Распространение устойчивости к низким дозам ампициллина в популяции Escherichia coli

Автор: Ступак Евгения Эдуардовна, Ступак Игорь Валериевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биотехнология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
В статье рассматривается зависимость устойчивости клеток Escherichia coli к низким дозам ампициллина от концентрации антибиотика в среде и длительности его воздействия. Показано участие в развитии антибиотикоустойчивости как наследственных, так и ненаследственных факторов.
Антибиорезистентность, ампициллин, гетерогенность клональной популяции
Короткий адрес: https://sciup.org/148202073
IDR: 148202073 | УДК: 579.253
The spreading of resistances to low doses of ampicillin in the population Escherichia coli
In this paper we study the dependence of resistance of Escherichia coli cells to low doses of ampicillin on antibiotic concentrations in the environment and the duration of its action. The participation in the development of antibiotic resistance both of genetic and non-genetic factors is shown.
Текст научной статьи Распространение устойчивости к низким дозам ампициллина в популяции Escherichia coli
Современному человеку уже трудно представить свою жизнь без антибиотиков. Эти препараты спасают немало жизней, однако появляется все больше штаммов бактерий, устойчивых к их действию [1]. Поскольку многие микроорганизмы, например Escherichia coli , способны жить как в организме человека, так и во внешней среде, то, наряду с чрезмерным и бесконтрольным применением антибактериальных препаратов, одной из причин развития резистентности бактерий считается загрязнение антибиотиками окрестностей фармацевтических предприятий и животноводческих комплексов [2-3]. Повышенное содержание антибиотиков в отходах животноводства связано с применением антибиотиков в ветеринарии, в том числе в качестве пищевых добавок [4]. В загрязнении среды участвует и население, применяя антибиотики, используемые в медицине, для лечения животных и для обработки пораженных инфекциями декоративных и огородных растений.
В данной работе рассматривается распространение в популяции E.coli устойчивости к низким дозам ампициллина (Ap), полусинтетического антибиотика из группы пенициллинов, обладающего бактерицидным действием. Пенициллины, как и другие бета-лактамные антибиотики, препятствуют образованию пептидных связей между линейными молекулами пептидогликана, что приводит к потере прочности клеточной стенки и гибели бактерий в результате осмотического лизиса. Объединение пептидогликанов в единую структуру нарушается в результате инактивации пенициллинами пе-нициллин-связывающих белков, ферментов, принимающих участие в конечных стадиях формирования клеточной стенки бактерий [5].
Устойчивость к бета-лактамным антибиотикам связана с продукцией плазмидных и хромосомных бета-лактамаз, нарушением проницаемости наружной мембраны, модификацией пенициллин-связывающих белков [6, 7]. Существенную роль в формировании резистентности играет образование
биопленок [8] и способность бактериальных клеток к адгезии [9].
В работе был использован штамм E. coli HB101 (F-, thi-1, hsdS20(r B- , m B- ), supE44, recA13, ara-14, leuB6, proA2, lacY1, rpsL20, xyl-5, mtl-1, galK2 ). Культуру выращивали в жидкой или на агаризо-ванной среде LB [10]. Оптическую плотность культуры измеряли при длине волны 600 нм. Все эксперименты повторяли минимум в трех повторностях.
Эксперименты показали, что кривые роста при концентрации ампициллина 1 мкг/мл и в отсутствие антибиотика совпадают. Изменения в скорости роста штамма E. coli HB101 выявляются начиная с 2 мкг/мл Ap в среде. При посеве клеток, не контактировавших ранее с ампициллином, в жидкую среду, содержащую 2 или 3 мкг/мл Ap, первые 3-4 ч клетки делятся, затем рост останавливается и наблюдается гибель части клеток. Адаптация культуры к новым условиям происходит через 9 ч при концентрации 2 мкг/мл Ap и через 20 ч при 3 мкг/мл Ap (рис. 1). Культура, посеянная с чашки Петри, содержащей 2 мкг/мл Ap, адаптируется в жидкой среде к 3 мкг/мл за 13 ч, к 4 мкг/мл за 14 ч (рис. 2). Из графиков видно, что длительное время требуется для адаптации культуры к 2 и 3 мкг/мл Ap, в то время как разница во времени адаптации к 3 мкг/мл и 4 мкг/мл составляет всего час. Можно предположить, что механизмы обеспечивающие рост штамма E. coli HB101 при 3 мкг/мл Ap позволяют значительной части клеток выживать при 4 мкг/мл Ap.
Рассмотрим последствия воздействия ампициллина на изначально чувствительную к нему бактериальную популяцию. В отличие от других энтеробактерий, у E.coli отсутствует регуляторный ген ampR , поэтому экспрессия хромосомного гена ampC , кодирующего бета-лактомазу, не индуцируется бета-лактамами [11]. Ген ampC конститутивно экспрессируется на низком уровне, что связано со слабым промотором и наличием аттенюатора транскрипции в лидерной области.
1,8
1,6
1,4
1,2
0,8
0,6
0,4
0,2
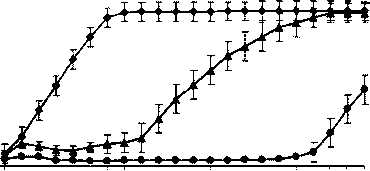
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
♦ Среда без Ap время, ч
Среда с Ap 2 мкг/мл
• Среда с Ap 3 мкг/мл
Рис. 1. Кривые роста периодической культуры E. coli НВ101 в среде без ампициллина (Ap), в среде, содержащей 2 мкг/мл или 3 мкг/мл ампициллина. Для посева использовались клетки, ранее культивировавшиеся в среде без ампициллина
1,8
1,6
1,4
1,2
0,8
0,6
0,4
0,2
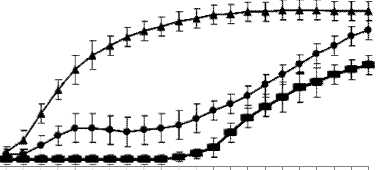
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
* Среда с Ap 2 мкг/мл время, ч
-
• Среда с Ap 3 мкг/мл
-
■ Среда с Ap 4 мкг/мл
Рис. 2. Кривые роста периодической культуры E. coli НВ101 в среде, содержащей 2, 3 или 4 мкг/мл ампициллина (Ap). Для посева были использованы клетки, ранее культивировавшиеся в среде, содержащей 2 мкг/мл ампициллина
Действие аттенюатора уменьшается с увеличением скорости роста культуры [12]. Образование бета-лактамазы в клетках E.coli может возрасти в результате мутаций, затрагивающих область промотора ampC или аттенюатора, интеграции последовательности IS2 в область промотора, а также при амплификации гена ampC [13]. Устойчивость к бета-лактамным антибиотикам, связанная с нарушением проницаемости внешней мембраны, - результат полной или частичной утраты поринов [14].
В популяции могут изначально присутствовать клетки нечувствительные к используемым дозам антибиотика. В обычных условиях такие клетки имеют меньшую скорость роста и составляют незначительную часть популяции. В новых условиях они получают селективное преимущество, а в дальнейшем, в результате накопления компенсационных мутаций, скорость роста популяции может быть восстановлена [15]. Воздействие антибиотиков индуцирует мутагенез, повышая содержание в бактериальных клетках активных форм кислорода, повреждающих ДНК [16]. В индуцированном антибиотиками мутагенезе принимает участие ДНК-полимераза IV, вызывающая в клетках, обработанных ампициллином, нуклеотидные замены. Наиболее часто встречаются AT->GC, GC->AT транзиции и AT -> CG трансверсии [17].
Динамика роста культуры в жидкой среде позволяет оценить способность популяции адаптироваться к новым условиям, но не дает представления об устойчивости к антибиотикам отдельных клеток. Чтобы исследовать фенотипический состав популяции в зависимости от длительности воздействия ампициллина и концентрации антибиотика в среде, был проведен эксперимент по схеме, представленной на рис. 3.

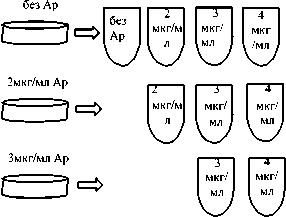
четыре
Из каждой пробирки рассевали клетки на ки:
без Ap
2 мкг/мл Ap
3 мкг/мл Ap
чаш-

4 мкг/мл Ap
Рис. 3. Схема пересева клеток. Периодическая культура росла в 3 мл среды при 37оС (160 об/мин). Чашки инкубировали 24 ч при 37оС
Эксперименты показали, что если популяция ранее не контактировала с ампициллином, то при рассеве на агаризованную среду с разными концентрациями ампициллина 3% клеток способны дать начало колониям на чашках, содержащих 2 мкг/мл Ap, и 0,002% - при 3 мкг/мл Ap (табл.). При посеве клеток в жидкую среду и последующем рассеве на чашки Петри было обнаружено, что в культуре, растущей в присутствии 2 мкг/мл Ap, 50% клеток не способны образовать колонии на агаризованной среде с той же концентрацией ампициллина и, соответственно, чувствительны к его воздействию. В популяции, растущей в жидкой культуре с 3 мкг/мл Ap или 4 мкг/мл Ap, без предварительной селекции рассевом, только 10% клеток способны образовать колонии при данной концентрации.
Таблица. Распространение устойчивости к ампициллину (Ap) в популяции E. coli.
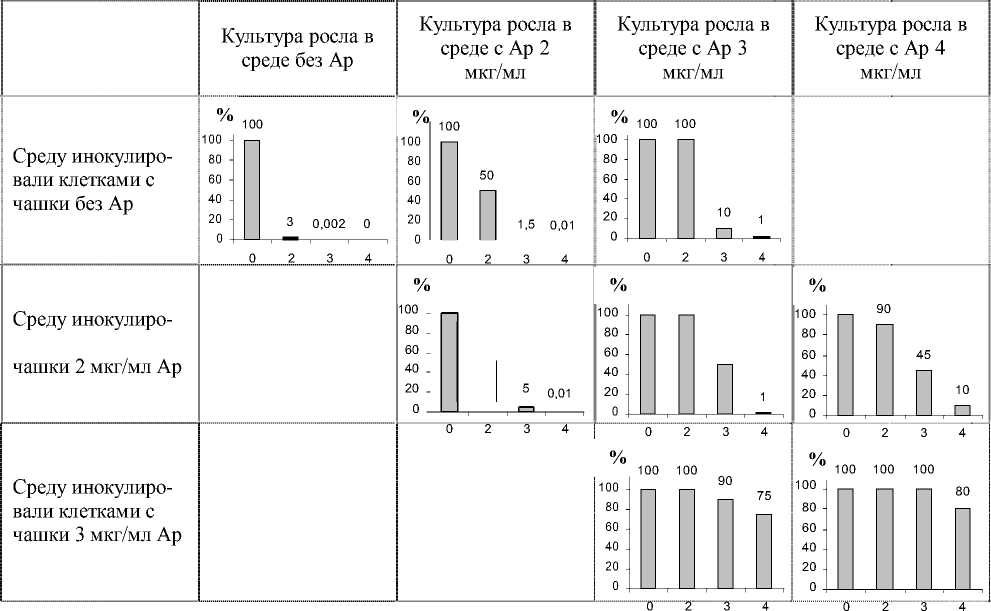
Прим. Жидкую среду с различной концентрацией ампициллина инокулировали клетками колоний, выросших на чашках со средой, содержащей от 0 до 3 мкг/мл ампициллина, затем культуру вновь рассевали на чашки Петри. Результаты рассева представлены на диаграммах. Номера столбцов диаграмм соответствуют содержанию ампициллина в агаризованной среде, высота столбцов – проценту клеток жидкой культуры, способных образовывать колонии на среде с указанной концентрацией ампициллина
С одной стороны, присутствие в популяции, успешно растущей в среде с ампициллином, чувствительных к ампициллину клеток, вероятно, связано с секрецией устойчивыми клетками бета-лактомазы. В результате концентрация антибиотика в среде становится несколько ниже первоначальной, и появляется возможность роста клеток, не имеющих генетически закрепленных механизмов устойчивости к данной концентрации ампициллина. С другой стороны, не все неустойчивые клетки могут расти в данных условиях. Показано, что клетки, не наследующие устойчивость к ампициллину, но способные в определенных условиях расти в его присутствии, как правило, отличаются от остальных паттерном экспрессии генов: чаще всего изменен уровень экспрессии генов бета-лактомазы, глутомат декарбоксилазы и dam-метилазы [18]. Метаболическая и фенотипическая гетерогенность бактериальной клональной популяции является результатом неоднородности клеток по концентрации каждого конкретного белка в результате стохастических процессов в экспрессии генов и распределении белков при делении клетки [19]. При стрессовых воздействиях, в силу тех же стохастических процессов, скорость изменения концентраций регуляторных белков и сигма-факторов РНК-полимеразы и, соответственно, паттернов экспрессии генов в подконтрольных им генетических сетях существенно колеблется от клетки к клетке, что увеличи- вает неоднородность популяции даже в отсутствие мутаций.
Увеличение времени культивирования клеток в присутствии 2 мкг/мл Ap, в том числе и с селективным отбором на чашках, не приводит к полному вытеснению из популяции неустойчивых к ампициллину клеток (табл.).
Возврат к неустойчивому фенотипу при рассеве до единичных колоний подтверждает, что часть клеток, образующих колонии на чашках, также не имеет генетически детерминированной устойчивости.
Устойчивость к 3 мкг/мл Ap растет при увеличении длительности воздействия и достигает 100% в культуре, прошедшей несколько повторных селекций рассевом до единичных колоний.
Исходя из полученных данных, можно предположить, что клетки, растущие в популяции на границе своей индивидуальной ингибирующей концентрации ампициллина, являются плацдармом для развития следующей ступени антибиотикоустойчи-вости.
Проведенные эксперименты показывают, что распространение устойчивости в популяции E.coli может быть обусловлено не только генетическими, но и ненаследственными адаптационными механизмами.
-
1. Bush K., Courvalin P., Dantas G. et al. Tackling antibiotic resistance // Nature reviews Microbiology. 2011. V. 9. P. 894-895.
-
2. Kim K.R., Owens G., Kwon S.I. et al. Occurrence and environmental fate of veterinary antibiotics in the terrestrial environment // Water, Air and Soil Pollution. 2011. V. 214. P. 163-174.
-
3. Tello A., Austin B., Telfer T.C. Selective pressure of antibiotic pollution on bacteria of importance to public health // Environ Health Perspect. 2012. V. 120. P. 1100–1106.
-
4. Sarmah A.K., Meyer M.T., Boxall A.B.A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment // Chemosphere. 2006. V. 65. P. 725-759.
-
5. Kong K.F., Schneper L., Mathee K. Beta-lactam antibiotics: from antibiosis to resistance and bacteriology // APMIS. 2010. V. 118. P. 1-36.
-
6. Georgopapadakou N.H. Penicillin-binding proteins and bacterial resistance to beta-lactams // Antimicrob Agents Che-mother. 1993. V. 37. P. 2045-2053.
-
7. Семёнов В.М., Дмитраченко Т.И., Жильцов И.В. Микробиологические и биологические аспекты резистентности к антимикробным препаратам // Медицинские новости. 2004. № 2. С. 10-17.
-
8. Ito A., Taniuchi A., May T. et al. Increased antibiotic resistance of Escherichia coli in mature biofilms // Appl. Environ. Microbiol. 2009. V. 75. P. 4093-4100.
-
9. Hart A., Nowicki B.J., Reisner B. et al. Ampicillin-resistant Escherichia coli in gestational pyelonephritis: increased occurrence and association with the colonization factor Dr adhesion // J. Infect. Diseases. 2001. V. 183. P. 1526-1529.
-
10. Миллер Дж. Эксперименты в молекулярной генетике. М.: Мир, 1976. 436 с.
-
11. Honoré N., Nicolas M.H., Cole S.T. Inducible cephalospo-rinase production in clinical isolates of Enterobacter cloacae is controlled by a regulatory gene that has been deleted from Escherichia coli // EMBO J. 1986. V. 5. P. 3709-3714.
-
12. Jaurin B., Grundstrom T., Edlund T., Normark S. The E. coli beta-lactamase attenuator mediates growth rate-dependent regulation // Nature (London). 1981. V. 290. P. 221-225.
-
13. Nelson E.C., Elisha B.G. Molecular basis of AmpC hyperproduction in clinical isolates of Escherichia coli // Antimicrob Agents Chemother. 1999. V. 43. P. 957-959.
-
14. Harder K.J., Nikaido H., Matsuhashi M. Mutants of Escherichia coli that are resistant to certain β-lactam compounds lack the OmpF porin // Antimicrob Agents Chemother. 1981. V. 20. P. 549-552.
-
15. Andersson D.I., Hughes D. Antibiotic resistance and its cost: is it possible to reverse resistance? // Nat. Rev. Microbiol. 2010. V. 8. P. 260-271.
-
16. Kohanski M.A., DePristo M.A., Collins J.J. Sub-lethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis // Mol. Cell. 2010. V. 37. P. 311-320.
-
17. Gutierrez A., Laureti L., Crussard S. et al. B-lactam antibiotics promote mutagenesis via RpoS-mediated replication fidelity reduction // Nat. Commun. 2013. V. 8. URL: http://www.nature.com/ncomms/journal/v4/n3/full/ncomm s 2607.html
-
18. Adam M., Murali B., Glenn N.O., Potter S.S. Epigenetic inheritance based evolution of antibiotic resistance in bacteria // BMC Evol. Biol. 2008. V. 8. URL: http://www.biomedcentral.com/1471-2148/8/52/
-
19. Ступак Е.Э., Ступак И.В. Переключение фенотипов клеток Escherichia coli , содержащих циклические диген-ные системы с отрицательными обратными связями, при изменении условий культивирования // Генетика. 2010. Т. 46. № 5. С. 610-615.
THE SPREADING OF RESISTANCES TO LOW DOSES OF AMPICILLIN IN THE POPULATION ESCHERICHIA COLI
Institute of Biology, Ufa Sci. Center of RAS, Ufa
In this paper we study the dependence of resistance of Escherichia coli cells to low doses of ampicillin on antibiotic concentrations in the environment and the duration of its action. The participation in the development of antibiotic resistance both of genetic and non-genetic factors is shown.
Список литературы Распространение устойчивости к низким дозам ампициллина в популяции Escherichia coli
- Bush K., Courvalin P., Dantas G. et al. Tackling antibiotic resistance//Nature reviews Microbiology. 2011. V. 9. P. 894-895.
- Kim K.R., Owens G., Kwon S.I. et al. Occurrence and environmental fate of veterinary antibiotics in the terrestrial environment//Water, Air and Soil Pollution. 2011. V. 214. P. 163-174.
- Tello A., Austin B., Telfer T.C. Selective pressure of antibiotic pollution on bacteria of importance to public health//Environ Health Perspect. 2012. V. 120. P. 1100-1106.
- Sarmah A.K., Meyer M.T., Boxall A.B.A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment//Chemosphere. 2006. V. 65. P. 725-759.
- Kong K.F., Schneper L., Mathee K. Beta-lactam antibiotics: from antibiosis to resistance and bacteriology//APMIS. 2010. V. 118. P. 1-36.
- Georgopapadakou N.H. Penicillin-binding proteins and bacterial resistance to beta-lactams//Antimicrob Agents Chemother. 1993. V. 37. P. 2045-2053.
- Семёнов В.М., Дмитраченко Т.И., Жильцов И.В. Микробиологические и биологические аспекты резистентности к антимикробным препаратам//Медицинские новости. 2004. № 2. С. 10-17.
- Ito A., Taniuchi A., May T. et al. Increased antibiotic resistance of Escherichia coli in mature biofilms//Appl. Environ. Microbiol. 2009. V. 75. P. 4093-4100.
- Hart A., Nowicki B.J., Reisner B. et al. Ampicillin-resistant Escherichia coli in gestational pyelonephritis: increased occurrence and association with the colonization factor Dr adhesion//J. Infect. Diseases. 2001. V. 183. P. 1526-1529.
- Миллер Дж. Эксперименты в молекулярной генетике. М.: Мир, 1976. 436 с.
- Honoré N., Nicolas M.H., Cole S.T. Inducible cephalosporinase production in clinical isolates of Enterobacter cloacae is controlled by a regulatory gene that has been deleted from Escherichia coli//EMBO J. 1986. V. 5. P. 3709-3714.
- Jaurin B., Grundstrom T., Edlund T., Normark S. The E. coli beta-lactamase attenuator mediates growth rate-dependent regulation//Nature (London). 1981. V. 290. P. 221-225.
- Nelson E.C., Elisha B.G. Molecular basis of AmpC hyperproduction in clinical isolates of Escherichia coli//Antimicrob Agents Chemother. 1999. V. 43. P. 957-959.
- Harder K.J., Nikaido H., Matsuhashi M. Mutants of Escherichia coli that are resistant to certain β-lactam compounds lack the OmpF porin//Antimicrob Agents Chemother. 1981. V. 20. P. 549-552.
- Andersson D.I., Hughes D. Antibiotic resistance and its cost: is it possible to reverse resistance?//Nat. Rev. Microbiol. 2010. V. 8. P. 260-271.
- Kohanski M.A., DePristo M.A., Collins J.J. Sub-lethal antibiotic treatment leads to multidrug resistance via radicalinduced mutagenesis//Mol. Cell. 2010. V. 37. P. 311-320.
- Gutierrez A., Laureti L., Crussard S. et al. B-lactam antibiotics promote mutagenesis via RpoS-mediated replication fidelity reduction//Nat. Commun. 2013. V. 8. URL: http://www.nature.com/ncomms/journal/v4/n3/full/ncomms 2607.html
- Adam M., Murali B., Glenn N.O., Potter S.S. Epigenetic inheritance based evolution of antibiotic resistance in bacteria//BMC Evol. Biol. 2008. V. 8. URL: http://www.biomedcentral.com/1471-2148/8/52/
- Ступак Е.Э., Ступак И.В. Переключение фенотипов клеток Escherichia coli, содержащих циклические дигенные системы с отрицательными обратными связями, при изменении условий культивирования//Генетика. 2010. Т. 46. № 5. С. 610-615.
