Распространение популяции промежуточной формы елей европейской и сибирской в российской части ареала

Автор: Попов П. П.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесная генетика и селекция
Статья в выпуске: 1, 2020 года.
Бесплатный доступ
В статье рассматривается фенотипическая структура популяций елей европейской и сибирской в восточно-европейской части их ареала. По преобладанию особей промежуточных фенотипов определено географическое положение популяций промежуточной формы елей европейской и сибирской на территории России, которые могут быть перспективными для лесоводства.
Ель европейская и сибирская, фенотипы, популяции, промежуточная форма
Короткий адрес: https://sciup.org/143170882
IDR: 143170882 | УДК: 630.165.1:630.165.5 | DOI: 10.24419/LHI.2304-3083.2020.1.07
Distribution of intermediate populations of the European and Siberian spruces in the Russian part of the area
The work is devoted to the study of the structure and distribution of intermediate populations of European and Siberian spruce in eastern Europe. Since a sufficiently objective reception of the definition of the region of intermediate populations of spruce has not yet occurred, the metric methods for evaluating the shape of seed scales were used to solve this problem, on the basis of which the corresponding phenotypes of individuals were selected. According to the predominance of individuals of intermediate phenotypes, intermediate populations and their geographic location were identified. In the Russian part of the range, on a wide strip of territory from Karelia to the central bed of the Kama (in the Perm Territory), there are three groups of populations of the intermediate form of European and Siberian spruces. They are characterized by the predominance of individuals of intermediate phenotypes (50% or more), intermediate parameters of many characters, but an increased indicator of the diversity of phenotypes and some genetic characters. The first of the groups of the intermediate form is geographically and in terms of the phenotypic structure of the populations closer to the European spruce, the third to the Siberian spruce, the second occupies the middle position. The results of the study may be useful for solving the issues of population-geographical diversity of spruce and its use in forestry.
Текст научной статьи Распространение популяции промежуточной формы елей европейской и сибирской в российской части ареала
Для ссылок: –3083.2020.1.07
Попов, П. П. Распространение популяции промежуточной формы елей европейской и сибирской в российской части ареала / П. П. Попов . – DOI 10.24419/LHI.2304-3083.2020.1.07. – Текст : электронный // Лесохоз. информ. : электронный сетевой журнал. – 2020. – № 1. – С. 69–75. URL:
В общем ареале елей европейской Picea abies (L.) Karst. и сибирской Picea obovata Ledeb. на востоке Европы под влиянием естественно-исторических и почвенно-климатических факторов по многим признакам образовалась большая группа популяций промежуточной формы. Эти популяции отличаются, прежде всего, наличием особей промежуточных фенотипов или форм [1, 2]. Предполагается, что основной причиной их образования является естественная гибридизация елей европейской и сибирской [3–5]. Ф. Федорович в свое время отметил: «Для объяснения существования переходных форм, я укажу весьма смелую, но не невозможную причину – перекрестное оплодотворение, на том основании, что северо-восточные и юго-западные ветры для Европы считаются господствующими» [3, с. 17]. Вероятно, он придавал большее значение ветрам, как фактору, способствующему переносу пыльцы ели. Промежуточные формы и их распространение в том или ином районе в большинстве случаев выявляют визуальным способом, определяя формы семенных чешуй, в частности их верхней части. В результате субъективной оценки признака часто получают несопоставимые результаты даже для одних и тех же районов [1–6]. Поэтому до сих пор нет четкой (объективной) картины распространения популяций промежуточной формы ели и их фенотипической структуры.
Цель настоящей работы – изучение фенотипической структуры и географического распространения популяций промежуточной формы елей европейской и сибирской на основе метрической оценки семенных чешуй. Район исследований — ареал ели от украинских Карпат до Урала. Дополнительно использовали шишки ели с территории Норвегии и Швеции, собранные С. П. Арефьевым и В. Р. Цибульским по методике автора [2, 7].
Объективной оценкой формы семенных чешуй может быть определение коэффициентов сужения (coefficient of narrowing – Cn) и вытянутости (coefficient of projection – Cp) их верхней части [7]. Более информативным показателем дифференциации особей и популяций елей европейской сибирской и их промежуточных форм является разность этих коэффициентов (Cn–Cp). На востоке Европы выделены 9 районов распространения популяций разных фенотипов (групп), различающихся в среднем на 10% показателя Cn–Cp [8]: P.e. – Picea europaea; P.eem. – Picea europaea-europaea-medioxima; P.em. – Picea europaea-medioxima; P.emm. – Picea europaea-medioxima-medioxima; P.m. – Picea medioxima; P.mms. – Picea medioxima-medioxima-sibirica; P.ms. – Picea medioxima-sibirica; P.mss. – Picea medioxima-sibirica-sibirica; P.s. – Picea sibirica.
Такую же градацию принимаем и для определения фенотипов особей в популяциях [9].
|
Фенотип особи |
e |
eem |
em |
emm |
m |
mms |
ms |
mss |
s |
|
Cn – Cp, % |
–50 |
–40 |
–30 |
–20 |
–10 |
0 |
10 |
20 |
30 |
Первый блок фенотипов ( e, eem, em ) представляет ель европейскую (evr), следующий блок ( emm, m, mms ) – промежуточную форму елей европейской и сибирской (med), последний блок ( ms, mss, s ) – ель сибирскую (sib). Такое разделение фенотипов соответствует сложившимся представлениям об интрогрессивной гибридизации елей европейской и сибирской, а также географической дифференциации популяций разных групп (фенотипов) в общем пространстве их ареала [8].
В пределах ареала елей европейской и сибирской наблюдается большая изменчивость структуры разных групп (фенотипов) популяций [8, 9] по частоте фенотипов особей (рис. 1). Группы популяций (1–9) существенно различаются и по блокам фенотипов особей (evr, med, sib). Кривые частот фенотипов пересекаются в точке, соответствующей частоте 50%. Популяции с частотой особей промежуточных фенотипов 50% и более представляют популяции промежуточной формы ели. Точка пересечения кривых, обозначающих блоки evr и med, соответствует положению между группами 3 и 4, пересечение кривых med и sib – между группами 6 и 7. Здесь и проходят юго-западная и северо-восточная границы популяций промежуточной формы елей европейской и сибирской (рис. 2). К западу и к востоку от нее в смежных районах располагаются популяции с частотой промежуточных фенотипов 30–35%, еще далее в тех же направлениях частота их резко снижается, а затем полностью исчезает. Популяции с частотой промежуточных фенотипов около 1/3 по существу являются переходными к елям европейской и сибирской.
Популяции промежуточной формы включают блок med из трех фенотипов особей ( emm , m , mms ), заметно различающихся по частоте встречаемости (рис. 3). Соотношение частот фенотипов, как видим, практически одинаковое с преобладанием (29—30%) в группах 4, 5, 6 и несколько меньшей частотой (22–24%) в смежных группах. В группах 1–3 и 7–9 она резко снижается – до 8–10 и 1–4% соответственно.
Структура популяций промежуточной формы ели в некоторых ключевых пунктах исследований в целом соответствует средним показателям по группам (таблица).
Показатель разнообразия по частоте фенотипов (при их 9-классной градации) составляет значительную величину и также близок к средним по группам (6–7) при минимальных значениях (около 2–3) в группах 1 и 9 [9].
Популяции промежуточной формы ели распространены на обширной полосе территории от западной части Республики Карелии к основному руслу Камы с достаточно благоприятными почвенно-климатическими условиями: почвы примерно севернее 60-й параллели – дерновые, южнее – дерново-подзолистые, изотерма января в среднем близка к –16 °C, июля – +12...15 °С. Еловые древостои характеризуются классами бонитета: IV – на севере, III – в центральной части, II – в южной части ареала [10–12]; средними параметрами генеративных органов: средняя длина шишек – 7–9 см [13], масса 1 000 семян – 4,5–5,5 г, [14], число семядолей – 7–7,5 шт. [15]. Относительная высота семенного потомства популяций равна 113–138 (групп 4 и 5) и 88–113% (группа 6) [16]. В группах 4, 5, 6 зафиксирован наибольший показатель разнообразия по частоте фенотипов [9], наибольшая частота аллеля Gdh0.75 и промежуточное значение частоты аллеля Gpi0.80
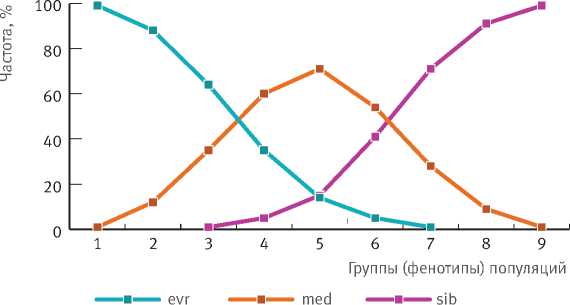
Рис. 1. Частота особей фенотипов ели европейской (evr), промежуточной формы (med) и ели сибирской (sib) по группам популяций (1–9) на востоке Европы и в Сибири
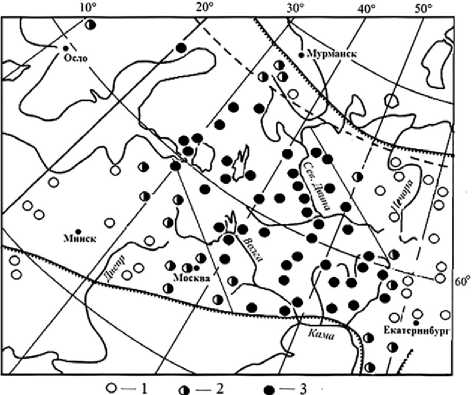
Рис. 2. Частота промежуточных фенотипов (1 – до 20%; 2 – 30–35%; 3 – 50% и более) в популяциях елей европейской и сибирской на востоке Европы
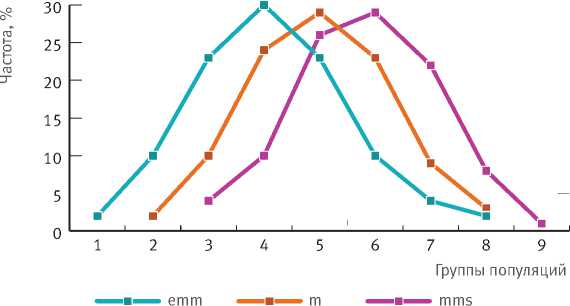
Рис. 3. Изменение частоты особей промежуточных фенотипов (emm, m, mms) по группам популяций (1–9) на востоке Европы и в Сибири.
Фенотипическая структура популяций промежуточной формы елей европейской и сибирской
[17]. Высокая частота промежуточных фенотипов позволяет предположить относительно большее их соответствие условиям произрастания [18] и повышенные потенциальные возможности расселения ели в районы с большим разнообразием условий произрастания.
Таким образом, можно отметить, что на востоке Европы в российской части ареала ели европейской и сибирской существует обширный район распространения популяций с преобладанием особей промежуточных фенотипов. Он располагается в направлении с северо-запада
(Республика Карелия) на юго-восток до основного русла Камы в Пермском крае. К юго-западу и к северо-востоку от него на узкой полосе территории размещаются переходные популяции с частотой промежуточных фенотипов около 30%. В составе популяций, произрастающих далее – к юго-западу и к северо-востоку, – доля особей промежуточных фенотипов мала, или они полностью отсутствуют. Результаты исследования могут быть полезны для решения вопросов популяционно-географического разнообразия ели и его использования в лесоводстве.
|
Населенный пункт сбора шишек |
Число особей, N, шт. |
Разность коэффициентов, Cn–Cp |
Частота фенотипов, %, по группам |
Блоки фенотипов, ∑ |
Разнообразие по частоте фенотипов, M |
||||||||||
|
e |
eem |
em |
emm |
m |
mms |
ms |
mss |
s |
1–3 |
4–6 |
7–9 |
||||
|
Группа 3 |
|||||||||||||||
|
Сортавала |
164 |
-17 |
4 |
4 |
23 |
27 |
23 |
13 |
5 |
1 |
- |
31 |
63 |
6 |
6.57 |
|
Ладва |
175 |
-18 |
1 |
10 |
21 |
26 |
29 |
7 |
4 |
2 |
- |
32 |
62 |
6 |
6.39 |
|
Иваново |
210 |
-18 |
4 |
8 |
17 |
29 |
24 |
10 |
7 |
1 |
- |
29 |
63 |
8 |
6.78 |
|
Группа 5 |
|||||||||||||||
|
Реболы |
252 |
-13 |
- |
6 |
13 |
30 |
27 |
14 |
9 |
1 |
- |
19 |
71 |
10 |
5.99 |
|
Коноша |
130 |
-10 |
- |
3 |
13 |
22 |
26 |
23 |
11 |
2 |
- |
16 |
71 |
13 |
6.08 |
|
Кологрив |
739 |
-13 |
1 |
3 |
14 |
27 |
28 |
15 |
10 |
2 |
- |
18 |
70 |
12 |
6.46 |
|
Группа 6 |
|||||||||||||||
|
Кемь |
160 |
-2 |
- |
1 |
8 |
16 |
19 |
24 |
17 |
12 |
3 |
9 |
59 |
32 |
6.97 |
|
Горка |
200 |
1 |
- |
1 |
2 |
11 |
14 |
31 |
25 |
13 |
3 |
3 |
56 |
41 |
6.44 |
|
Ижевск |
150 |
3 |
- |
1 |
3 |
5 |
19 |
27 |
25 |
15 |
5 |
4 |
51 |
45 |
6.57 |
Примечание. Географическое расположение пунктов: Сортавала, Ладва, Реболы, Кемь – в Республике Карелия; Иваново – в Ивановской обл.; Коноша, Горка – в Архангельской обл.; Кологрив – в Костромской обл.; Ижевск – в Республике Удмуртия.
Работа выполнена в рамках госзадания: проект № АААА-А17-117050400146-5 НИР Тюменского научного центра СО РАН.
Список литературы Распространение популяции промежуточной формы елей европейской и сибирской в российской части ареала
- Правдин, Л. Ф. Ель европейская и ель сибирская в СССР / Л. Ф. Правдин. - М.: Наука,1975. - 200 с.
- Попов, П. П. Распространение особей промежуточной формы в популяциях елей европейской и сибирской / П. П. Попов // Сиб. лесн. журн. - 2018. - № 4. - С. 13-19.
- Федорович, Ф. Новые наблюдения над сибирской елью (Picea obovata Ledeb.) / Ф. Федорович // Лесн. журн. - 1876. - Вып. 1. - С. 15-26.
- Данилов, Д. Н. Изменчивость семенных чешуй Picea excelsa / Д. Н. Данилов // Ботан. журн. - 1943. - Т. 28. - № 5. - С. 191-202.
- Бобров, Е. Г. Интрогрессивная гибридизация в роде Picea A. Dietr. / Е. Г. Бобров // Труды ин-та ЭРиЖ УНЦ АН СССР. - 1974. - Вып. 90. - С. 60-66.
- Орлова, Л. В. К систематике и географическому распространению ели финской (Picea fennica (Regel) Kom., Pinaceae) / Л. В. Орлова, А. А. Егоров // Новости систематики высших раст. - 2010. - Т. 42. - С. 5-23.
- Попов, П. П. Географическая изменчивость формы семенных чешуй ели в Восточной Европе и Западной Сибири / П. П. Попов // Лесоведение. - 1999. - № 1. - С. 68-73.
- Попов, П. П. Фенотипическая структура популяций Picea abies и P. obovata (Pinaceae) на востоке Европы / П. П. Попов // Ботан. журн. - 2013. - Т. 98. - № 11. - С. 1384-1402.
- Попов, П. П. Структура и разнообразие популяций елей европейской и сибирской по фенотипам / П. П. Попов // Экология. - 2017. - № 5. - С. 325-330.
- Цепляев, В. П. Леса СССР / В. П. Цепляев. - М.: Сельхозиздат, 1961. - 216 с.
- Атлас лесов СССР. - М.: ГУГК, 1973. - 222 с.
- Рысин, Л. П. Еловые леса России / Л. П. Рысин, Л. И. Савельева.- М.: Наука, 2002. - 335 с.
- Попов, П. П. Популяционно-географическая изменчивость шишек ели европейской и сибирской / П. П. Попов // Лесоведение. - 2011. - № 5. - С. 54-60.
- Попов, П. П. Популяционно-географическая изменчивость семян ели / П. П. Попов // Лесн. хоз-во. - 2006. - № 5. - С. 25-26.
- Попов, П. П. Популяционно-географическая изменчивость числа семядолей у всходов ели европейской и сибирской / П. П. Попов // Лесоведение. - 2013. - № 1. - С. 9-15.
- Попов, П. П. Изменчивость некоторых морфометрических показателей и фитомассы двухлетних сеянцев Picea abies и P. obovata (Pinaceae) разного географического происхождения / П. П. Попов // Растит. ресурсы. - 2007. - Т. 43. - № 4. - С. 32-41.
- Гончаренко, Г. Г. Популяционная и эволюционная генетика елей Палеарктики / Г. Г. Гончаренко, В. Е. Падутов. - Гомель: Институт леса НАНБ, 2001. - 197 с.
- Грант, В. Видообразование у растений / В. Грант. - М.: Мир, 1984. - 528 с.
