03 мм), чем у желтопыльниковой формы (0,50±0,04 мм). Эти различия достоверны
(t-критерий, р<0.05). Средние значения годичного прироста у отдельных деревьев краснопыльниковой формы колеблются от 0.12 до 0.86 мм, а желтопыльниковой от 0,16 до 1,13 мм. Временная динамика средних значений радиального годичного прироста показала, что в более молодом возрасте (40-60 лет) преимущественно в интенсивности роста имеет краснопыльниковая форма, а в 60-80 лет их показатели выравниваются. В старшем возрасте (80 и более лет) желтопыльниковая форма опережает в росте по диаметру ствола краснопыльниковую (рис. 1). К возрасту спелости, деревья с желтыми пыльниками формируют более широкие годичные кольца. Подобные различия во временной динамике ширины годичного кольца были выявлены нами ранее у деревьев с различной шириной кроны [15, 16]. Одновременно с осцили-рующими колебаниями радиального прироста, связанными с изменениями погодных условий, имеют место длинноволновые трендовые колебания, связанные с изменением экологических условий [2]. Это положение правоверно и для выборок деревьев сосны с разным цветом пыльников.
Для выборок деревьев каждой формы определены значения коэффициентов вариации и уровень хронологической изменчивости радиального прироста. Согласно эмпирической шкале С.А. Мамаева (1970) эти показатели (в пределах дерева) для обеих форм близки, и характеризуются очень высоким уровнем (C.V. =47-49%). Показатели чувствительности деревьев к воздействию внешней среды на болотных верховых почвах у желтопыльниковой и краснопыльниковой форм сосны также близки (9-10%). По Т.Т. Битвинскасу [1], при значении этого показателя более 25% деревья считаются чутко реагирующими на изменения условий внешней среды. Можно сделать вывод о слабой чувствительности рассматриваемых форм к изменениям внешних факторов при произрастании сосны в условиях постоянного избыточного увлажнения почвы. По-видимому, в неблагоприятных для роста условиях реакции деревьев у разных форм сосны на избыточное увлажнение почвы, вызывающее их корневую гипоксию и гипотермию, особенно в весенний и раннелетний периоды, сходны. Вероятно, деревья, которым в экстремальных условиях свойственен так называемый «стрессовый метаболизм», слабо чувствительны к воздействию других факторов (климатических и др.). Ранее [11], было установлено сходство и общий характер биохимических процессов у этих форм в стрессовых условиях.
Анализ колебаний индекса прироста для разных форм сосны показал, что среднее значение колеблется в пределах 102,5-103%. Для краснопыльниковой формы минимальное значение индекса прироста составляет 71%, а максимальное - 167%. Желтопыльниковая форма имеет более низкое минимальное значение (56%), но значительно превосходит краснопыльниковую форму по максимальному значению (357%). В целом амплитуда индекса прироста у разных форм высокая, а их колебания довольно синхронны. Наибольшая амплитуда колебаний индекса радиального прироста у желтопыльниковой и краснопыльниковой форм наблюдается в молодом возрасте (до 40 лет). По-видимому, на ранних стадиях онтогенеза деревьям свойственна более высокая чувствительность к действию внешних факторов, влияющих на камбиальную активность (рис. 2).
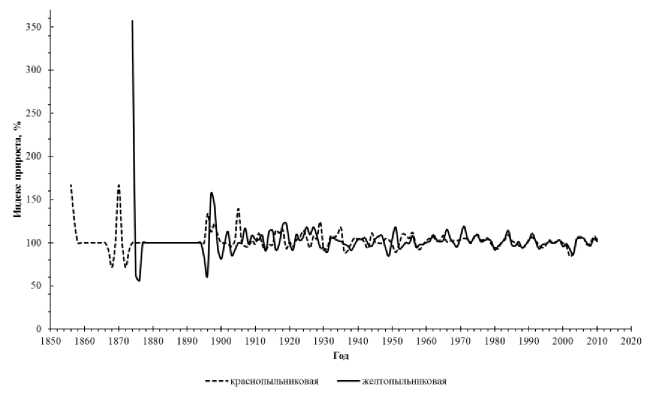
Рис. 2. Хронологическая динамика средних значений индексов прироста у форм сосны
При анализе динамики ширины годичных колец сосны выявлено несколько типов возрастных кривых годичного прироста. Для разных морфологических форм отсутствует четко выраженная кривая «большого роста» в динамике годичного прироста. Для большинства деревьев различных форм динамика годичного прироста характеризуется выраженными кривыми, когда
величина радиального прироста увеличивается с возрастом. Для меньшего количества деревьев выявлены равномерные кривые и кривые «большого роста», когда прирост максимален в молодом возрасте. Для краснопыльниковой формы сосны в процентном соотношении преобладают деревья с кривой увеличения прироста в «зрелом» возрасте (40%), для 32% деревьев характерна кривая «большого роста» и 28% деревьев характеризуются равномерным ростом (рис. 3). Для желтопыльниковой формы частота встречаемости деревьев с кривой увеличения прироста в «зрелом» возрасте больше (52%), по сравнению с краснопыльниковой формой (40%). Кривые «большого роста» и равномерная кривая для этой формы представлены примерно для одинакового числа деревьев (24-25%) (рис. 3).
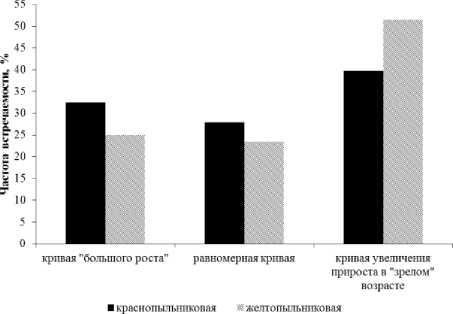
Рис. 3. Процентное соотношение деревьев с разными типами роста у форм сосны
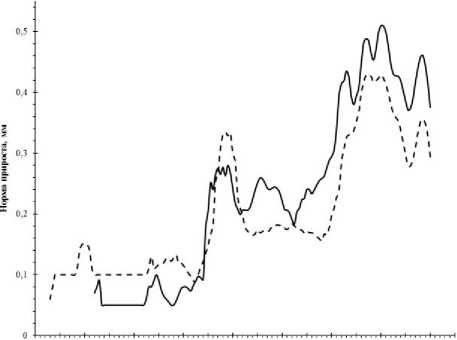
1850 1860 1870 1880 1890 1900 1910 1920 1930 1940 1950 1960 1970 1980 1990 2000 2010 2020
--краснопыльниковая ----желтопыльниковая
Рис. 4. Цикличность в динамике радиального прироста у разных форм сосны
Цикличность в колебаниях ширины годичных колец разных форм сосны изучена по кривым «нормы прироста». Средний период цикла для желтопыльниковой формы сосны между максимальными пиками прироста составил 13,7, а между минимальными показателями прироста - 13,9 года. Средний период цикла для краснопыльниковой формы между максимальными пиками прироста составил 8,7 лет, а между минимальными показателями прироста 8,5 лет (рис. 4).
Выводы: в условиях длительного избыточного увлажнения почв в северной тайге сосна с желтым цветом пыльников имеет более высокие средние значения абсолютных величин годичного радиального прироста по сравнению с краснопыльниковой формой ( t -критерий, p <0,05). Изменчивость годичного прироста сосны (в пределах дерева) соответствует очень высокому уровню ( C.V. =47-49%) как в выборке краснопыльниковой, так и желтопыльниковой форм. Амплитуда индексов прироста у разных форм высокая (102,5-103%), а их колебания довольно синхронны, что свидетельствует о сходстве их реакций на воздействие факторов внешней среды. В динамике ширины годичных колец у этих форм сосны выявлено несколько типов возрастных кривых годичного прироста и отсутствие четко выраженной кривой «большого роста». Большинство деревьев красно- и желтопыльниковой форм характеризуются кривыми, когда величина радиального прироста увеличивается с возрастом. Камбиальная активность в условиях постоянно избыточного увлажнения почв у деревьев краснопыльниковой формы выше, чем у деревьев с желтым цветом пыльников в более молодом возрасте (40-60 лет). В 6080 лет показатели их роста довольно сходны, что характерно для деревьев различных типов роста. В этот период в большей степени выражаются их наследственные особенности, реализованные и фиксированные во временном ряду. В старшем возрасте (более 80 лет) желтопыльниковая сосна по величине радиального прироста опережает краснопыльниковую форму. Таким образом, наиболее перспективной в хозяйственном отношении является желтопыльниковая форма сосны.
Работа выполнена при поддержке: ИЭПС УрО РАН в рамках темы ФНИР № 0410 – 2014 - 0025; Программы фундаментальных исследований УрО РАН, проект № 12-у-5-1005.
Список литературы Радиальный прирост желтопыльниковой и краснопыльниковой форм сосны (Pinus sylvestris L.) в условиях избыточного увлажнения почв северной тайги
- Битвинскас, Т.Т. Дендроклиматические исследования. -Л.: Гидрометеоиздат, 1974. 172 с.
- Ваганов, Е.А. Анализ роста дерева по структуре годичных колец/Е.А. Ваганов, И.А. Терсков. -Новосибирск: Наука, 1977. 150 с.
- Ефремов, С.П. Морфология и жизнеспособность пыльцы желто-и краснопыльниковой форм сосны обыкновенной на болотах и суходолах Западной Сибири/С.П. Ефремов, Т.С. Пименов, Т.С. Седельникова и др.//Хвойные бореальной зоны. 2010. № 1-2. С. 126-129.
- Козубов, Г.М. Внутривидовое разнообразие сосны обыкновенной (Pinus sylvestris L.) в Карелии и на Кольском полуострове: Автореф. дис. … канд. с.-х. наук. -Л., 1962. 15 с.
- Козубов, Г.М. О краснопыльниковой форме сосны обыкновенной//Ботан. журн. 1962 б.1. Т. 47, № 2. С. 276-278.
- Ловелиус, Б.В. Изменчивость прироста деревьев. -Л.: Наука, 1979. 232 с.
- Мамаев, С.А. Уровни изменчивости анатомо-морфологических признаков сосны//Ботанич. исслед. на Урале. Зап. Свердловск. отд-я Всес. бот. общ-ва. Вып. 5. Свердловск, 1970. С. 58-67.
- ОСТ 56-69-83 Площади пробные лесоустроительные. Метод закладки. ЦБ НТИГослесхоза, СССР, 1983. 14 с.
- Правдин, Л.Ф. Сосна обыкновенная. Изменчивость, внутривидовая систематика и селекция. -М.: Наука, 1964. 172 с.
- Румянцев, Д.Е. Влияние климатических факторов на рост сосны в южной Карелии//Лесоведение. 2004. № 5. С. 73-75.
- Тарханов, С.Н. Индивидуальная изменчивость биохимических признаков сосны обыкновенной в условиях аэротехногенного загрязнения//Лесоведение. 2011. № 1. С. 58-65.
- Тарханов, С.Н. Морфоструктурные особенности и изменчивость биохимических признаков форм Pinus sylvestris L. (Pinaceae) в условиях избыточного увлажнения почв северной тайги/С.Н. Тарханов, Е.А. Пинаевская, Ю.Е. Аншукова//Растительные ресурсы. 2014. Вып. 4. С. 63-74.
- Феклистов, П.А. Биологические и экологические особенности роста сосны в северной подзоне европейской тайги/П.А. Феклистов, В.Н. Евдокимов, В.М. Барзут. -Архангельск: Арх. гос. тех. ун-т, 1997. 140 с.
- Шиятов, С.Г. Дендрохронология верхней границы леса на Урале. -М.: Наука, 1986. 136 с.
- Щекалев, Р.В. Радиальный прирост и качество древесины сосны обыкновенной в условиях атмосферного загрязнения/Р.В. Щекалев, С.Н. Тарханов. -Екатеринбург: УрО РАН, 2006. 117 с.
- Щекалев, Р.В. Радиальный прирост сосны обыкновенной при аэротехногенном загрязнении в бассейне Северной Двины/Р.В. Щекалев, С.Н. Тарханов//Лесоведение. 2007. № 2. С. 45-50.
- Cook, E.R. A time series analysis approach to tree-ring standardization. -University of Arizona, Tucson. 1985. 171 p.
