Распределение молекул межклеточной адгезии на лейкоцитах крови в зависимости от их морфологии

Автор: Шерстенникова Александра Константиновна, Кашутин Сергей Леонидович, Неклюдова Виктория Сергеевна, Калмин Олег Витальевич, Николаев Валентин Иванович
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 2 т.27, 2019 года.
Бесплатный доступ
Физиологическое осмысление процессов миграции иммунокомпетентных клеток, а также условий, при которых может активизироваться или замедляться миграция, является особенно важным для интерпретации резервов стабильности и сохранения гомеостаза. Миграция лейкоцитов через эндотелий микроциркуляции регулируется молекулами адгезии. Цель исследования - изучение распределения молекул межклеточной адгезии на лейкоцитах в зависимости от их морфологии. Проведено обследование 50 практически здоровых лиц в возрасте от 20 до 60 лет, не имеющих хронической патологии в анамнезе. Венозную кровь для исследования брали утром натощак. На проточном цитометре FC-500 фирмы Beckman Coulter (США) определяли количество нейтрофилов, моноцитов и лимфоцитов венозной крови, содержащих молекулы L-селектина (CD62L), LFA-1 (CD11a), LFA-3 (CD58), ICAM-1 (CD54), PECAM-1 (CD31), при этом дифференцировали их в зависимости от морфологии. Результаты свидетельствуют, что более 50% нейтрофилов и 42% моноцитов готовы вступить в фазу скольжения, тогда как количество лимфоцитов, вступающих в данную фазу, сравнительно невелико - 11%...
Клетки крови, лейкоциты, молекулы адгезии, l-селектин
Короткий адрес: https://sciup.org/143177301
IDR: 143177301 | DOI: 10.20340/mv-mn.19(27).02.33-36
The distribution of molecules of the intercellular adhesion on blood leukocytes depending on their morphology
Physiological understanding of the processes of migration of immunocompetent cells, as well as the conditions under which migration can be activated or slowed down, is especially important for the interpretation of the reserves of stability and preservation of homeostasis. Migration of leukocytes through the endothelium of the microcirculation is regulated by adhesion molecules. The aim of the study was to study the distribution of intercellular adhesion molecules on leukocytes depending on their morphology. An examination of 50 healthy individuals aged 20 to 60 years with no history of chronic disease was carried out. Venous blood for the study was taken in the morning on «empty» stomach. On a flow cytometer FC-500 company Beckman Coulter (USA) determined the number of neutrophils, monocytes and lymphocytes of venous blood containing molecule L-selectin (CD62L), LFA-1 (CD11a), LFA-3 (CD58), ICAM-1 (CD54), PECAM-1 (CD31), while differentiated them based on morphology. The results is shown that more than 50% of neutrophils and 42% of monocytes are ready to enter the slip phase, while the number of lymphocytes entering this phase is relatively small - 11%...
Текст научной статьи Распределение молекул межклеточной адгезии на лейкоцитах крови в зависимости от их морфологии
Шерстенникова А.К., Кашутин С.Л., Неклюдова В.С., Калмин О.В., Николаев В.И. Распределение молекул межклеточной адгезии на лейкоцитах крови в зависимости от их морфологии// Морфологические ведомости.- 2019.- Том 27.- № 2.- С. 33-36. (27).02.33-36
Sherstennikova AK, Kashutin SL, Neklyudova VS, Kalmin OV, Nikolaev VI. The distribution of molecules of the intercellular adhesion on blood leukocytes depending on their morphology. Morfologicheskie Vedomosti – Morphological Newsletter. 2019 August 5;27(2):33-36. (27).02.33-36
Введение. Миграция лейкоцитов через эндотелий микроциркуляции регулируется молекулами адгезии. Процесс миграции лейкоцитов в ткани представляет собой трехступенчатый процесс, включающий скольжение – «роллинг» лейкоцитов по поверхности эндотелия с последующей фазой прочной адгезии и заканчивающийся фазой трансмиграции лейкоцитов через эндотелий [1-2]. Роллинг лейкоцитов опосредуется низкоаффинными рецепторами – селектинами, в том числе L-селектинами [3]. Фаза прочной адгезии реализуется с участием молекул ICAM-1, LFA-1, LFA-3 [4]. Собственно миграция лейкоцитов через эндотелий связана с экспрессией молекул PECAM [5]. Сведения о концентрациях молекул адгезии на нейтрофилах, моноцитах и лимфоцитах в условиях отсутствия антигенной нагрузки единичны и противоречивы [6-7]. В литературе нет сведений относительно уровня экспрессии молекул адгезии в зависимости от морфологических особенностей нейтрофилов, моноцитов и лимфоцитов. Таким образом, следует признать, что литературные данные по этому вопросу явно недостаточны, а сама проблема находится в стадии накопления фактов [8]. Учитывая значительную физиологическую роль процессов миграции лейкоцитов из русла микроциркуляции в ткани, сведения о концентрации молекул адгезии на нейтрофилах, моноцитах и лимфоцитах с учетом их морфологических особенностей представляют теоретический и практический интерес [9]. Физиологическое осмысление процессов миграции иммунокомпетентных клеток, а также условий, при которых может активизироваться или замедляться миграция, особенно важно для интерпретации резервов стабильности и сохранения гомеостаза [10-11]. Все перечисленное определяет актуальность, перспективность и практическую значимость научного поиска регуляции миграционной активности иммунокомпетентных клеток [12-14].
Цель исследования – изучение распределения молекул межклеточной адгезии на лейкоцитах в зависимости от их морфологии.
Материал и методы исследования. Проведено обследование 50 практически здоровых лиц (22 мужчин и 28 женщин) в возрасте от 20 до 60 лет, не имеющих хронической патологии в анамнезе. Обследование проводили с письменного согласия респондентов, с соблюдением основных норм биомедицинской этики в соответствии с документом «Этические принципы проведения медицинских исследований с участием людей в качестве субъектов исследования» (Хельсинкская декларация Всемирной медицинской ассоциации 1964 года с изменениями и дополнениями на 2008 год). Венозную кровь для исследования брали утром натощак. На проточном цитометре FC-500 фирмы Beckman Coulter (США) определяли количество нейтрофилов, моноцитов и лимфоцитов венозной крови, содержащих молекулы L-селектина (CD62L, FITC), LFA-1 – Lymphocyte Function-Assosiated antigen, 1 (CD11a, FITC), LFA-3 – Lymphocyte Function-Assosiated antigen, 3 (CD58, FITC), ICAM-1 – Intercellular Adhesion Molecule-1 (CD54, FITC), PECAM-1 – Platelet/Endothelium Cell Adhesion Molecule (CD31, FITC). На мазке крови, зафиксированном смесью Никифорова и окрашенном по Романовскому-Гимзе, определяли содержание нейтрофилов, эозинофилов, базофилов, моноцитов, лимфоцитов [15]. Цитоскопическое исследование нейтрофильных лейкоцитов проводили путем подсчета среднего количества фрагментов ядра у 100 клеток [16]. В соответствии с методом, предложенным Григоровой [17], проводили дифференцировку моноцитов по морфологии ядра на промоноциты, собственно моноциты и полиморфноядерные моноциты. При изучении лимфоцитограммы дифференцировали лимфоциты по величине клетки с учетом размеров цитоплазмы: малые лимфоциты – до 8 мкм, средние – от 8 до 12 мкм, большие – больше 12 мкм. Отдельно выделяли популяцию больших гранулярных лимфоцитов, которые являются морфологическими аналогами естественных киллеров [18]. Статистическую обработку результатов проводили с помощью SPSS 13.0 for Windows. Распределение параметров было ненормальным, в связи с чем описание выборок проводили с помощью подсчета медианы (Мe) и межквартильного интервала (25 и 75 процентили). Вероятность различий оценивали по непараметрическим критериям Колмогорова-Смирнова (Z) и Вилкоксона (W). Корреляционный анализ проводили с использованием коэффициента корреляции Спирмена ( ρ ).
Результаты исследования и обсуждение. В условиях отсутствия антигенной стимуляции общее содержание нейтрофилов в крови составило 3,9 · 109/л, моноцитов 0,35 · 109/л, лимфоцитов 2,56 · 109/л. Количество нейтрофилов, имеющих рецептор L-селектина составило – 65% (40,9; 87,32), моноцитов – 42,2% (14,82; 60,0), при этом лимфоцитов с данным рецептором был невелик и составил 11,42% (4,5; 23,38). Таким образом, в условиях отсутствия антигенной стимуляции более 50% нейтрофилов готовы вступить в фазу скольжения, тогда как количество лимфоцитов, вступающих в данную фазу, сравнительно невелико. Практически все нейтрофилы - 99,35% (93,56; 100,00) и моноциты - 99,72% (92,75; 100,0) содержали молекулу LFA-1. Количество лимфоцитов с данной молекулой составил 67,97% (44,71; 77,69). Наиболее часто молекулу ICAM-1 выявляли на моноцитах - 88,88% (61,33%; 96,83), в меньшей степени на нейтрофилах - 55,74% (31,45; 71,81) и только на 11,56% (4,37; 21,6) лимфоцитах. Уровень нейтрофилов и моноцитов с молекулой LFA-3 в периферической крови был одинаков и составил 94,11% (70,25; 98,29) и 94,11% (77,08; 100,0), соответственно. Только на 33,02% (12,66; 82,6) лимфоцитах определяли данную молекулу. Молекулу PECAM-1 практически одинаково часто выявляли на нейтрофилах и моноцитах: 98,72% (92,09; 99,53) и 97,5% (80,0; 98,75), соответственно. Только 12,56% (3,7; 28,6) лимфоцитов несли данный рецептор (рис. 1).
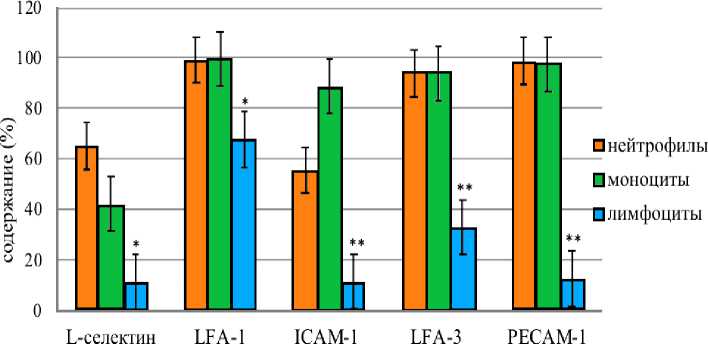
молекулы адгезии
Рис. 1. Распределение молекул адгезии на нейтрофилах, моноцитах и лимфоцитах % (Me). Статистически значимые различия *p<0,05 и при **p<0,001.
Таким образом, более чем 90% нейтрофилов и моноцитов в условиях отсутствия антигенной стимуляции готовы вступить в фазу прочной адгезии и трансмиграции. Содержание лимфоцитов в условиях отсутствия антигенной нагрузки, готовых к трансмиграции, не превышало 13%.
Анализ сегментации ядер клеток показал, что в венозной крови преобладали нейтрофилы с 3-мя (1,37·109/л; 1,04-1,81) и 4-мя (1,13·109/л; 0,9- 1,59) сегментами в ядре. Концентрации нейтрофилов с 1-м, 2-мя, 5-ю и более сегментами были сравнительно небольшие.
Корреляционный анализ между количеством нейтрофилов, несущих молекулы адгезии, и сегментацией их ядер показал наличие слабых и статистически недостоверных корреляций в случаях, когда нейтрофилы содержали 1, 2 и 3
сегмента в ядре. Статистически значимые корреляции были выявлены у 4-сегментных форм нейтрофилов: слабые - с нейтрофилами, содержащими молекулу L-селектина ( р =0,28; р=0,04), средней силы - с нейтрофилами, содержащими молекулы LFA-1 ( р =0,37; р=0,009), LFA-3 ( р =0,49; р=0,003) и сильные - при выявлении на нейтрофилах молекулы PECAM-1 ( р =0,52; р=0,01). В случаях с нейтрофилами, содержащими пять и более сегментов в ядре, все эти корреляции стали сильными и статистически значимыми: L-селектин (- р =0,51; р=0,003), LFA-1 (- р =0,55; р=0,0002), LFA-3 (- р =0,62; р=0,008), PECAM-1 (- р =0,60; р=0,002), ICAM-1 ( р =0,53; р=0,001).
Анализ структуры моноцитограммы показал, что количество промоноцитов составил 26,0% (20,0; 36,0), собственно моноцитов 40,0% (29,5; 47,0), полиморфно-ядерных моноцитов - 31,0% (24,5; 43,0) без существенных половых различий. Корреляционный анализ между содержанием моноцитов, несущих молекулы адгезии, и их различием по форме ядра показал, что в ходе дальнейшей дифференцировки ядра моноцитов появляются новые статистически достоверные корреляции. Если содержание промоноцитов коррелировало только с экспрессией молекул LFA-1 и ICAM-1 ( р =0,39; р=0,005 и р =0,43; р =0,002), то у собственно моноцитов появилась корреляция с концентрацией моноцитов, содержащих молекулу LFA-3 ( р =0,38; р=0,03), а у полиморфно-ядерных моноцитов еще и содержанием моноцитов с молекулой PECAM-1 ( р =0,45; р=0,03).
Малые лимфоциты в структуре лимфоцитограммы составили 26,0% (16,0; 37,0), средне-плазменные - 44,0% (36,0; 50,0), большие - 10,0% (5,0; 16,5), большие гранулярные лимфоциты - 16,0% (12,0; 23,5). Корреляционный анализ между содержанием лимфоцитов, несущих молекулы адгезии, и размером их цитоплазмы показал, что при увеличении размеров цитоплазмы лимфоцитов увеличивается количество статистически достоверных корреляций. Так, если содержание малых лимфоцитов коррелировало только с содержанием лимфоцитов с молекулой LFA-1 ( p =0,38; р=0,03), то концентрация среднеплазменных лимфоцитов была связаны с содержанием молекул LFA-1 ( p =0,57; р=0,001), LFA-3 ( p =0,43; р =0,016) и L-селектина ( р =0,35; р=0,013). Подобную ситуацию регистрировали также в отношении больших гранулярных лимфоцитов, тогда как между уровнем больших лимфоцитов и лимфоцитов, экспрессирующих изучаемые молекулы адгезии, статистически значимых корреляций не выявлено.
Если учитывать, что в сосудистом русле существуют два почти равных пула нейтрофилов - циркулирующий и пристеночный [19] и при заборе венозной крови считается только циркулирующий пул, можно полагать, что в условиях отсутствия антигенной стимуляции 65% нейтрофилов циркулирующего пула несут молекулы L-селектина, 99,35% - молекулы LFA-1, 55,74% - ICAM-1, 94,11% - LFA-3, 98,72% - PeCaM-1. Если учитывать, что L-селектин участвует в механизме формирования пристеночного пула, обеспечивая роллинг-эффект, можно полагать, что 65% циркулирующих нейтрофилов способны пополнить пристеночный пул [10]. Известно, что адгезия, вызванная селектинами, обратима, кратковременна и малоэффективна. Более прочную и необратимую адгезию нейтрофилов на эндотелии обусловливают В 2 -интегрины, к которым относится молекула LFA-1 [15, 19]. В соответствии с результатами исследования 99,35% нейтрофилов циркулирующего пула имели эту молекулу, что несколько больше, чем количество нейтрофилов с молекулой L-селектина (65%). Учитывая возможность протеолитического отщепления молекулы L-селектина при экспрессии на нейтрофилах 6 2 -интегринов на клеточной поверхности, что определяется как шеддинг-феномен, можно предполагать, что шеддинг молекул L-селектина проявляется среди нейтрофилов циркулирующего пула достаточно активно [16].
Циркулирующие нейтрофильные лейкоциты имеют на своей поверхности молекулы адгезии: ICAM-1, LFA-3, PECAM-1. Учитывая, что транс-эндотелиальная миграция нейтрофилов опосредована, в том числе этими молекулами, можно полагать, что у нейтрофилов циркулирующего пула имеются все необходимые возможности для трансмиграции [5, 14]. В соответствии с результатами исследования установлено отсутствие корреляций между концентрацией нейтрофилов, имеющих молекулы межклеточного взаимодействия с содержанием нейтрофилов с 1-, 2- и 3- сегментами в ядре. Наличие корреляций с 4-мя, а тем более с 5-сегментными формами, позволяет предполагать, что при отсутствии антигенной стимуляции молекулы адгезии начинают выявляться на 4-сегментных формах, тогда как в полной мере этот процесс проявляется у нейтрофилов с 5-ю и более сегментами в ядре.
В условиях отсутствия антигенной стимуляции 42,2% моноцитов имеют молекулы L-селектина, 99,72% молекулы LFA-1, 88,88% - ICAM-1, 94,11% - LFA-3, 97,50% - PECAM-1. В соответствии с результатами исследования практически все моноциты (99,72%) несли молекулу LFA-1, что в 2 раза больше, чем число моноцитов, содержащих молекулу L-селектина (42,2%). Можно полагать, что шеддинг молекул L-селектина среди моноцитов протекает достаточно активно, и значительно активнее, чем у нейтрофилов. С другой стороны, более низкая экспрессия молекул L-селектина на моноцитах при увеличении на них экспрессии молекулы ICAM-1 может быть связана с увеличением секреции TNF-a и активизации цитотоксичности моноцитов [13, 20].
Содержание моноцитов периферической крови, несущих молекулы адгезии ICAM-1, LFA-3, PECAM-1, был чрезвычайно высоким в сравнении с концентрацией нейтрофилов, несущих молекулы межклеточного взаимодействия, на основании чего можно полагать, что процессы трансмиграции, особенно полиморфно-ядерных моноцитов, в условиях отсутствия какой-либо антигенной стимуляции достаточно активны. Подтверждением этому может служить наличие статистически достоверных корреляций между содержанием полиморфно-ядерных моноцитов и уровнем моноцитов, имеющих молекулы адгезии, участвующих во всех этапах миграции, в фазе скольжения, прочной адгезии и трансмиграции.
В условиях отсутствия антигенной стимуляции 11,42% лимфоцитов содержали молекулы L-селектина, 67,97% молекулы LFA-1,11,56% - ICAM-1,33,02% - LFA-3,12,56% - PECAM-1. В соответствии с результатами исследования большая часть лимфоцитов (67,97%) экспрессировали молекулу LFA-1, что в 6 раз больше по сравнению с лимфоцитами, содержащими молекулу L-селектина (11,42%). Вероятно шеддинг молекул L-селектина среди лимфоцитов протекает достаточно активно, и значительно активнее, чем у нейтрофилов и моноцитов. Кроме того, учитывая статистически достоверные корреляции между уровнем средне-плазменных лимфоцитов, а также больших гранулярных лимфоцитов и концентрацией лимфоцитов, несущих молекулы LFA-1 и LFA-3, можно предполагать, что к миграции более предрасположены средне-плазменные лимфоциты и большие гранулярные лимфоциты, идентифицированные как естественные киллеры [18, 21]. С учетом того, что уровень больших лимфоцитов может свидетельствовать о лимфо-пролиферации, создается впечатление, что при лимфо-пролиферации лимфоциты обладают меньшей способностью к адгезии, а, следовательно, и трансмиграции [3, 11].
Заключение . Итак, в условиях отсутствия антигенной стимуляции наибольшая готовность к реализации фазы скольжения, прочной адгезии и непосредственно трансмиграции наблюдалась у нейтрофилов с 5-сегментами в ядре и полиморфно-ядерных моноцитов. Готовность к трансмиграции лимфоцитов была сравнительно невелика и касалась, в основном, средне-плазменных лимфоцитов и больших гранулярных лимфоцитов.
Список литературы Распределение молекул межклеточной адгезии на лейкоцитах крови в зависимости от их морфологии
- Smith CJ. Adhesion molecules and receptors. J Allergy Clin Immunol. 2008;3(2):375-379.
- Wallez Y, Huber P. Endothelial adherens and tight junction in vascular homeostasis, inflammation and angiogenesis. J Biochim Biophys Acta. 2008;1778(3):794-809.
- Vestweber D. Lymphocyte trafficking through blood and lymphatic vessels: more than just selectins, chemokines and integrins. J Eur J Immunol. 2003;33(5):1361-1364.
- Scheglovitova O, Scanio V, Fais S et al. Antibody to ICAM-1 mediates enhancement of HIV-1 infection of human endothelial cells. J Arch Virol. 1995;140(5):951-958.
- Murciano JC, Muro S, Koniaris L et al. ICAM-directed vascular immunotargeting of anti-thrombotic agents to the endothelial surface. J Blood. 2003;101(10):3977-3984.
- Cinkernagel' R. Osnovy immunologii.- M.: Mir, 2008.- 136s.
- Vestweber D. Endothelial cell contacts in inflammation and angiogenesis. J Int Congr Ser. 2007;1302:17-25.
- Filippova O.E., Popovskaya E.V., Shashkova E.YU., Shchegoleva L.S. Aktivnost' limfoidnyh receptorov limfoproliferacii i apoptoza v fiziologicheskoj regulyacii immunnogo gomeostaza pri adaptacii cheloveka k menyayushchimsya usloviyam sredy (obzor). Vestnik Severnogo (Arkticheskogo) federal'nogo universiteta. Seriya: Mediko-biologicheskie nauki. 2016;1;87-99.
- Pal'cev M.A. Mezhkletochnye vzaimodejstviya.- M.: Medicina, 1995.- 224s.
- Dolgushin I.I., Buharin O.V. Nejtrofily i gomeostaz.- Ekaterinburg, 2001.- 136s.
- Bajgil'dina A.A., Galieva A.T., Kamilov F.H. Patogeneticheskoe znachenie nekotoryh citokinov i belka kletochnoj adgezii VCAM*1 v razvitii vospalitel'nyh izmenenij ehndoteliya sosudov pri gemorragicheskoj lihoradke s pochechnym sindromom. Morfologicheskie vedomosti. 2008;3-4:158-161.
- Hayashida K, Bartlett AH, Chen Y, Park PW. Molecular and cellular mechanisms of ectodomain shedding. J Anat Rec (Hoboken). 2010;293(6):925-937.
- Reymond N. DNAM-1 and PVR regulate monocyte migration through endothelial junction. J Exp Med. 2004;199(10):1331-1341.
- Witkowska AM, Borawska MH. Soluble intercellular adhesion molecule-1 (sICAM-1): an overview. J Eur Cytok Netw. 2004;15(2):91-98.
- Pinegin B.V. Nejtrofily: struktura i funkciya. Immunologiya.- 2007;28(6):374-382.
- Todorov J.T. Klinicheskie laboratornye issledovaniya v pediatrii.- Sofiya, 1968.- 784s.
- Grigorova O. P. Rol' monocitarnoj sistemy v reaktivnosti organizma.- M.: Medgiz, 1958.- 105s.
- Zak K.P., Kindzel'skij L.P., Butenko A.K. Bol'shie granulosoderzhashchie limfocity i patologicheskie processy.- Kiev: Naukova dumka, 1992.- 205s.
- Garton KJ, Gough PI, Raines EW. Emerging role for ectodomain shedding in the regulation of inflammatory responces. J Leukem Biol. 2006;79(6):1105-1679.
- Rauert H, Wicovvsky A, Miller N et al. Membrane tumor necrosis factor (TNF) induces p100 processing via TNF receptor-2 (TNFR2). J Biol Chem Mar. 2010;285(10):7394-7404.
- Camagna A, Cedrone L, Care A et al. Polyclonal expansion of CD3(+)/CD4(+)/CD56(+) large granular lymphocytes and autoimmunity associated with dysregulation of Fas/FasL apoptotic pathway. Br J Haematol. 2001:112(1):204-207.
