Распределение и использование 14С-ассимилятов в растениях с различной морфологической и физиологической организацией донорно-акцепторной системы

Автор: Табаленкова Галина Николаевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-3 т.20, 2018 года.
Бесплатный доступ
Изучено распределение и использование 14С- фотоассимилятов у травянистых столонообразующих растений Stachys sieboldii Mig. и Ajuga reptans L. Установлено, что основной акцептирующей зоной S. sieboldii являются диатропные побеги, которые образуют мощные аттрагирующие центры, куда транспортируются продукты текущего фотосинтеза из листьев и ассимиляты из мест временного депонирования. Рост плагиотропных побегов A. reptans обеспечивают ассимиляты из материнской розетки. Показано, что для S. sieboldii и A. reptans характерна высокая скорость использования ассимилятов, что связано с процессами формирования подземных и надземных побегов и позволяет растениям в короткий период образовывать большие клоны.
Столонообразующие растения, 14с-ассимиляты, донорно-акцепторные отношения, стахис зибольда, живучка ползучая
Короткий адрес: https://sciup.org/148314085
IDR: 148314085 | УДК: 581.131
Distribution and using of 14C - assimilates in plants with different mophological and physiological organization of sink-source system
The distribution and use of 14C - photoassimilates in perennial herbaceous stoloniferous plants Stachys sieboldi and Ajuga reptans were studied. The general patterns of changes in sink- source relations during the vegetative growth were established. It was shown that the directionality of the carbon transport flows in S. sieboldii and A. reptans is related to the growth of underground and above-ground shoots of renewal. This contributes to the formation of large clones that causes high competitiveness and survival of species.
Текст научной статьи Распределение и использование 14С-ассимилятов в растениях с различной морфологической и физиологической организацией донорно-акцепторной системы
Работа выполнена в рамках темы «Физиология и стресс-устойчивость фотосинтеза растений и пойкилогидрических фотоавтотрофов в условиях Севера» (№ГР АААА - А17-117033010038-7)
Рост растений в значительной степени определяется обменом углерода, поскольку большую долю сухого вещества растительного организма составляет именно этот элемент. Одним из главных факторов, регулирующих фотосинтетическую функцию а, следовательно, определяющим рост растений являются взаимоотношения между производящими и потребляющими ассимиляты органами. У разных морфотипов растений направленность потоков фотоассими-лятов зависит от уровня их синтеза в листьях и от аттрагирующей активности отдельных органов, т.е. от донорно-акцепторных отношений. Концепция донорно-акцепторных связей во многом способствовала формированию представлений о растении как целостной системы. В процессе реализации морфогенетической программы роста и развития в растении происходит изменение соотношения экспортноимпортной функции органов, т.е. происходит перестройка донорно-акцепторных связей [11, 12]. Эти процессы, как правило, пространственно разделены, и роль связующего звена между ними выполняет транспорт [8]. Ассимиляты, образованные в процессе фотосинтеза, перемещаются к потребляющим тканям, которые связаны либо с активными ростовыми зонами или выполняют функцию запасающих органов. В ре зультате меняется интенсивность и направлен- Табаленкова Галина Николаевна, доктор биологических наук, ведущий научный сотрудник лаборатории экологической физиологии растений.
ность транспорта ассимилятов. Однако влияние эпигенеза на фотосинтез не является односторонним, а во многом определяется условиями внешней среды. Подавление фотосинтеза при неблагоприятных внешних условиях ограничивает процессы роста и отложения в запас [11, 12]. В случае ингибирования транспорта асси-милятов или процессов утилизации продуктов фотосинтеза в акцепторных зонах наблюдается метаболитная репрессия фотосинтеза и как результат накопление в хлоропластах глюкозы и крахмала [4, 8, 11, 17]. В зависимости от скорости утилизации продуктов фотосинтеза в акцепторных зонах изменяется градиент концентрации осмотически активных веществ между донором и акцептором [2,3]. В модели [16] показано, что дифференциальное развитие акцептирующих тканей зависит от общего пула резервов асси-милятов и скорости их утилизации. Чем быстрее используются ассимиляты в акцепторных зонах, тем выше вероятность их получения. Распределение и использование углерода в течение жизненного цикла исследовано в основном для культурных растений, у которых транспорт ас-симилятов направлен в хозяйственно ценные органы: колосья, клубни, корнеплоды [5,14, 9, 10]. В меньшей степени изучен этот вопрос для многолетних растений [13], особенно для видов, образующих большие клоны. Большую часть биомассы растений такой жизненной формы составляют надземные или подземные стелющиеся побеги, которые являются основным резервом углерода.
Цель работы: оценить вклад отдельных органов растений в ассимиляцию углерода, исследовать динамику распределения и использования углерода у столонообразующих растений стахиса Зибольди ( Stachys siboldii Mig.) и живучки ползучей ( Ajuga reptans L.) .
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили на растениях Stachys siboldii и Ajuga reptans , выращенных в условиях коллекционного питомника. Stachys siboldii ––– травянистое столонообразующее растение сем. Lamiaceae, размножается клубнями, которые формируются на столонах. В природе встречается в тропических и субтропических горных районах Китая. Как лекарственное и овощное растение выращивается в некоторых европейских странах, а также в Китае, Японии и Монголии.
Ajuga reptans ––– многолетнее травянистое растение со стелющимися и укореняющимися побегами сем. Lamiaceae – вид широко распространенный в Европе. В культуру введен в качестве декоративного почвопокровного растения [7]. Характерной чертой S. siboldii и A. reptans является хорошо выраженная способность к вегетативному размножению, которое у S. siboldii осуществляется за счет развития подземных диатропных, у A. reptans — надземных плагио-тропных побегов. В условиях Севера семенное размножение у S. siboldii практически отсутствует, а у A. reptans играет второстепенную роль.
Растения S. siеboldii выращивали из клубней при площади питания 50х50 см. Растения A. reptans выращивали из перезимовавших прикорневых розеток, которые высаживания на делянки конце апреля – начале мая. Об интенсивности ассимиляции углерода отдельными органами судили по их удельной радиоактивности после экспозиции растений в атмосфере с 14СО2. Растения S. siboldii экспонировали в атмосфере с 14СО2 в фазу бутонизации. Изучение ассимиляционной способности и распределение 14С у растений A. reptans проводили в два этапа: в первом опыте в атмосфере 14СО2 экспонировали растения, с плагиотропными побегами, во втором – молодые имматурные растения без побегов. Меченую углекислоту генерировали из Ва14СО3 при добавлении 0.1 н раствора HCl. Начальная концентрация СО2 в камере составляла 0.09%. Оба вида выдерживали в атмосфере 14СО2 в течение 20 мин. под прозрачной камерой. Пробы для анализа отбирали через 1 ч, 1, 3, 10, 30 и 40 суток после экспозиции. В каждую пробу входило 2 растения. Повторность 4-кратная. Растения разделяли по органам, фиксировали в сушильном шкафу при температуре 105oС, затем досушивали при 70oС. Удельную радиоактивность (УА) порошка определяли на альфа-бета радиометре УМФ-1500 Д (Россия) в 5-6 кратной повторности. Убыль радиоактивности из листьев в результате транспорта асси-милятов вычисляли с учетом затрат на дыхание согласно методике [4]. По величине УА и соотношению масс органов рассчитывали количество метки в растениях. Относительную радиоактивность органов рассчитывали как процент от общего количества 14С в растении и от первоначально ассимилированного количества углерода. У S. siboldii оценивали включение 14С в белки, свободные аминокислоты, сахара и остаточную фракцию [6,18]. Для этого каждую фракцию доводили до определенного объема, из которого 0.5-1 мл вносили во флаконы со сцинтилляционной жидкостью и просчитывали на счетчике «Бета-1» (Россия), повторность пяти-шестикрат-ная. Стандартная ошибка для радиологического анализа 5 -10%.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
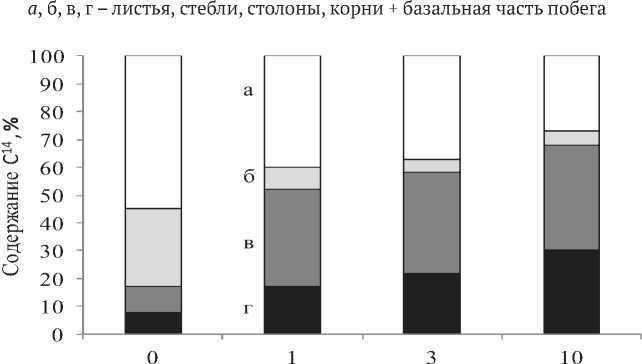
Ассимиляция 14СО2 и распределение 14С -ассимилятов в растениях S. siboldii . S. siboldii относится к растениям с высокой скоростью роста надземных и подземных органов. Ассимиляция 14СО2 происходит в основном листьями, на долю которых приходилось около 90% ассимилированного углерода. Несмотря на высокую долю стеблей, их активность в расчете на растение была довольно низкой и составляла всего около 7% от радиоактивности целого растения (рис.1), что значительно ниже, чем в злаковых травах [6,14]. Слабая ассимиляционная способность стеблей, по-видимому, обусловлена сильной опушенностью и низким содержанием в них зеленых пигментов. Из листьев 14С быстро транспортировался в подземную часть растений, где уже через 1 ч после экспозиции в атмосфере с 14СО2 отмечалось свыше 7% метки. Интенсивный отток ассимилятов в подземные органы в первые часы после экспозиции растений в атмосфере 14СО2 наблюдалось нами и для рапонти-ка сафлоровидного [14]. Спустя сутки около 10% 14С- ассимилятов оттекало из листьев в стебли и подземные органы S. siboldii. Через трое суток в листовых пластинках оставалось 50% ассимилированного углерода, около 30% находилось в стеблях и свыше 20% в подземных органах.
Через 30 суток при достаточно низкой удельной активности относительное содержание 14С в листьях составляло около 50% от целого растения (рис.1). Это связано с постоянным новообразованием ассимиляционной поверхности и свидетельствует о высокой аттрагирующей способности молодых листьев. В стеблях количество 14С - ассимилятов через 10 суток после экспозиции уменьшилось более чем в 3 раза
(рис.1). Следовательно, у S. siboldii продукты фотосинтеза не депонируются в стеблях, как показано для некоторых растений [1,15], а используются на формирование активно растущей надземной и подземной биомассы. Длинный световой день, характерный для условий Севера, способствовал образованию мощной надземной массы. Площадь листьев одного растения S. siboldii в фазу цветения составляла около 2 дм2. В этот же период начинался интенсивный рост подземных побегов и образование столонов, т.е. распределение 14С в системе целого растения в этот период обусловлено конкуренцией за ас-симиляты формирующихся столонов, и вновь образующихся надземных боковых побегов. Сокращение длины дня и понижение температуры воздуха в первой декаде августа приводило к торможению роста надземных органов и усилению аттрагирующей способности подземных побегов, где сосредотачивалось свыше 40% 14С, из них половина находилась в столонах (рис.1). Следовательно, с ростом столонов и началом формирования клубней изменялась направленность транспорта, ассимиляты, образованные в процессе фотосинтеза, перемещались к активно растущим запасающим органам.
Вк лючение 14С в метаболиты биомассы растений S. siboldii . Фракционирование позволяет получать информацию об участии 14С в синтезе и обновлении основных компонентов биомассы. На рисунке 2 представлены данные по включению и распределению 14С в метаболиты листьев. Как видно из рисунка, через 1 ч после экспозиции, основная часть (более 50%) ассимилированного листьями углерода находилась в сахарах, значительная доля была сосредоточена в свободных аминокислотах. Через сутки доля 14С в свободных аминокислотах снижалась более чем в 3 раза и возрастала в белках. Следовательно, основная часть аминокислот, образованных в процессе фотосинтеза, в первые сутки после экспозиции растений в атмосфере 14СО2 использовалась для синтеза белка (рис. 2).
На протяжении последующих суток доля 14С в аминокислотах и белках изменялась незначительно и составляла соответственно 5-8% и 35-38% от содержания 14С в листьях. Сразу после экспозиции, в сахарах сосредотачивалось свы-
100 -
90 -
80 -
70 -
60 -
50 -
40 -
30 -
20 -
10 -
0 -













г
а
б


в

0 1 3 10 30 40
Время от экспозиции в атмосфере 14СО2, сутки
Рис. 1. Распределение 14С в биомассе растений S. siboldii :
Время от экспозиции в атмосфере 14СО2 , сутки
Рис. 2. Распределение 14С во фракциях листьев S. siboldii :
а, б, в, г - сахара, аминокислоты, белки, остаточная фракция, % от содержания 14С в листьях
ше 50% 14С. Сахара активно транспортировались из листьев в акцепторные органы клубни. Через 10 сут. в листьях оставалось около 27% сахаров, значительная доля находилась в проводящих тканях - стеблях (37%) и в запасающих органах -клубнях (32%).Углерод постепенно включался в остаточную фракцию, состоящую в основном из структурных компонентов биомассы (деградирующие соединения белковой природы, структурные углеводы, клетчатка и т.п.) [18]. На 10 сут. доля 14С в остаточной фракции составляла до 30% от общего содержания в листьях (рис. 2).
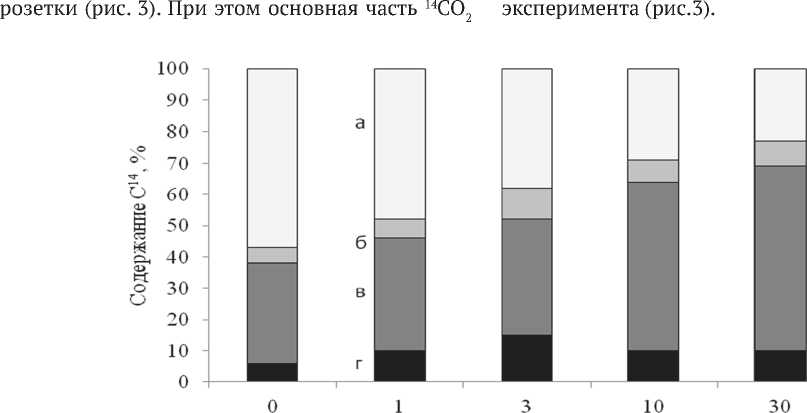
Ассимиляция 14СО2 и распределение 14С -ассимилятов в растениях A. reptans. При экспонировании в атмосфере с 14СО2 растений A. reptans (опыт 1) значительная доля около 50%, образовавшихся в процессе фотосинтеза асси-милятов, локализовалась в листьях материнской ассимилировалась зрелыми листьями, удельная активность которых в 2 раза выше, чем у старых (перезимовавших) и молодых листьев. Удельная активность листьев плагиотропных побегов была на порядок ниже, чем материнской розетки, что свидетельствует об их низкой ассимиляционной способности. Однако благодаря тому, что в среднем одно растение образует до 20 побегов, образующих значительную биомассу, на их долю приходилось до 30% ассимилированного углерода от целого растения (рис.3). Доля корней в растениях A. reptans не велика (корни/ побеги = 0.12), но их удельная активность сопоставима с удельной активностью листьев, что позволяет предположить наличие быстрого оттока ассимилятов в подземные органы. Доля 14С в корнях составляла 10- 15% от содержания 14С в целом растении и мало изменялась в течение
Время от экспозиции в атмосфере МСО2. сутки
Рис. 3. Распределение 14С в биомассе растений A. reptans (опыт 1): а, б, в, г - листья материнской розетки, соцветия, плагиотропные побеги (стебель+лист), корни, (доля от содержания 14С в целом растении, %)
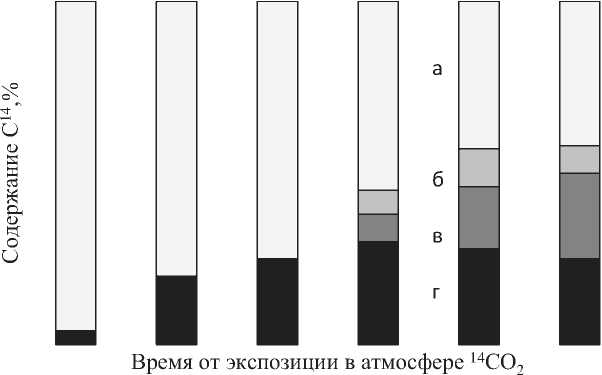
Рис. 4. Распределение 14С в биомассе растений A. reptans (опыт 2): а, б, в, г - листья материнской розетки, соцветия, плагиотропные побеги (стебель+лист), корни, (доля от содержания 14С в целом растении, %)
Время от экспозиции в атмосфере 14СО2, сутки
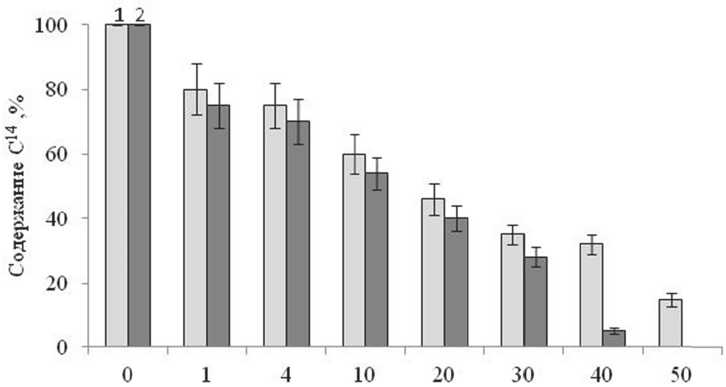
Рис. 5. Содержание 14С в биомассе растений A. reptans (1) и S. siboldii (2) после ассимиляции в атмосфере с 14СО2 (доля от первоначальной радиоактивности,%)
Плагиотропные (надземные) побеги в растениях A. reptans развиваются из почек в пазухах нижних хорошо развитых листьев розетки [7]. Для выяснения роли материнской розетки в их образовании, мы экспонировали в атмосфере с 14СО2 растения до начала формирования пла-гиотропных побегов (опыт 2). Согласно полученным данным, образование плагиотропных побегов происходит в основном за счет использования ассимилятов материнской розетки.
В пользу такого предположения говорит тот факт, что на протяжении 50 суток, удельная активность и доля С14 в материнской розетке постепенно снижалась, в корнях изменялась незначительно, а в плагиатропных побегах возрастала с 7 до 30% (рис.3). Следует отметить, что распределение С14 - ассимилятов по плаги-отропным побегам неравномерна, чем раньше был сформирован побег, тем выше его удельная активность, т.е. аттрагирующая способность этих побегов выше.
Полученные данные позволяют оценить во времени убыль 14С- ассимилятов из растений S. siboldii и A. reptans в течение вегетации. За сутки растения теряли около 20% 14С. В последующие 10 суток количество 14С снижалось и через месяц в растениях оставалось 30 -35% ассимилированного углерода. Через 40 дней ассимиляты практически полностью используются растениями S. siboldii , а в растениях A. reptans остается еще свыше 30% 14С от первоначальной радиоактивности (рис. 5).
Это объясняется тем, что для роста плагио-тропных побегов и формирования биомассы уA. reptans требуется значительно меньше ассими-лятов, чем для образования мощной биомассы у S. siboldii. Расчет показал [4], что за первые сутки на дыхание расходовалось до 16%, на транспорт около 12% ассимилированного углерода. Соотношение транспортных и дыхательных затрат составляло 0.8, т. е. доля радиоактивности приходящаяся на транспорт ассимилятов сопоставима с тратами на дыхание.
ЗАКЛЮЧЕНИЕ
Анализ распределения и использование ассимилированного углерода растениями Stachys siboldii и Ajuga reptans показал, что направленность транспортных потоков14С - ассмилятов связана с высокой акцептирующей активностью диатропных у S. siboldii и плагиотропных побегов у A.reptans. На их формирование используются продукты текущего фотосинтеза из листьев и ассимиляты из мест временного депонирования. Наличие высокой акцептирующей способности, вероятно связано с функциональной особенностью плагиотропных и диатроп-ных побегов, которые выполняют основную репродуктивную функцию. Высокая скорость роста вегетативных побегов плагиотропных у A. reptans и диатропных у S. siboldii обуславливает высокую конкурентоспособность и выживаемость видов при отсутствии семенного размножения и позволяет растениям в короткий период образовывать большие клоны.
Список литературы Распределение и использование 14С-ассимилятов в растениях с различной морфологической и физиологической организацией донорно-акцепторной системы
- Алиев Д.А. Транспорт и распределение 14С-ассимилятов у генотипов пшеницы, различных по фотосинтетическим признакам и урожайности / Д.А. Алиев, С.Х. Керимов, А.А. Джангиров, А.А. Ахмедов // Физиология растений. 1996. Т. 43, № 1. С. 57-61.
- Гамалей Ю.В. Транспортная система сосудистых растений.СПб.:Изд-во СПбГУ. 2004. 421 с.
- Гамалей Ю.В. Роль тонопласта клеток мезофилла в определении русла загрузки флоэмы. К 30-летию исследований загрузки флоэмы // Физиология растений. 2007. Т. 54. №1. С. 5-14.
- Глаголева Т.А., Чулановская М.В. Фотосинтетический метаболизм и транспорт ассимилятов у С4-галофитов Араратской долины // Физиология растений. 1996. Т. 43, № 3. С. 399-408.
- Гладун И.В., Карпов Е.А. Образование ассимилятов и их распределение после цветения между метелкой и вегетативными органами у риса // Физиология растений. 1993. Т. 40. № 5. С. 728-733.
