Распространение и разнообразие сульфатредуцирующих бактерий в слабоминерализованных щелочных гидротермах Байкальской рифтовой зоны

Автор: Бархутова Д.Д., Раднагуруева А.А., Данилова Э.В.
Журнал: Природа Внутренней Азии @nature-inner-asia
Рубрика: Биология
Статья в выпуске: 3 (25), 2023 года.
Бесплатный доступ
В слабоминерализованных щелочных гидротермах Байкальской рифтовой зоны создаются условия для развития термофильных и мезофильных сульфатредуцирующих бактерий. Исследовано разнообразие и распространение сульфатредукторов с помощью культуральных и молекулярных подходов. Сульфидогенное сообщество представлено бактериями классов Nitrospira, Deltaproteobacteria, Clostridia и Thermodesulfobacteria. Среди них многочисленными были термофильные бактерии рода Thermodesulfovibrio (класс Nitrospira) и мезофильные бактерии рода Desulfarculus (класс Deltaproteobacteria), редкими - бактерии родов Desulfobacca, Desulfococcus (класс Deltaproteobacteria), Desulfotomaculum, Thermodesulfobium, Desulfurispora (класс Clostridia). Сульфидогенное сообщество участвует в восстановительной фазе цикла серы при температуре 30-75 °С в диапазоне рН от 7 до 10,5 в условиях щелочных гидротерм Байкальской рифтовой зоны. Полученные данные свидетельствуют о том, что многочисленные и редкие таксоны сульфатредуцирующих бактерий могут занимать разные экологические ниши и вместе сохранять экологическую значимость сульфатредуцирующих бактерий в щелочных гидротермах Байкальской рифтовой зоны.
Сульфатредуцирующие бактерии, бактериальная сульфатредукция, щелочные гидротермы, байкальская рифтовая зона
Короткий адрес: https://sciup.org/148328086
IDR: 148328086 | УДК: 579.26 | DOI: 10.18101/2542-0623-2023-3-17-30
Distribution and diversity of sulfate-reducing bacteria in weakly mineralized alkaline hydrotherms of Baikal rift zone
Weakly mineralized alkaline hydrotherms of the Baikal Rift Zone (BRZ) create favourable conditions for the development of thermophilic and mesophilic sulfate-reducing bacteria. We have studied the diversity and distribution of sulfate reducers using cultural and molecular approaches. The sulfidogenic community is represented by bacteria of the classes Nitrospira, Deltaproteobacteria, Clostridia and Thermodesulfobacteria. These are numerous thermophilic bacteria of Thermodesulfovibrio genus (class Nitrospira) and mesophilic bacteria of Desulfarculus genus (class Deltaproteobacteria), rare bacteria of Desulfobacca, Desulfococcus (class Deltaproteobacteria), Desulfotomaculum, Thermodesulfobium, Desulfurispora (class Clostridia) genera. In the alkaline conditions of thermal springs thermophilic and mesophilic sulfate-reducing bacteria participate in the reducing phase of the sulfur cycle in the pH range from 7 to 10,5 and temperatures from 30 to 75 °С. The data obtained indicate that numerous and rare SRB taxa can occupy different ecological niches and together maintain the ecological significance of sulfate-reducing bacteria in alkaline hydrotherms of the BRZ.
Текст научной статьи Распространение и разнообразие сульфатредуцирующих бактерий в слабоминерализованных щелочных гидротермах Байкальской рифтовой зоны
Бактериальный процесс сульфатредукции является ключевым звеном в глобальном биогеохимическом цикле серы и углерода и играет важную роль на терминальных этапах анаэробной деструкции органического вещества [Заварзин, 2003]. Сульфатредуцирующие прокариоты, восстанавливающие сульфат, универсальны с точки зрения условий роста и могут развиваться в самых разных условиях окружающей среды. Они получают энергию за счет переноса электронов между различными донорами (органическими соединениями, водородом или окисью углерода) с сульфатом, образуя сероводород [Muyzer and Stams, 2008]. Экстремальные гидротермальные системы являются источником сульфатредукторов, которые могут расти при повышенных температурах и различных значениях pH.
С помощью культуральных и молекулярных подходов на основании нуклеотидных последовательностей 16S рРНК и с использованием гена сульфитредук-тазы dsrAB установлено большое разнообразие сульфатредукторов. Сульфатре-дуцирующие прокариоты включают представителей четырех таксономических групп: 1) дельтапротеобактерии, содержащие разнообразные роды грамотрица-тельных мезофильных сульфатредуцирующих бактерий (СРБ); 2) грамположи-тельные спорообразующие СРБ, устойчивые к более высоким температурам, чем мезофильная группа (между 40 °C и 60 °C) [Aüllo et al., 2013]; 3) термофильные СРБ, обитающие в условиях высокой температуры, с оптимальной температурой роста от 65 °C до 70 °C; 4) термофильные сульфатредуцирующие археи, которые распространены при температуре выше 80° [Rabus et al., 2015].
Большое количество горячих источников с различными физико-химическими свойствами широко распространены в Байкальской рифтовой зоне (БРЗ) [Борисенко, Замана, 1978]. Слабоминерализованные (минерализация от 200 до 920 мг/л) щелочные (рН от 8 до 10) гидротермы с температурой 41–90 °С являются наиболее известными среди них. Исследование изотопного состава сульфатной и сульфидной серы в азотных щелочных гидротерм БРЗ показало, что сульфидная сера обеднена тяжелым изотопом по сравнению с сульфатами, что возможно при бактериальной сульфатредукции [Замана и др., 2010].
Ранее приведенные исследования функционального разнообразия микробного сообщества и интенсивности биогеохимических процессов в микробных матах и осадках гидротерм БРЗ показали, что сульфатредукция является одним из ведущих процессов на терминальных этапах деструкции органического вещества [Намсараев и др., 2006; Лаврентьева и др., 2019: Бархутова и др., 2021]. Высокая скорость процесса сульфатредукции и образование сероводорода позволяют сделать вывод об важной роли сульфатредуцирующих бактерий в процессах анаэробной деструкции органического вещества и активной деятельности бактерий цикла серы в гидротермах.
Анализируя литературные данные, следует отметить слабую изученность сульфатредуцирующих бактерий щелочных гидротерм БРЗ. Настоящая работа была направлена на выявление СРБ из анаэробных зон слабоминерализованных щелочных гидротерм БРЗ с помощью культуральных и молекулярных подходов.
Исследование СРБ, обитающих в горячих источниках при щелочных значениях рН, является важным для расширения знаний о структуре и экологических функциях микробного сообщества в экосистемах горячих источников. Выявленные культивируемые бактерии открывают новые возможности для применения в биотехнологических процессах.
Методы исследований
В полевых условиях в местах отбора проб определялись температура, рН, минерализация, концентрация карбонат- и гидрокарбонат-ионов. В лабораторных условиях определяли сульфаты, хлориды, катионы натрия, калия, кальция, магния, сульфиды [Намсараев и др., 2005].
Учет численности и выделение мезофильных и термофильных сульфатредуци-рующих бактерий проводили на модифицированной среде Постгейта [Кузнецов, Дубинина, 1989]. В качестве индикатора процесса сульфатредукции использовалась металлическая скрепка или FeSO4, в качестве доноров электронов источника углерода — натриевые соли молочной или уксусной кислот. Для выявления мезофильных и термофильных СРБ инкубацию проводили при 30 °С и 50 °С соответственно.
ДНК выделяли с помощью коммерческого набора Genomic DNA from Soil (Macherey-Nagel, Германия). Для амплификации фрагментов гена 16S рРНК использовались праймеры 343F и 806R [Dubovskiy et al., 2016]. Секвенирование библиотеки на платформе MiSeq проводилось по протоколу производителя в ЦКП «Геномика» (ИХБФМ СО РАН, г. Новосибирск).
Результаты и обсуждение
Физико-химическая характеристика гидротерм
Объектами исследования были слабоминерализованные щелочные гидротермы БРЗ — Сеюя, Кучигер, Умхэй, Алла, Гарга, Уро, Гусиха, Горячинск.
Температура воды на изливе варьировала от 41 до 79 °С. Термальные воды имели щелочную реакцию — pH варьировал от 8,2 до 9,9. В основном для исследуемых гидротерм характерны сульфатно-гидрокарбонатные натриевые воды и восстановленные условия. По содержанию сероводорода исследуемые гидротермы подразделяются на сульфидные (концентрация H2S 1-26,3 мг/дм3) и бессульфидные (H2S ≤ 0,5 мг/дм3).
В таблице 1 показано изменение физико-химических параметров в горячих ручьях, которые образуются при выходе источника на земную поверхность.
Распространение сульфатредукторов в гидротермах
В осадках слабоминерализованных щелочных гидротерм была определена численность мезофильных и термофильных СРБ. При использовании в качестве единственного источника углерода и донора электронов лактата и ацетата численность термофильных достигала 106–108 кл/см3, максимальное количество было обнаружено в осадках гидротермы Горячинск. Численность мезофильных СРБ не превышала 106 кл/см3. Высокие численности термофильных и мезофильных СРБ выявлены в осадках гидротермы Кучигер и Горячинск. Численность СРБ в осадках из гидротерм Гарга, Сеюя, Гусиха и Алла не превышала 106 кл/см3 (табл. 2).
При этом в пробах осадков источников Сеюя и Алла численность сульфа-тредукторов, растущих на лактате, была выше, чем на ацетате.
Таблица 1
Характеристика физико-химических параметров (по изливу ручья источников)
|
Источник |
Станция |
t , ºС |
рН |
Eh, мВ |
H2S, мг/дм3 |
|
Алла |
Б-33 |
75,8 |
9,1 |
-172 |
11,4 |
|
Б-33с |
53 |
8,93 |
-100 |
1,3 |
|
|
Б-34 |
74 |
9 |
-178 |
8,4 |
|
|
Б-35 |
76,4 |
9,06 |
-154 |
18,4 |
|
|
Б-6 |
53 |
9,1 |
-150 |
16 |
|
|
Сеюя |
1 |
49 |
9,75 |
-96 |
2,5 |
|
2 |
45 |
9,74 |
-84 |
0,4 |
|
|
3 |
43 |
9,7 |
-20 |
0,01 |
|
|
Гусиха |
1 |
72 |
8,5 |
340 |
0,01 |
|
2 |
43 |
8,5 |
340 |
0,01 |
|
|
Гарга |
1 |
74 |
8,5 |
20 |
0,01 |
|
8 |
73 |
8,3 |
230 |
0,01 |
|
|
Горячинск |
1 |
51,1 |
8,95 |
-63 |
1,0 |
|
2 |
50 |
8,7 |
-63 |
1,0 |
|
|
3 |
47,6 |
8,72 |
60 |
1,0 |
|
|
4 |
19,3 |
7,43 |
100 |
1,0 |
|
|
Сухая |
1 |
48,4 |
7,92 |
-95 |
19,37 |
|
2 |
38,7 |
8,13 |
-66 |
7,77 |
|
|
Кучигер |
1 |
34 |
9,56 |
-180 |
23,0 |
|
2 |
41 |
9,72 |
-17 |
23,2 |
|
|
3 |
39,6 |
9,6 |
-70 |
16,5 |
|
|
4 |
33 |
9,85 |
-43 |
16,2 |
|
|
5 |
24,7 |
9,53 |
-40 |
26,3 |
Таблица 2
Численность СРБ в донных осадках гидротерм
|
Источник |
t °С |
рН |
Термофильные СРБ, кл/см3 |
Мезофильные СРБ, кл/см3 |
||
|
ацетат |
лактат |
ацетат |
лактат |
|||
|
Кучигер, 07 |
24–21 |
9,53–9,85 |
107 |
107 |
106 |
106 |
|
Сеюя, 08 |
49–33 |
9,1–9,4 |
103–105 |
104–106 |
103 –104 |
104–105 |
|
Горячинск, 08 |
49 |
9,0 |
106 |
108 |
104 |
105 |
|
Гарга, 08 |
74–45 |
8,1–8,2 |
103 –104 |
105–106 |
103 –104 |
103–105 |
|
Алла, 08 |
76–43 |
8,9–9,1 |
104–105 |
104–106 |
103 |
104–105 |
|
Гусиха, 08 |
63–43 |
8,1–8,2 |
103 |
104 |
103–104 |
104–105 |
В гидротерме Умхэй численность термофильных СРБ не превышала 105 кл/ см3. Максимальная численность сульфатредукторов выявлена на глубине 6–7 см, при этом концентрация сульфидов осадках достигала 22,7 мг/дм3. Выявлена корреляция между распределением СРБ и содержанием сульфидов в осадках гидротермы Умхэй (рис. 1).
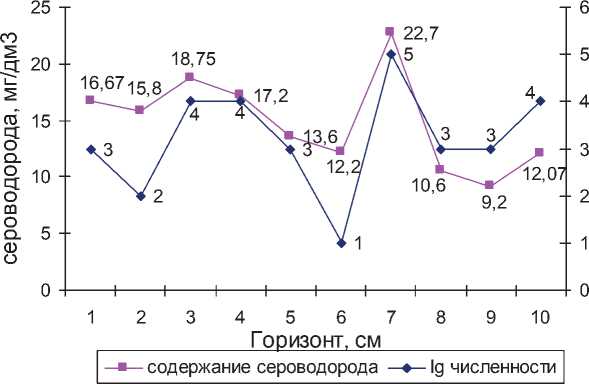
Рис. 1. Содержание H2S и lg численности СРБ в донных осадках источника Умхэй
Разнообразие сульфатредукторов
Микробное разнообразие гидротерм БРЗ, изученное нами с использованием высокопроизводительного секвенирования гена 16S, позволило выявить культивируемые и некультивируемые виды бактерий различных физиологических групп в микробных сообществах микробных матов и осадков [Лаврентьева и др., 2017, 2019; Бархутова и др., 2021; Будагаева и др., 2021]. Высокая доля термофильных бактерий филумов Acetothermia (до 58 %) и Deinococcus-Thermus (до 50 %) была выявлена в микробном сообществе в высокотемпературной зоне гидротерм Гарга и Алла. Экстремально термофильные представители филума Aquificae составляли до 10 %, археи типа Crenarchaeota — 4 % всего прокариотного сообщества. Представители Alphaproteobacteria , Bacteroidetes, Actinobacteria доминировали в осадках и в воде при снижении температуры. В микробных матах при высокой температуре доминировали аноксигенные бактерии Chloroflexi, при снижении температуры — оксигенные бактерии Cyanobacteria . В таблице 3 приведены доминирующие типы бактерий в микробном сообществе осадков (s) и микробных матов (m) гидротермы Алла.
Ранее нами было показано, что сульфатредукторы занимают значительную долю и могут составлять более трети эубактериального микробного сообщества (до 36 %) в осадках гидротермы Алла при температуре 73 °С [Раднагуруева и др., 2016]. Выявленные нуклеотидные последовательности сульфатредуцирующих бактерий, способных к диссимиляционной сульфатредукции, были родственны к культивируемым и некультивируемым термофильным бактериям рода Thermodesulfovibrio филотипа Nitrospirae.
Таблица 4
Многочисленные филотипы микробных сообществ гидротермы Алла
|
Источник, тип пробы |
t отбора, °С |
pH |
Минерализация, г/л |
Основные филотипы |
|
Al-1w Алла вода |
64,4 |
9,7 |
0,25 |
Deinococcus-Thermus 50 %, Chloroflexi 9,9 %, Aquifica e 7,7 % Gammaproteobacteria 5,9 %, Cyanobacteria/Chloroplast 5,4 % |
|
Al-0 Алла осадки |
68 |
9,8 |
0,28 |
Acetothermia 31,6 %, Firmicutes 28,6 %, Gammaproteobacteria 16 % |
|
Al-3s Алла осадки |
64 |
9,7 |
0,26 |
Acetothermia 57,9 %, Chloroflexi 6,3 %, Nitrospirae 4,6 % Crenarchaeota - 4 % |
|
Al-3w Алла вода |
58,1 |
9,6 |
0,34 |
Cyanobacteria/Chloroplast 70,7 %, Gammaproteobacteria 8 %, Chloroflexi 4,5 % |
|
Al-1m Алла мат |
41 |
9,6 |
0,25 |
Atribacteria 28,4 %, Nitrospirae 27 %, Chloroflexi 16,2 %, Alphaproteobacteria 7 %, Deinococcus-Th ermus (4,9 %) |
|
Al-3m Алла мат |
58,1 |
9,93 |
0,34 |
Chloroflexi 56 %, Nitrospirae 9,7 %, Deltaproteobacteria 2,1 %, Firmicutes 2,1 % |
|
Al-5m Алла мат |
34,4 |
9,26 |
0,35 |
Deltaproteobacteria 27 % , Gammaproteobacteria 26,4 %, Alphaproteobacteria 16,3 % Acidobacteria 17,5 % |
С использованием высокопроизводительного параллельного секвенирования (NGS) гена 16S рРНК было получено 27 506 и 9 483 нуклеотидных последовательностей в микробном мате и иловых отложениях гидротермы Алла соответственно. В таблице 4 представлены представители СРБ, выявленные в микробном сообществе Алла, которые относились к классам Nitrospira, Deltaproteobacteria, Clostridia и Thermodesulfobacteria. Бактериальные сообщества, как правило, неравномерны по численности и распределению: несколько видов с высокой численностью (многочисленные таксоны) и большинство с низкой численностью (редкие таксоны) [Pedrós-Alió, 2012].
Таблица 4
Наблюдаемые таксоны СРБ в микробных матах и осадках гидротермы Алла
|
Образцы |
Таксоны СРБ на уровне рода в порядке убывания численности (%) |
|
Микробный мат Al-1m (64,4 ºC, pH 9,7) |
Thermodesulfovibrio (36,2 %) Desulfarculus (0,5 %) Desulfovirgula (0,4 %) Desulfomonile (0,1 %) Desulfobacca (0,025) Desulfococcus (0,02 %) Desulfotomaculum (0,01 %) |
|
Осадки Al-1s (64,4 ºC, pH 9,7) |
Thermodesulfovibrio (0,6 %) Desulfovirgula (0,03 %) Thermodesulfobium (0,03 %) |
|
Микробный мат Al-3m (58,1ºC, pH 9,6) |
Thermodesulfovibrio (40 %) Desulfarculus (0,7 %) Thermodesulfobacterium (0,08 %) Desulfomonile (0,05 %) Desulfobacca (0,02 %) Desulfococcus (0,01 %) Desulfobulbus (0,01 %) |
|
Осадки Al-3s (58,1ºC, pH 9,6) |
Thermodesulfovibrio (14 %) Desulfarculus (1,1 %) Desulfovirgula (0,3 %) Thermodesulfobacterium (0,3 %) Desulfomonile (0,06 %) Desulfococcus (0,03 %) Desulfotomaculum 0,02 %) Desulfurispora (0,02 %) |
Наибольшее разнообразие СРБ выявлено в микробных матах. Многочисленными в микробном сообществе были представители рода Thermodesulfovibrio (класс Nitrospira). В микробных матах они составляли 36–40 %, в донных осадках их численность достигала 14 %. Следующими по численности были представители рода Desulfarculus (класс Deltaproteobacteria) (от 0,5 до 1,1 %), остальные СРБ были немногочисленны или встречались единично ( Desulfobacca, Desulfococcus, Desulfotomaculum, Thermodesulfobium, Desulfurispora ). Бактерии рода Thermodesulfobacterium (класс Thermodesulfobacteria) и Desulfurispora (класс Clostridia) были выявлены только в пробах образцов со станции с температурой при 58,1 °С.
Прогнозирование экологической функции микробов показывает, что многочисленные и редкие подсообщества по-разному реагируют на факторы окружающей среды, что позволяет предположить, что они могут занимать разные экологические ниши в горячих источниках. Редкие субсообщества могут играть более важную роль в деградации органического вещества, чем их многочисленные аналоги в горячих источниках [Jousset et al., 2017; Zhang et al., 2018].
Как правило, доминирующие таксоны демонстрируют сильную адаптацию к окружающей среде, но редкие таксоны становятся доминирующими при подходящих условиях окружающей среды. Также было обнаружено, что редкие таксоны обладают высокой активностью [Kurm et al., 2019]. Численность специализированных таксонов должна увеличиваться в ответ на конкретные изменения условий окружающей среды, которые приносят пользу им, а не другим таксонам.
Методом культивирования на элективных средах для выделения СРБ были показаны широкое распространение, высокая численность и морфологическое разнообразие мезофильных и термофильных СРБ в микробном сообществе щелочных гидротермах БРЗ.
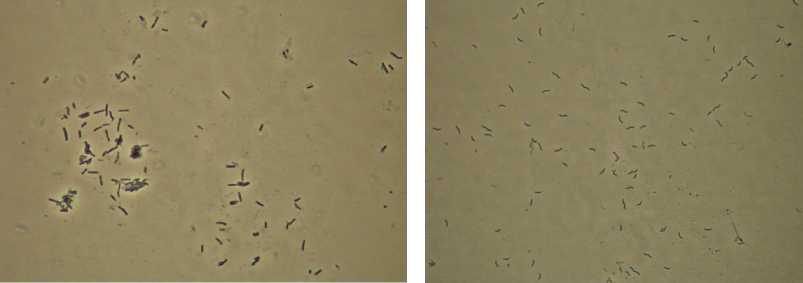
Накопительные культуры сульфатредуцирующих бактерий были выделены из микробных матов и донных осадков гидротерм Уро, Гарга, Горячинск, Умхэй, Сеюя и Гусиха (рис. 2, табл. 5).
а) б)
Рис. 2. Накопительные культуры Ga-1 (а) и Ur51L (б)
Таблица 5
Характеристика накопительных культур
|
Источник |
Культура (температура выделения, ° С) |
Тип пробы |
Морфотип клеток |
|
Уро |
Ur51L (30) |
д. ос. |
подвижные вибрионы, одиночные и сцепленные между собой |
|
Ur53L (30) |
д. ос. |
подвижные вибрионы, одиночные и сцепленные между собой |
|
|
Уро-10-ручей (30) |
д. ос. |
подвижные вибрионы, одиночные |
|
|
Уро-10-2 (55) |
д. ос. |
подвижные вибрионы, одиночные, иногда сцепленные по 2–4 клетки |
|
|
Уро-10-ручей (55) |
д. ос. |
подвижные вибрионы, одиночные, иногда сцепленные по 2–4 клетки |
Продолжение табл. 5
|
Источник |
Культура (температура выделения, ° С) |
Тип пробы |
Морфотип клеток |
|
Горячинск |
Гор-08-1-2 (30) |
м. м. |
подвижные вибрионы, одиночные |
|
Гор-10-1 (30) |
д. ос. |
неподвижные палочки, одиночные |
|
|
Гор-10-3 (30) |
д. ос. |
неподвижные палочки, одиночные |
|
|
Гор-10-1 (55) |
д. ос. |
неподвижные палочки , одиночные |
|
|
Гор-09 (55) |
д. ос. |
неподвижные палочки, одиночные и сцепленные по 2 клетки |
|
|
Гарга |
Га-07-2 (55) |
д. ос. |
тонкие прямые палочки, неподвижные, одиночные и сцепленные |
|
Га-10-3 (55) |
д. ос. |
тонкие слегка изогнутые палочки, одиночные, неподвижные |
|
|
Га-1 (55) |
д. ос. |
тонкие прямые палочки, неподвижные, одиночные и сцепленные |
|
|
Гусиха |
Гус-1-43 (55) |
м. м. |
тонкие изогнутые палочки, неподвижные, одиночные и сцепленные по 2 клетки |
|
Сеюя |
Se-08-1-2 (55) |
м. м. |
неподвижные палочки, одиночные |
|
Se-08-2 (55) |
д. ос. |
неподвижные палочки, одиночные |
Примечание: д. ос. — донный осадок, м. м. — микробный мат
В качестве доноров электронов и единственного источника углерода кроме лактата и ацетата СРБ могут использовать пируват, сукцинат, цистеин сахара и спирты (табл. 6).
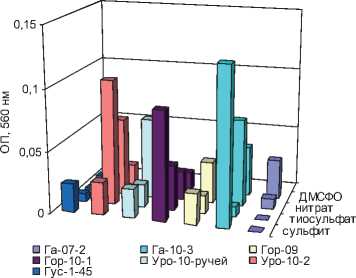
Большинство выделенных СРБ показало способность к росту при использовании сульфита, тиосульфата и нитрата в качестве единственного источника энергии и конечного акцептора электронов. Диметилсульфоксид могли использовать лишь некоторые СРБ (рис. 3).
0,12

0,08
0,04
ДМСФО нитрат тиосульфат сульфит
Ur51L Ur53L Уро-10-ручей Гор-10-1 Гор-10-3
Рис. 3. Рост СРБ на альтернативных акцепторах электронов: а) мезофильных; б) термофильных
Влияние различных субстратов на рост СРБ
Таблица 6
|
5 |
5 н CD я cd |
5 |
5 s H Ю |
5 К о s о Рч к |
Г) |
о к cd Н m |
cd s 2 |
cd О й |
cd О к к cd £ |
cd О к cd Рч |
ё s Рч о |
к s Рч о О |
к S CD Н О S д |
о
cd Н |
к S и cd Ц cd |
ё К S п к о |
ё S к |
|
|
Se-08-1-2 |
++ |
+ |
— |
++ |
— |
++ |
+ |
+ |
— |
++ |
+ |
— |
н. о. |
н. о. |
н. о. |
+ |
н. о. |
н. о. |
|
Se-08-2 |
++ |
++ |
+ |
— |
++ |
++ |
+ |
+ |
++ |
++ |
++ |
++ |
н. о. |
н. о. |
н. о. |
++ |
н. о. |
н. о. |
|
Ga-1 |
++ |
+ |
— |
++ |
— |
++ |
+ |
— |
— |
— |
+ |
— |
н. о. |
н. о. |
н. о. |
+ |
н. о. |
н. о. |
|
Um-09-1 |
++ |
— |
— |
— |
+ |
++ |
+ |
— |
+ |
— |
— |
— |
— |
н. о. |
— |
н. о. |
н. о. |
н. о. |
|
Um-09-2 |
+ |
— |
— |
— |
— |
— |
— |
+ |
— |
+ |
+ |
— |
— |
— |
— |
н. о. |
н. о. |
н. о. |
|
Um-09-3 |
++ |
+ |
— |
— |
— |
— |
+ |
— |
+ |
— |
— |
— |
— |
н. о. |
— |
н. о. |
н. о. |
н. о. |
|
Ur51L |
++ |
— |
— |
н. о. |
+ |
+ |
— |
++ |
— |
— |
— |
н. о. |
— |
++ |
+ |
н. о. |
+ |
+ |
|
Ur53L |
++ |
— |
+ |
н. о. |
+ |
— |
— |
— |
+ |
— |
н. о. |
— |
+ |
— |
н. о. |
+ |
++ |
|
|
Уро-10-ручей (30) |
++ |
+ |
+ |
н. о. |
+ |
— |
+ |
++ |
+ |
— |
н. о. |
++ |
++ |
— |
н. о. |
+ |
+ |
|
|
Гор-10-1 (30) |
++ |
+ |
+ |
н. о. |
+ |
+ |
+ |
— |
— |
— |
— |
н. о. |
++ |
++ |
— |
н. о. |
+ |
++ |
|
Гор-10-3 (30) |
++ |
+ |
— |
н. о. |
+ |
— |
+ |
— |
— |
— |
— |
н. о. |
— |
++ |
— |
н. о. |
н. о. |
н. о. |
|
Уро-10-2 (55) |
++ |
+ |
+ |
н. о. |
+ |
+ |
+ |
— |
++ |
+ |
+ |
н. о. |
++ |
+ |
— |
н. о. |
+ |
++ |
|
Уро-10-ручей (55) |
++ |
+ |
+ |
н. о. |
+ |
+ |
+ |
— |
+ |
+ |
+ |
н. о. |
— |
++ |
— |
н. о. |
+ |
+ |
|
Га-10-3 (55) |
++ |
+ |
+ |
н. о. |
+ |
+ |
— |
— |
+ |
— |
— |
н. о. |
++ |
++ |
— |
н. о. |
+ |
+ |
|
Гор-10-1 (55) |
++ |
+ |
+ |
н. о. |
+ |
++ |
+ |
+ |
++ |
— |
+ |
н. о. |
— |
++ |
— |
н. о. |
+ |
+ |
Примечание: «++» — активный рост, «+» — слабый рост, «–» — рост не обнаружен, «н. о.» — не определялось
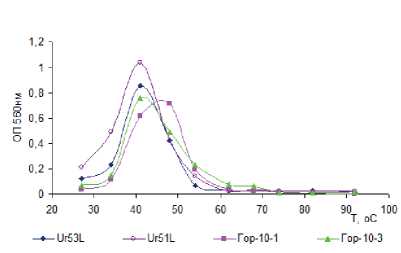
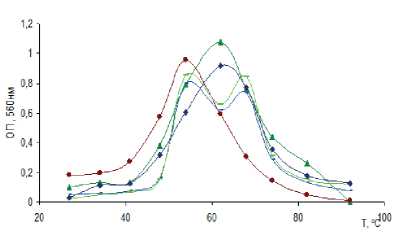
Все культуры СРБ, выделенные из щелочных гидротерм с различным температурным режимом, являются умеренными термофилами. Диапазон роста культур СРБ, выделенных при температуре 30 °С (рис. 4а), варьировал от 30 до 55 °С с оптимумом 40 °С, кроме Гор-10-1, у которого оптимум находился при температуре 45 °С. Диапазон роста культур СРБ, выделенных при температуре 55 °С, был шире — от 30 до 75 °С (рис. 4б) с оптимумом при 55 и 65 °С. Для бинарной культуры из микробного мата и осадка гидротермы Сеюя наблюдались два пика оптимума — первый при 55 °С , второй при 60 °С при диапазоне их развития 45–75 °С.
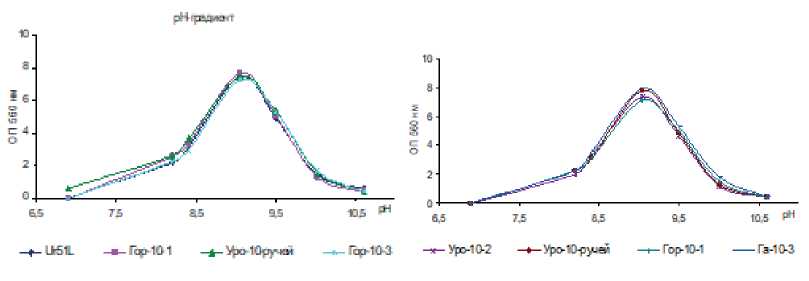
Культуры СРБ были способны развиваться в области рН от 7,0 до 10,5. Оптимальными значениями рН для роста исследуемых культур являются значения 9,0–9,2. По Вигелю [Wiegel, 1998], выделенные микроорганизмы относятся к алкалотолерантам.
Таким образом, культивируемое сульфидогенное сообщество слабоминерализованных щелочных гидротерм БРЗ представлено умеренно термофильными алка-лотолерантными СРБ.
а)
б)
в)
г)
Рис. 4. Рост накопительных культур СРБ в зависимости от температуры (а,б) и рН (в,г)
Заключение
В этой работе мы использовали комбинацию культуральных и молекулярных подходов для получения новой информации о разнообразии микроорганизмов, участвующих в биогеохимическом цикле серы в микробных матах и осадках вдоль температурного градиента в слабоминерализованных гидротермах Байкальской рифтовой зоны. Классическими микробиологическими и молекулярными методами показано, что термофильные и мезофильные алкалотолерантные сульфатре-дуцирующие бактерии широко распространены в щелочных условиях горячих источников БРЗ. Сульфидогенное сообщество щелочных гидротерм БРЗ участвует в восстановительной фазе цикла серы температуре от 30 до 75 ºС в диапазоне рН от 7 до 10,5.
Методом высокопроизводительного секвенирования гена 16S рРНК показано, что бактерии, осуществляющие диссимиляционное восстановление сульфатов, представлены сульфатредуцирующими бактериями классов Nitrospira, Deltaproteobacteria, Clostridia и Thermodesulfobacteria. В популяции сульфидогенного сообщества гидротерм многочисленными были термофильные бактерии рода Thermodesulfovibrio (класс Nitrospira) и мезофильные бактерии рода Desulfarculus (класс Deltaproteobacteria), редкими были бактерии рода Desulfobacca ,
Desulfococcus (класс Deltaproteobacteria), Desulfotomaculum, Thermodesulfobium, Desulfurispora (класс Clostridia).
Полученные данные свидетельствуют о том, что многочисленные и редкие таксоны СРБ могут занимать разные экологические ниши и вместе сохранять экологическую значимость в щелочных гидротермах БРЗ. Различие между редкими и доминирующими таксонами и их реакцией на изменения окружающей среды может иметь особое значение при попытке предсказать изменения в составе и функционировании сообщества.
Экстремальная щелочная среда горячих источников Байкальской рифтовой зоны имеет большой потенциал для изоляции сульфатредуцирующих бактерий.
Список литературы Распространение и разнообразие сульфатредуцирующих бактерий в слабоминерализованных щелочных гидротермах Байкальской рифтовой зоны
- Борисенко И. М., Замана Л. В. Минеральные воды Бурятской АССР. Улан-Удэ: Бурят. кн. изд-во. 1978. 162 с. Текст: непосредственный.
- Таксономический состав микробного сообщества горячего источника Алла (Байкальская рифтовая зона) / В. Г. Будагаева, А. А. Раднагуруева, А. В. Малыгин, Д. Д. Бар-хутова // Вестник Бурятского государственного университета. Биология. География. 2021. № 1. С. 3-11. DOI 10.18101/2587-7148-2021-1-3-11. Текст: непосредственный.
- Заварзин Г. А. Лекции по природоведческой микробиологии. Москва: Наука, 2003. 348 с. Текст: непосредственный.
- Изотопы сульфидной и сульфатной серы в азотных термах Баунтовской группы (Байкальская рифтовая зона) / Л. В. Замана, Ш. А. Аскаров, С. В. Борзенко [и др.] // Доклады Академии наук. 2010. Т. 435, № 3. С. 369-371. Текст: непосредственный.
- Кузнецов С. И. Дубинина Г. А. Методы изучения водных микроорганизмов. Москва: Наука, 1989. 288 с. Текст: непосредственный.
- Филогенетический анализ микробного мата в горячем источнике Гарга (Байкальская рифтовая зона) и разнообразие природных пептидаз / Е. В. Лаврентьева, А. А. Раднагуруева, Т. Г. Банзаракцаева [и др.] // Вавиловский журнал генетики и селекции. 2017. Т. 21(8). С. 959-963. DOI 10.18699/VJ17.319. Текст: непосредственный.
- Микробное сообщество термального озера Умхей (Байкальская рифтовая зона) в зоне разгрузки подземных вод / Е. В. Лаврентьева, Т. Г. Банзаракцаева, А. А. Раднагу-руева [и др.] // Сибирский экологический журнал. 2019. Т. 26, № 6. С. 715-726. Текст: непосредственный.
- Полевой практикум по водной микробиологии и гидрохимии / Б. Б. Намсараев, В. М. Горленко, З. Б. Намсараев [и др.] ; ответственный редактор М. Б. Вайнштейн. Улан-Удэ: Изд-во Бурят. гос. ун-та, 2005. 68 с. Текст: непосредственный.
- Микробные сообщества щелочных гидротерм / З. Б. Намсараев, В. М. Горленко, Намсараев Б. Б., Бархутова Д. Д. [и др.]. Новосибирск: Изд-во СО РАН, 2006. 111 с. Текст: непосредственный.
- Органотрофные бактерии горячих источников Байкальской рифтовой зоны / А. А. Раднагуруева, Е. В. Лаврентьева, В. Г. Будагаева [и др.] // Микробиология. 2016. Т. 85, № 3. С. 347-360. Текст: непосредственный.
- Aüllo T., Ranchou-Peyruse A., Ollivier B. and Magot M. Desulfotomaculum spp. and Related Gram-Positive Sulfate-Reducing Bacteria in Deep Subsurface Environments. Front. Microbiol. 2013; 4: 362. doi: 10.3389/fmicb.2013.00362
- Barkhutova D. D., Buryukhaev S. P., Dambaev V B. et al. Taxonomical and Functional Diversity of Microbial Communities in Two Hot Springs of the Baikal Rift
- Zone. IOP Conference Series: Earth and Environmental Science. 2021; 908: 012001. D01:10.1088/1755-1315/908/l/012001
- Dubovskiy I. M., Grizanova E. V, Whitten M. A. A. et al. Immuno-Physiological Adaptations Confer Wax Moth Galleria mellonella Resistance to Bacillus thuringiensis. Virulence. 2016; 7(8): 860-870. DOI: 10.1080/21505594.2016.1164367.
- Jousset A., Bienhold C., Chatzinotas A. et al. Where Less May Be More: How the Rare Biosphere Pulls Ecosystems Strings. ISME. 2017; 11: 853-862. doi: 10.1038/ismej.2016.174
- Kurm V, Geisen S., Hol W. H. G. A low Proportion of Rare Bacterial Taxa Responds to Abiotic Changes Compared with Dominant Taxa. Environ. Microb. 2019; 21: 750-758. 10.1111/1462-2920.14492
- Muyzer G., and Stams A. J. M. The Ecology and Biotechnology of Sulphate-Reducing Bacteria. Nature Reviews Microbiology. 2008; 6: 441-456. http://dx.doi.org/10.1038/ nrmicro1892
- Pedros-Alio, C. The Rare Bacterial Biosphere. Annu. Rev. Mar. Sci. 2012; 4: 449-466. doi: 10.1146/annurev-marine-120710-100948
- Rabus R., Venceslau S. S., Wohlbrand L. A Post-Genomic View of the Ecophysiology, Catabolism and Biotechnology Relevance of Sulfate-Reducing Prokaryotes. Advances in Microbial Physiology. 2015; 66: 55-321.
- Wiegel J. Anaerobic Alkalithermophiles, a Novel Group of Extremophiles. Extremophiles. 1998; 2: 257-267.
- Zhang Y., Wu G., Jiang H. et al. Abundant and Rare Microbial Biospheres Respond Differently to Environmental and Spatial Factors in Tibetan Hot Springs. Front. Microbiol. 2018; 9: 2096. doi: 10.3389/fmicb.2018.02096
