Распределение и наследование «A» типа оболочки семян в популяции brassica rapa l

Автор: Игнатов А.Н., Павлова М.Е., Терехин А.А., Хида Кен-Ичи
Журнал: Овощи России @vegetables
Рубрика: Современные направления селекции овощных культур
Статья в выпуске: 4 (25), 2014 года.
Бесплатный доступ
Оценивали частоту встречаемости и наследуемость «A» (ослизняющегося) и «B» (не слизистого) типов оболочки семян в популяциях B. rapa, происходящих из четырех географических регионов: Европы, Средней Азии, Китая и Японии. «A» тип оболочки семян встречался у растений среднеазиатской и японской субпопуляций этого вида, представленных торией, азиатской репой и группой B. rapa ssp. nipposinica (L.H.Bailey) Hanelt. В пределах внутривидовых скрещиваний «A» тип оболочки семян контролировался двумя комплементарно взаимодействующими доминантными генами, причем генотип зародыша семени оказывал влияние на фенотип материнской ткани оболочки.
Короткий адрес: https://sciup.org/14025125
IDR: 14025125 | УДК: 635.127:631.531.01
Geographic distribution and inheritance of «A» type seed coat in brassica rapa L. Population
Distribution and inheritance of «A» and «B» types of seed coat in populations of B. rapa originated from Europe, Central Asia, China and Japan was studied. «A» type seeds were found in Central Asian and Japanese populations represented by toria, Asian turnip and plants of «B». rapa ssp. nipposinica (L.H.Bailey) Hanelt. In intraspecific crosses, «A» type of seed coat was controlled by two complementary dominant genes.
Текст научной статьи Распределение и наследование «A» типа оболочки семян в популяции brassica rapa l
2 Российский университет дружбы народов
Россия, Москва, ул. Миклухо-Маклая,
3Национальный институт овощных, декоративных культур и чая, в настоящее время место работы –
Университет провинции Судзука, Япония
Ано-чо, Мие-кен, 514-2392, Япония
Оценивали частоту встречаемости и наследуемость «A» (ослизняющегося) и «B» (не слизистого) типов оболочки семян в популяциях B. rapa, происходящих из четырех географических регионов: Европы, Средней Азии, Китая и Японии. «A» тип оболочки семян встречался у растений среднеазиатской и японской субпопуляций этого вида, представленных торией, азиатской репой и группой B. rapa ssp. nipposinica (L.H.Bailey) Hanelt. В пределах внутривидовых скрещиваний «A» тип оболочки семян контролировался двумя комплементарно взаимодействующими доминантными генами, причем генотип зародыша семени оказывал влияние на фенотип материнской ткани оболочки.
Оболочка семени имеет ключевое значение для сохранения его жизнеспособности и осуществления функции прорастания в нужное время в оптимальных условиях для развития нового поколения растения. Свойства оболочки определяют устойчивость семени к биотическим и абиотическим стрессам [1]. Оболочка семени – ткань материнского растения, которая формируется из дифференцированных клеток интегумента семязачатка. Во время роста и развития эпи- дермальные клетки увеличивают толщину клеточной стенки и накапливают полисахариды в апопласте. Эти полисахариды способны накапливать значительное количество воды и увеличивать свой объем за счет гидратации, окружая семя вязкой слизистой оболочкой. Формирование слизи на семенах (миксоспермия) не носит систематического характера у покрытосеменных, но эта черта была отмечена у семян растений, по крайней мере, ста семейств [2, 3]. Процесс образования ослизняющейся оболочки семени хорошо изучен у резухи Таля (Arabidopsis thaliana) [1, 4, 5], что схематически показано на рисунке 1 [6].
Дифференциация тканей оболочки семени начинается после оплодотворения и заканчивается в момент начала созревания и накопления запасных веществ. Дифференциация начинается со значительного увеличения размера клеток и центральной вакуоли. Когда эмбрион входит в фазу глобулы, гранулы крахмала синтезируются в ци-
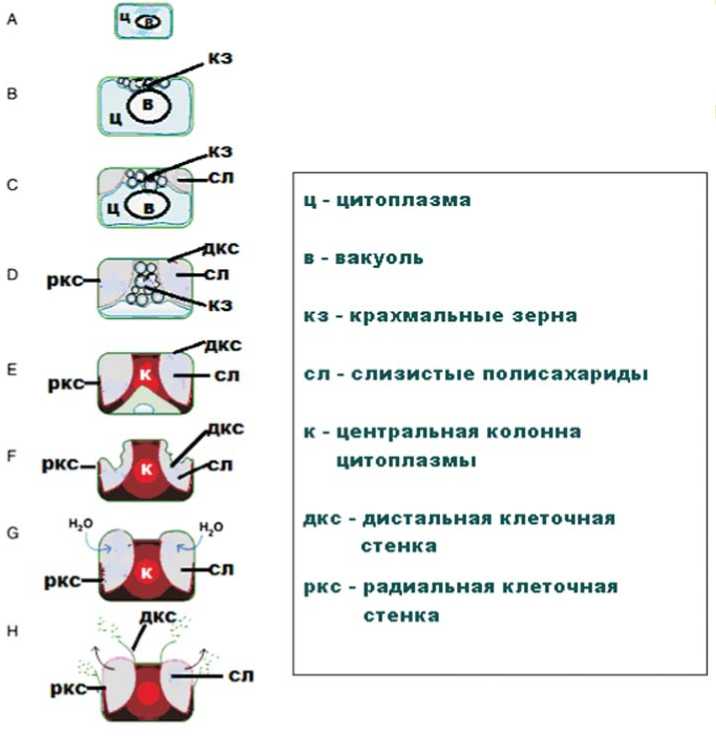
Рис. 1. Схематическое изображение дифференциации клеток оболочки семени резухи Таля [6].
-
(A) Первоначально, клетки имеют большую вакуоль, оттесняющую цитоплазму к краям клетки.
-
(B) В стадии сердечка формируются крахмальные зерна
-
(C) В стадии торпеды слизистые полисахариды накапливаются в апопласте.
-
(D) Центральная колонна цитоплазмы заполнена гранулами крахмала.
-
(E) Цитоплазматическая колонна заполнена материалом вторичной клеточной стенки, который также откладывается вдоль основания клетки и радиальной стенки клетки. Цитоплазма уменьшается, крахмальные зерна деградируют.
-
(F) Клетки отмирают к моменту созревания семени, и семя теряет воду за счет дегидратации слизистых полисахаридов, которые тонким слоем располагаются вокруг центральной колонны цитоплазмы.
-
(G) При увлажнении полисахариды набухают и давят на дистальную часть первичной клеточной стенки.
-
(H) Дистальная стенка клетки разрушается под давлением увеличившихся в объеме полисахаридов, которые выходят наружу, образуя слизистый слой на поверхности семени.
топлазме и накапливаются у дистального части клеточной стенки, по мере превращения ее в торпеду (рис. 1B). Этот процесс сопровождается отложением полисахаридов в апопласте на стыке между радиальными и дистальными клеточными стенками, и продолжается до тех пор, пока эмбрионы не достигнут в стадии изогнутой семядоли (рис. 1С). В конце развития семени гранулы крахмала деградируют, и вторичная клеточная стенка откладывается на границе цитоплазмы (рис. 1E).
У резухи Таля признак ослизняющейся оболочки присутствует не у всех растений, и определяется, по меньшей мере, одним геном, кодирующим фермент биосинтеза пектина (MUCILAGE-MODIFIED4) и несколькими транскрипционными факторами влияющими на него – APETALA2, TRANSPARENT TESTA GLABRA1, GLABRA2 [7], AtDOF4.2 и MYB5 [8], взаимодействующими с по крайней мере одним специфичным промотором в тканях оболочки семян [9].
Семена растений Brassica rapa L. (турнепс, тория, пак-чой и другие) также имеют два четко различимых типа оболочки – ослизняющийся и не слизистый [10]. Так называемые семена «A» типа имеют ясно различимую под микроскопом регулярную (ячеистую) структуру эпидермиса, способную быстро адсорбировать воду и набухать, создавая слой слизи на поверхности семян. Семена «B» типа с гладким эпидермисом не имеют набухающего при поглощении воды слоя. Среди других растений рода Brassica, «А» – тип оболочки семян встречается только среди европейской суб-популяции B. juncea Coss. [11] и среди дикорастущих видов B. barrelieri Janka, B. robertiana J.Gay, B. tornefortii Gouan и др. [12].
При изучении гибридных популяций японских реп, полученных от скрещивания сортов с «A» и «B» типами семян, было показано доминирование «A» типа. На основании этих результатов «A» тип семян был признан исходным или «диким» признаком, а «B» тип – производным [13]. При межвидовых скрещиваниях растений B. rapa «A» типа семян с B. juncea или B. carinata (Brown) ( «B» тип семян), этот признак наследовался строго по материнской линии [12]. Из-за вариации в пределах одного вида тип оболочки семян рассматривается как важный маркерный признак, полезный для таксономии и селекции овощных и масличных культур B. rapa. Целью данного исследования было оценить распределение этого признака среди коллекции азиатских и европейских образцов B. rapa и изучить особенности его наследования в пределах разнообразных популяций этого вида.
1. Происхождение образцов B. Rapa и тип оболочки семян
|
Образец |
Подвид |
Происхождение |
ТОС |
ЧАТ, % |
|
Swan, C-472, C-473, C-476, Oshita kanamachi, Kuronami, Sugulruna, Yori |
rapa |
ЯП |
A |
100.0 |
|
Shogoin oomaru |
- |
- |
AB |
69.0 |
|
Someya |
- |
- |
AB |
80.0 |
|
Green Globe, Purple Top, Leilainder, Milan white, Petrovskaya 01, ..., 12 |
- |
ЕВР |
B |
0 |
|
Сурепка, 01, ….23 |
campestris |
ЕВР |
B |
0 |
|
93-292, 94-85 |
- |
КИТ |
B |
0 |
|
Siroguki Sensuji |
nipposinica |
ЯП |
A |
100.0 |
|
Mizuna 94-210 |
- |
ЯП |
AB |
76.0 |
|
Mizuna 91-155, 85-502 |
- |
- |
B |
0 |
|
Tooka |
- |
- |
AB |
67.0 |
|
Wase nagasaki, Rijusai, 91-5, 87-410 |
narinosa |
КИТ |
B |
0 |
|
Xi fu, Osaka Shirona, Shinajuma, Tango Chihili, 91340, 94-158, 93-8, 92-13, 93-7, 93-4, 94-157, 93-36, 87-413, 94-190, 94-224, 95-100, Men yaku |
pekinensis |
КИТ |
B |
0 |
|
1231 |
- |
- |
AB |
26.0 |
|
Shiki Sei, 88-22, 87-356, 94-208, 87-411, Aomi, Mei quing choi |
chinensis |
- |
B |
0 |
|
Hon-tsai-tai, Hong-kong, 91-333, 91-23, 92-563 |
parachinensis |
- |
B |
0 |
|
Toria 1, 3159, 3170, 3174 |
dichotoma |
САЗ |
A |
100.0 |
|
3091 |
- |
- |
AB |
71.0 |
|
3096 |
- |
- |
AB |
62.5 |
|
3124 |
- |
- |
BA |
26.3 |
|
3161 |
- |
- |
BA |
31.0 |
|
3163 |
- |
- |
AB |
70.0 |
|
3166/1 |
- |
- |
BA |
27.0 |
|
3166/2 |
- |
- |
BA |
29.5 |
|
3167 |
- |
- |
AB |
58.0 |
|
3171 |
- |
- |
AB |
54.0 |
|
3172 |
- |
- |
AB |
79.0 |
|
3177 |
- |
- |
BA |
28.6 |
|
3181 |
- |
- |
BA |
39.0 |
|
3188 |
- |
- |
BA |
45.0 |
|
3197 |
- |
- |
AB |
87.0 |
|
3198 |
- |
- |
AB |
95.0 |
|
85-247, 3103, BRrc |
- |
- |
B |
0 |
A – «A» тип семян, B – «B» тип, AB и «A»BA – смешанный тип с преобладанием «A» или «B». ЕВР, Европа и Россия, ЯП, Япония, КИТ, Китай, САЗ, Средняя Азия, включая Индию и Непал. ТОС, тип оболочки семян; ЧАТ, частота А типа оболочки семян.
Методы и материалы
Семена 112 образцов B. rapa (табл. 1), собранных в четырех разных регионах: Европе, Средней Азии (включая Пакистан и Непал), Китае и Японии, были предоставлены лабораторией селекции крестоцветных Национального института овощных, декоративных культур и чая (Япония) и Всероссийским НИИ селекции и семеноводства овощных культур. Названия, таксономическая принадлежность и происхождение образцов приведены в таблице 1. Тип оболочки семян определяли путем визуальной оценки состояния эпидермиса семян, погруженных в деионизированную воду на 30 секунд. Как минимум 150 семян было обследовано для каждого образца. Частота встречаемости семян «A» типа приведена в таблицах 1 и 2.
Семена F1 гибридов репы Just Right, Tokyo Cross и сорта Seven Green Top были предоставлены компанией Таки, Япония. Растения репы Just Right, Seven Green Top, White Milan, Leielander, Yorii, дигаплоидной линии пекинской капусты ( B. rapa ssp. pekinensis ) G006, образцов тории ( B. rapa ssp. dichotoma ) 3091, 3177, 85247 и B. rapa ssp. parachinensis 91-333 были самоопылены и скрещены в бутонах между собой, как показано в таблицах 3 и 4.
Результаты и обсуждение
При оценке типа оболочки семян был выявлен полиморфизм внутри образцов, подвидов и географических групп (табл. 1). Европейские образцы, включая турнепс, репу и сурепку (B.rapa ssp. rapa, B.rapa ssp. campestris (L.) A.R. Clapham) имели исключительно семена «B» типа. Растения пак-чой, тай-тсай, хонг-тсай-тай, тсай-шин (B. rapa ssp. chinensis (L.) Halnet, B.rapa ssp. narinosa (L.H. Bailey) Hanelt, B.rapa ssp. parachinensis (L.H. Bailey) Hanelt) и большинство пе-тсай (B. rapa ssp. pekinensis (Lour.) Halnet) сортов также имели семена только «B» типа. Напротив, азиатская репа (B. rapa ssp. rapa), японские листовые овощи мизуна, мибуна (B. rapa ssp. nipposinica (L.H. Bailey) Hanelt) и тория (B. rapa ssp. dichotoma (Roxb.) Hanelt) имели преобладающий «A» тип оболочки семян (табл. 1 и 2). Гибриды F1, полученные от родителей с контрастным типом оболочки семян, всегда имели тип семян аналогичный материнской форме (табл. 3). Среди популяций F2, полученных в результате скрещивания сортов азиатской реп Sugurina и Yorii («A» тип семян) с европейскими репами Milan White и Leielander («B» тип семян), не было обнаружено ни одного семени «B» типа. Аналогично не было обнаружено ни одного семени «A» типа в F2 популяциях, полученных от реципрокных комбинаций («B» х «A»). Оче- видно, что в этих случаях тип оболочки семян полностью определялся материнским фактором (геном). Скрещивание растений с семенами «B» типа между собой, как и ожидалось, не приводило к появлению «A» типа.
91-333, B. rapa ssp. parachinensis. A – «A» тип семян, B – «B» тип, сегрегация по типу семян внутри популяций не была обнаружена; Нд, нет данных
Согласно информации, предоставленной компанией Таки, родительские линии F1 гибрида Just Right имели семена разных типов, но F1 был представлен только семенами «A» типа. Сегрегация в F2 популяции была близкой к 9:7, что предполагает участие двух доминантных генов в эмбрионе с комплементарным взаимодействием, названных далее A1 и A2, и определяющих тип «A» оболочки семян в материнской ткани оболочки семени. Таким образом, в популяции B. rapa тип оболочки семян определялся как материнскими (пока не выявленными), так и отцовскими генами, что предполагает участие регуляторных белков (транскрипционных факторов) эмбриона в формировании фенотипа оболочки семени по примеру растений резухи Таля [7].
Когда семена с оболочкой «A» типа были впервые обнаружены у B. rapa ssp. nipposinica и азиатской репы, возникла гипотеза о новообразованном признаке,
2. Распределение типа оболочки семян в географических группах B. rapa.
|
Регион |
Подвид |
Тип оболочки семян |
ЧО |
ЧАТ, % |
ЧГ, % |
|
Европа |
rapa, campestris |
B |
39 |
0 |
- |
|
Средняя Азия |
dichitoma |
AB |
22 |
86.3 |
63.3 |
|
Япония |
nipposinica (включая rapa) |
AB |
15 |
95.7 |
75.2 |
|
parachinensis |
B |
6 |
0 |
- |
|
|
Китай |
chinensis pekinensis |
B B(A) |
7 19 |
0 5.3 |
-26.0 |
|
narinosa |
B |
4 |
0 |
- |
ЧО - число изученных образцов, ЧАТ - частота образцов с «A» типом семян, %, ЧГ - средняя частота ‘A» типа семян среди образцов группы, %.
3. Наследование типа оболочки семян у внутривидовых гибридов B. rapa.
|
Популяция |
Тип оболочки семян родителей, F1 и F2 |
|||
|
t : |
: u |
F 1 |
F 2 |
|
|
Just Right F1 x Seven Top Green |
A |
B |
A |
A |
|
Seven Top Green x Just Right F1 |
B |
A |
B |
B |
|
Sugulrina x Milan white |
A |
B |
A |
A |
|
Yori x Milan white |
A |
B |
A |
A |
|
Milan white x Yori |
B |
A |
B |
Нд |
|
Yori x Leielander |
A |
B |
A |
A |
|
3091 x G006 |
A |
B |
A |
Нд |
|
G006 x 3091 |
B |
A |
B |
B |
|
3177 x G006 |
A |
B |
A |
Нд |
|
G006 x 3177 |
B |
A |
B |
Нд |
|
G006 x 85-247 |
B |
B |
B |
B |
|
85-247 x G006 |
B |
B |
B |
B |
|
G006 x 91-333 |
B |
B |
B |
Нд |
|
91-333 x G006 |
B |
B |
B |
Нд |
Just Right, Seven Green Top, White Milan, Leielander, Yorii, сорта репы;
G006, дигаплоидная линия пекинской капусты; 3091, 3177, 85-247, образцы тории;
91-333, B. rapa ssp. parachinensis. A - «A» тип семян, B – «B» тип, сегрегация по типу семян внутри популяций не была обнаружена; Нд, нет данных свойственном только японской популяции этого вида [12]. Тип оболочки семян позволил надежно разделить азиатские репы и интродуцированные в Японии европейские турнепсы [14, 15]. Вид B. rapa включает в себя морфологически разнородные и географически разделенные подвиды. Подвиды среднеазиатского происхождения – масличные тория и желтый сарсон B.rapa ssp. trichu-laris считаются близкими к европейской популяции, представленной турнепсом, репой, рапсом и сурепкой. Восточноазиатские листовые овощи (B.rapa ssp. chinensis и др.) считаются группой, наиболее удаленной от гипотетического прародителя [16]. Подвиды B. rapa были определенны в основном на основе различий строения хозяйственно используемых частей растения: корнеплода, листьев и соцветия. В общем, они совпадают с генетическими, или точнее, биогеографическими группами, которые эволюционировали в течении длительного периода изоляции, адаптации к различным условиям среды и отбора для определенного способа использования человеком [16, 17]. Нередко растения со сходной морфологией, но разным географическим и филогенетическим происхождением, зачисляются в один подвид. Например, европейский турнепс и азиатская репа объединяются в подвид
B. rapa ssp. rapa . Наиболее показательным примером такого искусственного слияния является группа B. rapa ssp. campestris, представленная зачастую одичавшими растениями типичных для конкретного региона культурных форм вида. Часто растения, близкие друг другу по документированному происхождению и генетическим маркерам, разносятся в разные подвиды на основании различия по тем же самым хозяйственно ценным признакам, контролируемым одним-двумя генами. Примером такого разделения является рапс ( B. rapa ssp. oleifera ), отобранный из западноевропейского турнепса [17].
Наивысшая частота семян «A» типа была обнаружена среди растений среднеазиатской популяции. По мнению Синской [18] и других исследователей [17] эта популяция представляет собой наиболее древнюю из сохранившихся до наших дней группу культурных растений B. rapa. Азиатские репы и отобранные из них японские листовые овощи кабуна, мизуна и мибуна (B. rapa ssp. nip-posinica) с семенами «A» типа (К. Хида, неопубликовано) признаются родственными примитивным афганским и индийским репам [13, 14, 18]. Последние составляют одну генетическую популяцию с торией [17]. Семена «A» типа были также обнаружены у одного из образцов пе-тсай (пекинской капусты). Предыдущими исследователями было обнаружено незначительное число «A» семян в популяции хонг-тсай-тай (B. rapa ssp. parachinensis) [12, 19]. Сведения о происхождении пе-тсай и хонг-тса-тай противоречивы. Так как они имеют ряд общих биохимических и молекулярных маркеров с азиатской репой и торией [17, 20], возможно, что пе-тсай возник в результате прямого отбора из азиатской репы или после её гибридизации с пак-чой (B. rapa ssp. chinensis), а хонг-тсай-тай – как гибрид между пак-чой и масличными растениями, родственными тории.
Присутствие фенотипически нейтрального признака, контролируемого комплексом генов, в географически удаленных популяциях дает подтверждение идее о филогенетической связи азиатских (японских) реп с центральноазиатской популяцией B. rapa. Напротив, южно-китайская популяция B. rapa не является прямым прародителем японских листовых овощей B. rapa ssp. nipposinica . Вторичное сходство некоторых генетических и морфологических признаков этих популяций могло бы быть объяснено конвергенцией в сходной эколого-географической зоне. На основании сходства генетических признаков и общего происхождения, было
4. Наследование типа оболочки семян в потомстве репы Just Right.
|
Популяция |
Число семян типа: |
Ожидаемое соотношение |
P> |
||
|
A |
B |
Всего |
A:B |
||
|
Родительская линия 1 |
100 |
0 |
100 |
- |
- |
|
Родительская линия 2 |
0 |
100 |
100 |
- |
- |
|
F 1 Just Right |
150 |
0 |
150 |
- |
- |
|
Just Right F 2 |
282 198 |
480 |
270:210 (9:7) |
0.75 |
|
Данные по родительским линиям предоставлены компанией Таки.
A – «A» тип семян, B – «B» тип.
бы разумным перенести азиатскую группу реп из подвида B.rapa ssp. rapa в B. rapa ssp. nipposinica .
Поскольку ни один из изученных 68 образцов B. napus, представляющих Евразийскую популяцию от Великобритании до Японии, не имел семян «A» типа (данные не приведены), гипотетический диплоидный прародитель c геномом АА для амфидиплоида B. napus [16] вероят- нее всего не происходит из центральноазиатской популяции B. rapa. Вероятно, что синтез этого вида произошел в результате гибридизации B. oleracea с уже одомашненной популяцией B. rapa с семенами «B» типа.
Мы планируем провести анализ наличия и роли генов, контролирующих наследование гомологичного признака у растений резухи Таля [7, 8] в популяции
B. rapa , расщепляющейся по типу семян. Поскольку тип оболочки семени тесно связан у растений с продолжительностью жизни семян (скоростью биологического старения), знергией прорастания, адаптивностью к прорастанию в условиях дефицита влаги в почве [21], то он является желательным признаком у овощных и масличных культур вида B. rapa .
Список литературы Распределение и наследование «A» типа оболочки семян в популяции brassica rapa l
- Windsor J.B, Symonds V.V, Mendenhall J, Lloyd A.M. 2000.Arabidopsis seed coat development: morphological differentiation of the outer integument. Plant Journal22: 483-493.
- Western T.L. 2012.The sticky tale of seed coat mucilages: production, genetics, and role in seed germination and dispersal.Seed Science Research22: 1-25.
- Yang X, Baskin J, Baskin C, Huang Z. 2012b. More than just a coating: ecological importance, taxonomic occurrence and phylogenetic relationships of seed coat mucilage.Perspectives in Plant Ecology, Evolution and Systematics14: 434-442.
- Western T.L, Skinner D.J, Haughn G.W. 2000. Differentiation of mucilage secretory cells of the Arabidopsis seed coat. Plant Physiology122: 345-356.
- Western T.L, Burn J, Tan W.L, et al. 2001. Isolation and characterization of mutants defective in seed coat mucilage secretory cell development in Arabidopsis.Plant Physiology127: 998-1011.
- North H.M, Berger A, Saez-Aguayo S, Ralet M.C. Understanding polysaccharide production and properties using seed coat mutants: future perspectives for the exploitation of natural variants. Ann Bot. 2014 Oct;114(6):1251-63 DOI: 10.1093/aob/mcu011
- Western T.L, Young D.S, Dean G.H, Tan W.L, Samuels A.L, Haughn G.W. 2004. MUCILAGE-MODIFIED4 encodes a putative pectin biosynthetic enzyme developmentally regulated by APETALA2, TRANSPARENT TESTA GLABRA1, and GLABRA2 in the Arabidopsis seed coat. Plant Physiology 134: 296-306.
- Zou H.F, Zhang Y.Q, Wei W, Chen H.W, Song Q.X, Liu Y.F, Zhao M.Y, Wang F, Zhang B.C, Lin Q, Zhang W.K, Ma B, Zhou Y.H, Zhang J.S, Chen S.Y. The transcription factor At DOF4.2 regulates shoot branching and seed coat formation in Arabidopsis. Biochem J. 2013 Jan 15;449(2):373-88 DOI: 10.1042/BJ20110060
- Esfandiari E, Jin Z, Abdeen A, Griffiths J.S, Western T.L, Haughn G.W. Identification and analysis of an outer-seed-coat-specific promoter from Arabidopsis thaliana. Plant Mol Biol. 2013 Jan;81(1-2):93-104 DOI: 10.1007/s11103-012-9984-0
- E Kondo M. Uber die in der Landwirtschaft Japans gerbauchten Samen. I. Ber. Obara Inst. Fur Lanw. Forsch. 1918, 1:2-32.
- Aoba T. Histological observation of seed coat in Brassica juncea Coss.//Japan. J. Breed. 1972, 22: 323-328 (на японском языке).
- Aoba T. Morphological observation of seed coat of the species belonging to Brassiceae. Bull. Yamagata Univ., Agr. Sci. 1971, 6: 315-324.(на японском языке).
- Aoba T. Characteristics of Asian turnips and their phylogenetical relationships. Agriculture & Horticulture 1987, 62: 63-64 (на японском языке).
- Shibutani S., Okamura T. On the classification of turnips in Japan with regard to the types of epidermal layers of the seed. J. J. Horticultural Soc. 1954, 22: 235-238 (на японском языке).
- Shibutani S., Okamura T. Studies on the morphological characters of seeds in Brassica. J. Hort. Ass. Japan. 1955, 24: 3-10 (на японском языке).
- Song K.M., Osborn T.C., Williams P.W. Brassica taxonomy based on nuclear restriction fragment polymorphism (RFLPs). 3. Genome relationships in Brassica and related genera and the origin of B. oleracea and B. rapa (syn. campestris). Theor. Appl. Genet. 1990, 79: 497-506.
- McGrath J.M., Quiros C.F. Genetic diversity at isozyme and RFLP loci in Brassica campestris as related to crop type and geographical origin. Theor. Appl. Genet. 1992, 83: 783-790.
- Sinskaia E.N. The oleiferous plants and root crops of the family Cruciferae. Bull. Appl. Bot. Plant Breed. 1928, 19: 1648.
- Yazawa S., Ueyama H, Namiki T. Simple method to distinguish seed coat type of Brassica campestris. Agriculture & Horticulture 1986, 61: 556-558. (на японском языке).
- Li C.W. The origin, evolution, taxonomy and hybridization of Chinese cabbage. In. P. Takelar, N.S., Griggs, T.D. (eds.) Chinese cabbage. Proc. 1st Int Symp Asian Veg. Res. Dev. Center. Taiwan, 1981: 3-9.
- Yazawa S., Ueyama H., Namiki T. Growing habit and longevity of seeds of Brassica campestris classified by their seed coat type. Agriculture & Horticulture 1988, 63: 1417-1419. (на японском языке).
