Распределение коллембол (Collembola) в градиенте влажности среднетаежных сосновых лесов (на примере заказника «Белый»)
 в градиенте влажности среднетаежных сосновых лесов (на примере заказника «Белый»)")
Автор: Таскаева А.А., Дгтева С.В., Лаптева Е.М., Лапшина Е.Н., Конакова Т.Н.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 2 (14), 2013 года.
Бесплатный доступ
Исследованы состав, структура и численность ногохвосток органогенных го- ризонтов альфегумусовых почв, формирующихся в градиенте влажности среднетаежных сосновых лесов. Показана связь видового разнообразия и численности сообщества коллембол с характером почвенно-растительного по- крова и погодными условиями года (температура воздуха, сумма осадков). Выявлены изменения численности доминирующих видов, соотношения раз- личных групп гигропреферендума и жизненных форм ногохвосток в градиен- те влажности сосновых лесов.
Коллемболы, сосновые леса, подзолы, градиент влажности, особо охраняемые природные территории
Короткий адрес: https://sciup.org/14992602
IDR: 14992602 | УДК: 595.713:630*187:582.475:502.4(470.13)
Collembola distribution at gradient of soil moisture of the middle taiga pine forests (on the example of reserve «Belyi»)
Composition, structure and density of collembolan of organic horizons of alphehumus soils, formed at gradient of soil moisture of the middle taiga pine forests, are researched. There is shown connection between collembolan species diversity and community density and the character of soil-vegetative cover and weather conditions (air temperature, precipitations). Changes of number of dominants, correlation between different groups of hygropreferences and vital forms of collembolan at gradient of soil moisture of pine forests are revealed.
Текст научной статьи Распределение коллембол (Collembola) в градиенте влажности среднетаежных сосновых лесов (на примере заказника «Белый»)
В таежных экосистемах ведущую роль в разложении растительного опада и формировании органогенных горизонтов таежных почв играют такие представители группы микроартропод, как ногохвостки и панцирные клещи [1]. Они не только участвуют в измельчении и минерализации растительных остатков, но и регулируют состав и сукцессии микроорганизмов, скорость круговорота зольных элементов, могут служить индикаторами процессов почвообразования и антропогенного воздействия на природные экосистемы.
В настоящее время детально изучено население почвообитающих коллембол еловых лесов [2,3,7,8]. Сосновые леса, особенно среднетаежные, в этом плане исследованы в существенно меньшей мере. Основной массив данных о структуре и составе сообществ коллембол получен на примере как естественных, так и антропогенно нарушенных сосновых лесов южной тайги [4,5]. Имеются работы, характеризующие комплексы ногохвосток северотаежных сосняков Кольского полуострова, Архангельской обл. и сосновых лесов Печоро-Илычского заповедника, расположенных в подзоне средней тайги [2]. Кроме того, частично исследовано насе- ление почвообитающих коллембол в сосновых лесах, находящихся в зоне влияния Сыктывкарского лесопромышленного комплекса [6]. В данных работах, направленных на выявление закономерностей формирования фауны и населения коллембол в лесных почвах, рассматривались тенденции изменения сообществ ногохвосток в биотопах, образующих ординационный ряд либо по увеличению влажности и заболоченности [4,5,7], либо по уровню почвенного плодородия [8]. В результате проведенных исследований показано, что основными факторами, определяющими структуру и динамику комплексов ногохвосток, являются влажность и трофность почвы [2]. При этом влажность биотопа для коллембол в целом более значима, чем троф-ность почв [8].
В Республике Коми сосновые леса из сосны обыкновенной – Pinus sylvestris – составляют около 16.8% территории. Сообщества данной лесной формации занимают второе место в структуре лесопокрытой площади после ельников. При этом они распространены неравномерно. Основные массивы сосняков приурочены к подзоне средней тайги. В подзонах крайнесеверной и северной тайги, а также в Приуралье и на Урале сосна обыкновенная встречается редко [9]. С учетом изложенного выявление закономерностей формирования комплекса ногохвосток в светлохвойных лесах республики представляет несомненный интерес. При изучении пространственно-временного распределения и естественных сукцессий почвенной биоты наиболее удобны экосистемы особо охраняемых природных территорий (ООПТ), где антропогеннное воздействие либо отсутствует, либо минимально. Нами была поставлена цель – выявить закономерности изменения фауны и населения коллембол сосновых лесов в градиенте влажности на территории ландшафтного заказника «Белый». Данный резерват расположен в подзоне средней тайги Республики Коми на территории МО МР «Усть-Вымский» и создан в 1989 г. для регулирования антропогенных нагрузок в сосновых лесах, традиционно используемых местным населением для рекреации [9].
Материал и методы
Исследования проведены в 2007–2010 гг. В качестве ключевых участков на территории заказника «Белый» были выбраны сообщества трех ассоциаций: сосняк вересково-лишайниковый (С1), сосняк бруснично-лишайниково-зеленомошный (С2) и сосняк пушицево-багульниково-сфагновый (С3). Они расположены на боровой террасе р. Вычегда и образуют естественный ряд по степени нарастания увлажнения почв. Участки С1 и С2 приурочены к хорошо дренированным плоским элементам рельефа. Участок С3 расположен в заболоченном понижении и граничит с облесённым пушицевосфагновым болотом. В пределах каждого участка изучены состав и структура фитоценозов, исследованы специфика морфологического строения и физико-химических свойств соответствующих им почв, состав и численность сообществ почвообитающих ногохвосток. Исследования проводили с применением стандартных методов геоботаники и фитоценологии [10–12], почвоведения [13,14] и почвенной зоологии [15].
Отбор проб почв для изучения населения коллембол осуществляли по методу, предложенному Н.М. Черновой [15]. Пробы площадью 25 см2 каждая отбирали в 10-кратной повторности с учетом мощности органогенного горизонта (от 1–2 см в сосняке вересково-лишайниковом до 15 см – в сосняке багульниково-сфагновом) в 2007 г. (август), 2008 г. (август) и 2010 г. (июнь, август, сентябрь). Всего за весь период было отобрано 170 почвенных образцов, определено около 7 тыс. экземпляров коллембол. Для более полного выявления таксономического разнообразия ногохвосток в период с 26 мая по 30 июня 2010 г. на каждом участке были установлены ловушки Барбера, с помощью которых учтено более 3.5 тыс. экземпляров ногохвосток. За основу выделения классов доминирования принята шкала Энгельмана [2]. Группы гигропреферендумов определены согласно данным Н.А. Кузнецовой [16].
Краткая характеристика погодных условий по годам наблюдений представлена в табл. 1. Как видно из приведенных данных, вторая половина летне-
Таблица 1
Средние показатели температуры воздуха и осадков по годам наблюдений (по данным метеостанции «Усть-Вымь»)
|
Год наблюдения |
Июнь |
Июль |
Август |
Сентябрь |
||||
|
I |
II |
I |
II |
I |
II |
I |
II |
|
|
2007 |
11.2 |
49 |
18.9 |
63 |
16.0 |
93 |
8.8 |
39 |
|
2008 |
14.3 |
48 |
18.6 |
42 |
13.3 |
144 |
6.5 |
72 |
|
2010 |
13.2 |
111 |
19.8 |
12 |
15.1 |
42 |
7.7 |
39 |
Примечание. I – среднемесячная температура, °С; II – сумма осадков за месяц, мм.
го периода 2008 г. отличалась избыточной влажностью и недостатком тепла, а 2010 г. – недостатком влаги при более теплом и избыточно влажном начале лета (июнь) по сравнению с двумя остальными годами наблюдений.
Результаты исследований
Как показали проведенные исследования, наиболее сухое местообитание в рассмотренном ряду биотопов занимает сосняк вересково-лишайниковый (ключевой участок С1) . Древостой из сосны обыкновенной данного насаждения чисто сосновый. Он состоит из двух пологов. Общая сомкнутость крон – от 0.3 до 0.7 м, максимальная высота древесного яруса – 14–18 м. В подросте зарегистрированы три вида: сосна обыкновенная – Pinus sylvestris , осина – Populus tremula , ель сибирская – Picea obovata . Наиболее активно возобновляется сосна. Подлеска нет. Сомкнутость травяно-кустарничкового яруса невелика. Значение показателя общего проективного покрытия (ОПП) не превышает 25%, растения располагаются небольшими группами. Их разнообразие невелико, всего в составе яруса отмечено 15 видов сосудистых растений. Более 80 % от общего проективного покрытия приходится на вереск обыкновенный ( Calluna vulgaris ), до 5–10% занимают брусника ( Vaccinium vitis-idaea ), осока верещатниковая ( Carex ericetorum ) и толокнянка обыкновенная ( Arctostaphylos uva-ursi ). Единично встречаются ястребинка волосистая ( Hiera-cium pilosella ), черника ( Vaccinium myrtillus ) и водяника гермафродитная ( Empetrum hermaphroditum ). Напочвенный покров сплошной (ОПП 90–98 %), мозаичного сложения. Преобладают кустистые лишайники: кладония лесная ( Cladonia arbuscula ) , к. оленья ( C. rangiferina ) , к. звездчатая ( C. stellaris ), по ковру которых разбросаны пятна зеленых мхов, образованные плеурозием Шребера ( Pleurozium schreberi ), дикраном многоножковым ( Dicranum polysetum ). Состав и структура нижних ярусов фитоценоза отражают особенности экотопа, характеризующегося наиболее сухими и бедными почвами. Их облик определяют лишайники, отличающиеся высокой толерантностью по отношению к дефициту влаги и элементов минерального питания, и ксеро-мезофитные сосудистые растения.
Ключевой участок С2, где представлен фитоценоз ассоциации сосняк бруснично-лишайниковозеленомошный, занимает более влажное место- обитание. Состав и структура древостоя такие же, как и на ключевом участке С1. Травяно-кустарничковый покров бедный по видовому составу, разреженный (ОПП до 10 %). Его облик определяет уже не вереск обыкновенный (Calluna vulgaris), а брусника (Vaccinium vitis-idaea). Среди константных малообильных видов следует упомянуть вейник наземный (Calamagrostis epigeios), иван-чай узколистный (Chamaenerion angustifolium), золотарник обыкновенный (Solidago virgaurea). Напочвенный покров сплошной (ОПП 90-98 %). В нем более значимую ценотическую роль играют не кустистые лишайники, а мохообразные, из которых обилен плеурозий Шребера (Pleurozium schreberi).
Завершает рассматриваемый ряд местообитаний на градиенте влажности ключевой участок С3, где сформировался сосняк пушицево-багульниково-сфагновый. Древостой более сложный по составу и структуре. Он образован сосной обыкновенной с редкой примесью берёзы пушистой, состоит из трёх пологов, сомкнутость которых составляет от 0.2 до 0.4. В подлеске обычна берёза карликовая. Средняя видовая насыщенность травянокустарничкового яруса, как и в сообществах сосняков других типов, невелика – 10-11 таксонов, но значения общего проективного покрытия растений значительно выше – 40-70 %. Наиболее постоянные и обильные виды – пушица влагалищная ( Eryophorum vaginatum ) и багульник болотный ( Ledum palustre ) . Заметно участие морошки ( Rubus chamaemorus ), менее обильны при высокой константности кассандра ( Chamaedaphne calyculata ), голубика ( Vaccinium uliginosum ). В сплошном напочвенном покрове ключевые позиции занимают влаголюбивые сфагнум узколистный ( Sphagnum angustifolium ), с. береговой ( S. riparium ), в отдельных случаях до 33 % составляет удельное обилие с. центрального ( S. centrale ). Роль зеленых мхов снижена, они встречаются в основном на кочках. Наиболее обычны плеурозий Шребера ( Pleurozium schreberi ), кукушкин лен обыкновенный ( Polytrichum commune ).
По мере нарастания влажности в рассмотренном ряду биотопов происходит закономерное изменение морфологического строения почв, формирующихся под пологом сосновых лесов. Почвенный покров участков С1 и С2 представлен подзолами иллювиально-железистыми1, участка С3 – торфяно-подзолами глеевыми. Для первых строение профиля может быть выражено формулой О-Е-BF-C, для вторых – Т-Eg-BFHg-BG-CG (рис.1).
Почвы ключевых участков С1 и С2 имеют типичное для подзолов иллювиально-железистых строение. Их профиль характеризуется сочетанием слабооторфованного рыхлого подстилочноторфяного (О), белесого подзолистого (Е) и альфе-гумусового (BF) горизонтов, постепенно переходящих в материнскую породу (С). Подстилочно-торф-яный горизонт, как правило, дифференцирован на
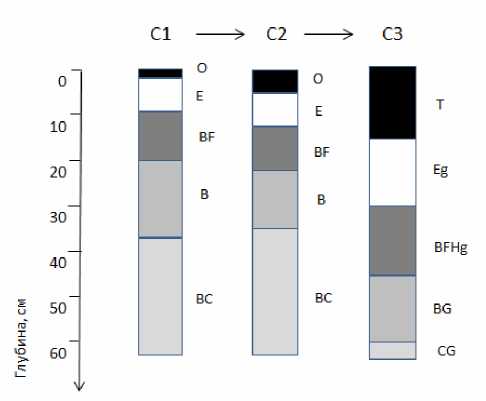
Рис.1. Схема морфологического строения профиля почв на ключевых участках сосняка вересковолишайникового (С1), бруснично-лишайниково-зеленомошного (С2), пушицево-багульниково-сфагнового (С3).
две части – слаборазложенную (верхнюю) и более хорошо разложенную (нижнюю). Присутствие на границе с подзолистым горизонтом углистых частиц свидетельствует о послепожарном происхождении современного растительного покрова заказника «Белый». На участке С1 мощность подстилочноторфяного горизонта составляет 1–2 см, подзолистого – не превышает 10 см, что позволяет отнести почву данного участка к иллювиально-железистым поверхностно-осветленным подзолам. На участке С2, где напочвенный покров представлен преимущественно зелеными мхами, на поверхности почвы накапливается органический материал в виде более мощного (3–5 см) оторфованного органогенного горизонта О. В зависимости от мощности подзолистого горизонта здесь представлены три вида подзолов иллювиально-железистых: поверхностно-осветленные (мощность горизонта Е < 10 см), мелко-осветленные (10–20 см) и неглубоко-осветленные (20–30 см). Мощность подзолистого горизонта возрастает в направлении от межкроновых пространств к прикомлевым участкам. В рассмотренных профилях отсутствуют морфохроматические признаки оглеения, однако наличие в минеральной толще мелких Fe-Mn-конкреций и примазок свидетельствует о возможном временном переувлажнении почв участка С2. Специфической особенностью морфологического строения торфяно-подзолов глеевых, развитых под пологом сосняков сфагновых (участок С3), является формирование мощного (от 10 до 40 см) торфяного горизонта. Под ним залегает оглеенный подзолистый горизонт Eg, последовательно переходящий в темноокрашенный иллювиально-гумусово-железистый BHFg горизонт с признаками оглеения, оглеенный горизонт BG и почвообразующую породу CG.
Подзолы иллювиально-железистые кислые, ненасыщены основаниями, малогумусны, бедны элементами питания [17]. Песчаный состав почво- образующих пород и глубокое залегание почвенногрунтовых вод обусловливают хорошую прогреваемость верхних горизонтов в летние месяцы (рис.2), дефицит влаги в период вегетации растений (табл.2), и, соответственно, невысокую биологическую продуктивность растительного покрова, приуроченного к данному типу почв. По уровню кислотности и ненасыщенности основаниями торфяно-подзолы глеевые близки к подзолам иллювиальножелезистым. При этом они характеризуются повышенной влажностью даже в наиболее «сухие» по гидрологическому режиму годы (табл.2) и благодаря застою влаги на 2-4° холоднее подзолов, что наиболее ярко прослеживается в летний период (рис.2).
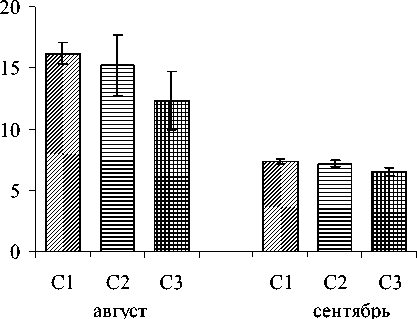
Рис. 2. Показатели температуры (по данным 2010 г.) органогенных горизонтов почв, формирующихся на ключевых участках соснового леса. Условные обозначения те же, что на рис.1.
Таблица 2
Показатели влажности (% массы почвы) органогенных горизонтов почв ключевых участков по годам наблюдений и датам отбора проб
|
Ключевой участок |
2008 г. |
2010 г. |
||
|
VIII |
VI |
VIII |
IX |
|
|
С1 |
71,9±19,9 |
59,2±27,4 |
1,6±0,6 |
99,4±16,9 |
|
49,0-104,1 |
35,5-89,2 |
0,9-2,2 |
81,4-114,9 |
|
|
С2 |
168,1±62,1 |
111,2±69,8 |
2,4±2,6 |
243,9±165, 1 |
|
48,3-326,1 |
138,1-224,7 |
1,2-7,6 |
311,2-436,5 |
|
|
С3 |
800,2±143,6 |
770,9±205,4 |
171,1±142,3 |
556,4±3,5 |
|
685,9-1040,3 |
597,3-997,7 |
10,2-280,2 |
552,4-559,2 |
|
Примечание. В числителе х ± s , где Х - среднее арифметическое, S – стандартное отклонение среднего арифметического; в знаменателе – минимум-максимум.
температуры и влажности соответствует подзолам сосняка вересково-лишайникового, под зеленомошными – занимает промежуточное положение между почвами сосняка вересково-лишайникового и сосняка пушицево-багульниково-сфагнового.
На территории заказника «Белый» в совокупности зарегистрировано 49 видов коллембол, заселяющих органогенные горизонты подзолов иллювиально-железистых и торфяно-подзола глеевого. Впервые для Республики Коми здесь выделены такие виды, как Entomobrya multifasciata (Tullberg, 1871) , Entomobrya superba (Reuter, 1876) , Sminthu-rus viridis (Linnaeus, 1758) . Наибольшим таксономическим разнообразием отличались ключевые участки С1 и С3 (рис.3). Полученные нами данные свидетельствуют о существенно меньшем видовом разнообразии населения коллембол в почвах среднетаежных сосновых лесов по сравнению с еловыми, где выявлено 59 видов [3]. При этом уровень таксономического разнообразия близок к величине показателя (46 видов), отмеченной для сосновых лесов подзоны южной тайги [2]. В исследованных нами фитоценозах южнотаежных сосновых лесов как и в ельниках средней тайги, наиболее богаты в видовом отношении семейства Isotomidae (12 видов), Neanuridae (8), Onychiuridae (6) и Hypogastru-ridae (5).
Число видов, учтенных при помощи ловушек Барбера, варьирует от 8 (сосняк бруснично-лишайниково-зеленомошный) до 13 (сосняк пушицево-багульниково-сфагновый). Наибольшая уловистость ногохвосток отмечена в сосняке лишайниковозеленомошном (68 экз./10 лов.-сут). Во всех сосняках основу населения коллембол, собранных при помощи ловушек, составляют два вида: Orchesella cincta (Linnaeus, 1758) и Orchesella flavescens (Bour-let, 1839). На их долю в разных сообществах приходится от 71 до 98 %. Если уловистость последнего вида в конце мая варьирует от 8 до 30 экз./10 лов.-сут, то в конце июня она значительно ниже – 1–7 экз./10 лов.-сут.
В сосняке вересково-лишайниковом как наиболее сухом биотопе в совокупности выявлен 31 вид коллембол (рис.3). Их общая численность здесь варьировала в зависимости от срока наблюдения от 0.4 до 82 тыс. экз./м2 (табл. 3), а улови-стость составила 37-44.7 экз./10 лов.-сут. По численности преобладали ксерорезистентные виды – 57.7%, в то время как доли мезофильных форм и видов без четкого гигропреферендума составляли 14.1 и 26.5% соответственно (рис. 4). В подстилочно-торфяном горизонте почвы данного биотопа отмечено четыре потенциальных доминанта, т.е. вида, превышающего порог доминирования хотя бы в один год исследования. Постоянным эудоминантом выступал кортицикольный вид Anurophorus palae-arcticus , доля которого во все сроки наблюдений была стабильно выше 40%. В зависимости от погодных условий временное преимущество получали разные виды. Например, почвенные виды Protapho-rura boedvarssoni и Isotomiella minor достигали высокого уровня обилия в 2007 и 2008 гг. соответственно. В 2010 г., характеризующемся минимумом
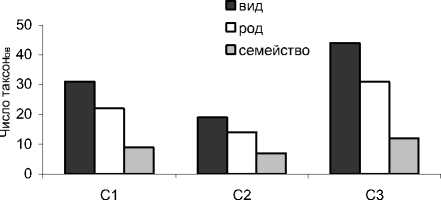
Рис.3. Таксономическое разнообразие коллембол в разных типах сосняков. Условные обозначения те же, что на рис.1.
С1
С2
С3
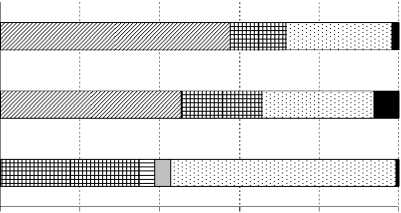
0% 20% 40% 60% 80% 100% и Ксерорезистентные ш Мезофильные
□ Гигрофильные
□ Ксеро-мезофильные в Мезо-гигрофильные
□ Без четкого гигропреферендума
Таблица 3
Характеристика сообществ коллембол в различных биотопах заказника «Белый» по годам наблюдений
|
Показатель |
2007 г. |
2008 г. |
2010 г. |
|
август |
август |
июнь август сентябрь |
Сосняк вересково-лишайниковый (участок С1)
Количество видов
|
в серии |
12 |
15 |
11 |
1 |
10 |
|
Среднее |
|||||
|
число видов |
|||||
|
в пробе |
6.2 |
13.0 |
4.7 |
0.7 |
3.7 |
|
Общая численность |
63.5± |
82.1± |
10.2± |
0.4± |
12.8± |
|
(тыс. экз./м2) ± m |
18.6 |
23.4 |
1.9 |
0.1 |
2.0 |
|
Агрегированность* |
10.9 |
9.1 |
3.0 |
0.6 |
2.8 |
Сосняк бруснично-лишайниково-зеленомошный
|
(участок С2) |
|||||
|
Количество видов в серии |
- |
11 |
8 |
2 |
4 |
|
Среднее число видов в пробе |
- |
6.5 |
4.0 |
0.4 |
1.4 |
|
Общая численность |
47.6± |
9.4± |
1.8± |
3.8± |
|
|
(тыс. экз./м2) ± SD |
- |
10.6 |
1.7 |
1.0 |
1.4 |
|
Агрегированность* |
- |
5.4 |
2.8 |
2.3 |
3.6 |
|
Сосняк пушицево-багульниково-сфагновый |
|||||
|
(участок С3) |
|||||
|
Количество видов в серии |
20 |
9 |
14 |
6 |
13 |
|
Среднее число видов в пробе |
8.5 |
7.2 |
3.7 |
1.2 |
3.4 |
|
Общая численность |
61.3± |
6.6± |
6.7± |
2.6± |
9.2± |
|
(тыс. экз./м2) ± SD |
12.6 |
3.1 |
2.5 |
1.0 |
2.0 |
|
Агрегированность * |
7.6 |
1.5 |
4.7 |
2.9 |
3.3 |
Примечание. * – индекс Лексиса; знак « - » означает, что пробы не отбирали в данный срок наблюдения; m – ошибка средней.
осадков, наиболее многочисленным был Desoria tolya . Соотношение численности указанных видов оказалось весьма неустойчивым: некоторые из них в отдельные годы исследований не были обнаружены (табл. 4). Перестройки набора доминантов сопровождались изменениями в спектре жизненных форм. Так, в 2007 и 2008 гг. в населении ногохвосток преобладали представители кортицикольной и почвенной групп, в 2010 г. – кортицикольные и верхнеподстилочные формы (рис. 5). Пространственное распределение коллембол во влажные годы (2007–2008 гг.) было более агрегированным (значения индекса Лексиса 9.0-10.9) по сравнению с сухим 2010 г. (0.6–3.0).
■ Не определен
Рис. 4. Соотношение групп гигропреферендумов кол-лембол по численности в разных типах сосняков. Условные обозначения те же, что на рис. 1.
2007 г.
2008 г.
2010 г.
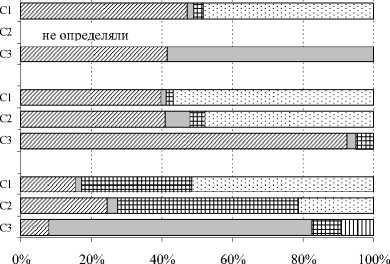
и Почвенная
□ Кортицикольная
□ Гемиэдафическая я Верхнеподстилочная
□ Нейстонная
Рис. 5. Изменение спектров жизненных форм кол-лембол в разных типах сосняков по годам наблюдений. Условные обозначения те же, что на рис. 1.
В сосняке бруснично-лишайниково-зеленомошном (С2) зарегистрировано наименьшее число видов ногохвосток – 19 (рис.3). Общая численность коллембол невысока, по срокам наблюдения варьировала от 1.8 до 47.6 тыс. экз./м2 (табл.3), что в целом соответствует данным литературы. В среднетаёжных сосняках лишайниково-зеленомошных Средней Сибири отмечено около 1.4 тыс. экз./м2 [18], Московской обл. – 14–35 тыс. экз./м2 [4]. На участке С2, как и на участке С1, ведущие позиции по численности занимали ксерорезистентные виды – 45.5%, мезофилы – 20.1 и виды без четкого гигро-преферендума – 28.1% (рис. 4). Среди доминантов в целом преобладали ксерорезистентные виды, но иногда возрастала роль типичных мезофилов – Mi-craphorura absoloni, Willemia anophthalma, Isoto-miella minor. Кроме того, в некоторые сроки отбора в число доминантов входили Anurophorus palaearc-ticus , Desoria tolya, Orchesella flavescens , Xenylla mucronata (табл.4). Постоянным субдоминантом был Protaphorura boedvarssoni . Среди жизненных форм также, как и в сосняке вересково-лишайниковом, преобладали две группы: почвенная и корти-цикольная, однако в 2010 г. отмечено возрастание роли верхнеподстилочных обитателей (рис. 5).
Таблица 4
Численность (тыс. экз./м2) доминирующих видов коллембол в сосняках заказника «Белый» по годам наблюдений (I – 2007 г.; II – 2008 г.; III – 2010 г.)
|
Вид |
С1 |
С2 |
С3 |
||||||
|
I |
1 II |
III |
I |
II 1 |
III |
I |
II |
III |
|
|
Protaphorura boedvarssoni |
27.4 |
2.2 |
0.4 |
н/о |
2.2 |
0.4 |
3.4 |
4.6 |
0.3 |
|
Anurophorus palaearcticus |
30.6 |
46.1 |
3.8 |
-- |
12.0 |
– |
– |
– |
< 0.1 |
|
Desoria tolya |
– |
1.1 |
1.2 |
-- |
1.0 |
1.6 |
– |
< 0.1 |
< 0.1 |
|
Isotomiella minor |
1.2 |
19.3 |
0.6 |
-- |
1.1 |
– |
15.9 |
0.8 |
< 0.1 |
|
Micraphorura absoloni |
– |
1.7 |
– |
-- |
3.8 |
– |
– |
– |
< 0.1 |
|
Xenylla mucronata |
– |
– |
0.1 |
-- |
0.6 |
1.0 |
– |
– |
– |
|
Mesaphorura italica |
– |
– |
0.2 |
-- |
– |
0.8 |
– |
– |
< 0.1 |
|
Orchesella flavescens |
0.6 |
– |
< 0.1 |
-- |
– |
0.6 |
– |
– |
< 0.1 |
|
Folsomia quadrioculata |
0.5 |
– |
< 0.1 |
-- |
– |
– |
29.5 |
– |
1.4 |
|
Parisotoma notabilis |
– |
– |
– |
-- |
– |
– |
– |
– |
1.8 |
|
остальные виды |
4.4 |
14.2 |
1.8 |
-- |
6.7 |
0.4 |
12.5 |
1.1 |
2.5 |
Примечание. «–» – вид не обнаружен; н/о – пробы не отбирали.
Пространственное распределение ногохвосток в почве данного биотопа по годам наблюдений оказалось более стабильным (значения индекса Лек-сиса 2.3-5.4) по сравнению с другими исследованными биотопами, для которых величины этого показателя варьируют в широких пределах (для С1 разброс значений составил 0.6–10.9, для С3 – 1.5– 7.6) (табл. 3).
В заболоченном сосняке (С3) отмечено самое высокое разнообразие коллембол – 44 вида. Возможно, это обусловлено переходным положением данного участка на градиенте от автоморфных (сосняки лишайниковые и зеленомошные) к гидроморфным (верховое болото) позициям. В связи с этим в зависимости от погодных условий года здесь можно наблюдать совмещение (или смену) экологических условий, характерных как для лесных, так и болотных местообитаний. Общая численность коллембол в данном биотопе варьировала от 2.6 до 61.3 тыс. экз./м2 (табл.3), а уловистость составила 30–35 экз./10 лов.-сут. В сообществе ногохвосток были зарегистрированы представители практически всех типов гигропреферендума – от ксерорези-стентного до гигрофильного, однако ведущие позиции занимали в основном мезофильные таксоны (34.8%) и виды без четкого гигропреферендума (56.5%). Среди доминантов следует отметить такие виды, как Fol-somia quadrioculata , Isotomiella minor , Protaphorura boedvarssoni . Их вклад в таксоцен различался в зависимости от года наблюдений (табл. 4). Спектр жизненных форм также существенно варьировал по годам (рис. 5). В прохладные, избыточно влажные годы (2008 г.) на первое место выходили представители почвенной жизненной формы, в сухие, жаркие – геми-эдафические виды. Пространственное распределение ногохвосток было наиболее агрегировано во влажный 2007 г. (значение индекса Лексиса 7.6). В остальные годы агрегированность была либо ниже, чем на участках С1 и С2 (2008 г.), либо примерно на таком же уровне (2010 г.) (табл. 3).
Обсуждение результатов
Согласно данным литературы [4,5,7,19], на градиенте влажности почвы от экотопов с максимальными значениями экологического фактора (сосняки сфагновые) в направлении к местообитаниям со средними величинами параметра (сосняки зеленомошные), как правило, возрастают общее обилие коллембол и численность большинства видов, увеличивается доля представителей почвенной жизненной формы. Доля видов поверхностно-обитаю-щей группы увеличивается от центральной части градиента к его концам: в направлении к сухим биотопам – в основном кортицикольной, к сырым – нейстонной и верхнеподстилочной жизненных форм [2]. Результаты, полученные нами для сосновых лесов подзоны средней тайги, характеризующейся умеренно-континентальным климатом, отражают иную закономерность. Разнообразие и численность коллембол, населяющих органогенные горизонты почв, снижаются от экотопов, занимающих крайние позиции на градиенте влажности (участки С1, С3) к сосняку бруснично-лишайниковозеленомошному, местообитание которого (участок С2) располагается ближе к его центру. Высокое обилие кортицикольных и верхнеподстилочных обитателей, отмеченное на участке С2, сближает население коллембол сосняков бруснично-лишайниково-зеленомошного и вересково-лишайникового. Это вполне закономерно, поскольку под пологом сосновых лесов данных ассоциаций формируются однотипные почвы – подзолы иллювиально-железистые, характеризующиеся маломощным (от 1–2 до 3–5 см) подстилочно-торфяным горизонтом. Следует отметить, что для почвенной мезофауны была выявлена обратная картина [20]. В сосняке лишайниково-зеленомошном по сравнению с двумя другими биотопами, отмечено самое разнообразное и отличающееся высокой численностью сообщество мезофауны.
Анализ населения ногохвосток в разные по погодным условиям годы позволяет заключить, что наиболее четко связь численности и разнообразия сообществ коллембол с условиями гидротермического режима почв прослеживается в прохладные, избыточно влажные годы (как, например, в 2008 г.). Как видно из табл. 3, в 2008 г. в рассмотренном ряду сосняков C1→C2→C3 по мере нарастания влажности почв наблюдается последовательное снижение всех показателей, характеризующих сообщества коллембол (численность, видовое разнообразие, агрегированность). Избыточная влажность почвы сосняка пушицево-багульниково-сфагнового, гидрологический режим которого определяется близостью верхового болота (уровень его почвенно-грунтовых вод напрямую связан с количеством поступающих в летний период осадков и испаряемостью с поверхности почвы), способствует созданию для функционирования комплекса ногохвосток неблагоприятных условий. В аномально теплые годы с недостаточным количеством осадков (2010 г.) экологические условия под пологом сосновых лесов средний тайги, относящихся к различным ассоциациям, выравниваются, и все биотопы характеризуются относительно близкими показателями численности и видового разнообразия коллембол (табл.3). Однако в периоды максимального дефицита влаги (август 2010 г.) наиболее увлажненные почвы сосняка пушицево-багульниково-сфагнового (табл.2), обеспечивают самые благоприятные условия для функционирования сообщества коллембол. В этом биотопе показатели видового разнообразия, численности и агрегированности по сравнению с участками С1 и С2 были несколько выше (табл.3).
Заключение
Проведение комплексных исследований на территории заказника «Белый» показало, что в подзоне средней тайги таксономическое разнообразие коллембол, составляющих население микроартро-под органогенных горизонтов почв сосновых лесов, ниже, чем в ельниках (49 и 59 видов соответственно). На численность коллембол в большей мере оказывают влияние погодные условия вегетационного периода (количество осадков, температура воздуха в летний период), определяющие макро- и микроклиматические условия (влажность и температура почв) конкретных биотопов. В прохладные с большим количеством осадков годы наиболее оптимальные условия для функционирования комплекса ногохвосток складываются под пологом сосняков вересково-лишайниковых. Напротив, в аномально жаркие с минимумом осадков периоды преимущество получают биотопы заболоченных местообитаний, где развиты сосняки пушицево-багульниково-сфагновые. Различия типов почв, формирующихся под сосновыми лесами в подзоне средней тайги, и специфика их гидрологического режима (степень гидроморфизма) находят свое отражение, в первую очередь, в структуре доминирования коллембол и соотношении видов, принадлежащих к различным группам гигропреферендума. На градиенте, отражающем увеличение влажности почв сосновых лесов, наблюдается закономерное уменьшение доли ксерорезистентных ногохвосток и возрастание мезофильных таксонов и видов, не имеющих четко выраженного гигропреферендума. Сообщества заболоченных сосняков включают представителей практически всех групп ногохвосток, в составе которого по численности преобладают мезофилы и виды без четкого гигропрефе-рендума. Это отражает их положение в полосе экотона между лесными и болотными сообществами.
Работа выполнена при поддержке Программы фундаментальных исследований ОБН РАН, проект 12-Т-4-1001 «Структурная организация сообществ почвенных беспозвоночных в лесах таежной зоны Республики Коми».
Список литературы Распределение коллембол (Collembola) в градиенте влажности среднетаежных сосновых лесов (на примере заказника «Белый»)
- Структурно-функциональная роль почв и почвенной биоты в биосфере/Г.В. Добровольский, И.П. Бабьева, Л.Г. Богатырев и др./Отв. ред. Г.В. Добровольский. М.: Наука, 2003. 364 с.
- Кузнецова Н.А. Организация сообществ почвообитающих коллембол. М.: ГНО «Прометей» МПГУ, 2005. 244 с.
- Таскаева А.А., Долгин М.М. Микростациальное распределение коллембол в еловых лесах//Бюлл. МОИП. Отд. биол., 2008. Т.113. Вып. 5. С. 16-22.
- Кузнецова Н.А. Распределение коллембол в сосняках с разным режимом увлажнения в Дарвинском государственном заповеднике//Фауна и экология беспозвоночных животных. М.: Наука, 1984. С. 19-28.
- Кузнецова H.А., Кpестьянинова А.И. Динамика сообществ ногохвосток (Collembola) в гидpологическом pяду южно-таежных сосняков//Зоол. жуpн., 1998. Т. 77. № 9. С.1009-1020.
- Таскаева А.А. Коллемболы сосновых лесов в градиенте загрязнения выбросами лесопромышленного комплекса//Известия ПГПУ им. В.Г. Белинского. 2011. № 25. С. 449-457.
- Kaszmarek M. An analysis of Collembola communities in different pine forest environments//Ecol. Pol., 1975. Vol. 23. № 2. Р. 265-293.
- Hägvar S. Collembola in Norwegian coniferous forest soils. 1. Relations to plant communities and soil fertility//Pedobiologia. 1982. Bd. 24 (5-6). P. 255-296.
- Леса Республики Коми/Под ред. Г.М. Козубова и А.И. Таскаева. М., 1999. С. 134-139.
- Сукачев В.Н., Зонн С.В., Мотовилов Г.П. Методические указания к изучению типов леса. М., 1957. 115 с.
- Нешатаев Ю.Н. Методы анализа геоботанических материалов. Л., 1987. 192 с.
- Ипатов В.С. Описание фитоценоза. Методические рекомендации. СПб., 1998. 93 с.
- Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
- Теория и практика химического анализа почв/Под ред. Л.А. Воробьевой. М.:ГЕОС, 2006. 400 с.
- Чернова Н.М. Распределение панцирных клещей в лесной подстилке//Материалы II акарологического совещания. Киев: Наукова думка, 1970. Ч. 2. С. 203-204.
- Кузнецова Н.А. Влажность и распределение коллембол//Зоол. журн., 2003. Т. 82. № 2. С. 239-247.
- Путеводитель научной почвенной экскурсии. Лесная зона (сезонно-промерзающие почвы). Сыктывкар, 2002. 100 с.
- Краснощекова Е.Н. Комплексы почвенных беспозвоночных среднетаежных сосняков лишайниково-зеленомошных//Ботанические исследования в Сибири. Красноярск, 2005. Вып. 13. С. 101-105.
- Juceviča E., Melecis V. Long-term dynamics of Collembola in a pine forest ecosystem//Pedobiologia, 2002. Bd 46. P. 365-372.
- Конакова Т.Н., Колесникова А.А. Формирование и распределение почвенной мезофауны по градиенту влажности в сосновых лесах Республики Коми//Известия Самарского НЦ РАН. 2011. Т. 13. № 1 (4). С. 1001-1004.
