Растительность долинных комплексов Среднего Приамурья (Хинганский заповедник)
")
Автор: Рец Е.П.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 3, 2012 года.
Бесплатный доступ
Выявлены закономерности дифференциации ботанического разнообразия на локальном уровне и факторы, его определяющие.
Среднее приамурье, долинный комплекс, растительность, α-разнообразие, ценотический анализ флоры
Короткий адрес: https://sciup.org/14082253
IDR: 14082253 | УДК: 581.93
The Central Priamurye valley complex vegetation (Khingansk national reserve)
The laws of the botanical variety differentiation at the local level and the factors, which determine it, are revealed.
Текст научной статьи Растительность долинных комплексов Среднего Приамурья (Хинганский заповедник)
Введение. Исследованию различных аспектов растительного покрова долинных комплексов посвящены многочисленные публикации. Охарактеризованы различные типы пойменных болот, долинных лугов, лесов [3, 4, 6, 7, 17, 18], разработаны классификации долинной растительности, в том числе по методу Браун-Бланке [1, 2, 5, 8, 13, 21], рассмотрено влияние различных экологических факторов на растительные ком- плексы долин [9]. Показано, что растительный покров речных долин формируется в динамичных условиях и отличается большой пестротой, сложностью и крайней неустойчивостью, как в пространстве, так и во времени.
Объект настоящего исследования – растительный покров долины Среднего Амура. Целью исследования является выявление закономерностей дифференциации ботанического разнообразия на локальном уровне и факторов его определяющих. В качестве модельной нами выбрана территория Хинганского заповедника (Архаринская низменность, юго-восточная часть Зейско-Буреинской равнины) [15, 16], никогда не испытавшая значительного антропогенного влияния, а с 1960 года полностью исключенная из любого вида хозяйственного использования.
Материалы и методы. Работа основана на материалах (766 описаний растительных сообществ), собранных автором в ходе 4 вегетационных сезонов (2006–2009 гг.). Описания выполнены в соответсвии с фитоценологическими стандартами, разработанными Muller-Dombois & Ellenberg [26]. Помимо этого используются результаты флористических [5,11], геоботанических [10, 20] и геоморфологических [12, 14, 15] исследований, а также космические снимки (Aster, Landsat и др.).
В качестве опорных единиц изучения таксономического разнообразия нами используется α-разнообразие (таксономическое и типологическое). В работе применяется стандартный набор методов (классификация по методу Браун-Бланке, эколого-флористический анализ ценофлор, ординация по методу шкал Раменского), неоднократно доказавших свою эффективность и поэтому часто используемых в современных исследованиях.
Обсуждение результатов. В результате проведенных исследований установлено, что флора изучаемых долинных комплексов включает в себя 410 видов растений, относящихся к 91 семейству. Из 10 самых богатых видами семейств преобладающими являются Compositae, Poaceae, Cyperaceae и Ranunculaceae , их суммарная доля превышает 50%. Классификация растительности выполнена по методу Braun-Blanquet с использованием пакета программ Turboveg и Juice . Широкий набор местообитаний и изменяющиеся по градиентам факторы среды обусловили существование большого числа ассоциаций (12), относящихся к 5 классам и 7 порядкам [1, 2, 8, 23].
Продромус растительности долинных комплексов Хинганского заповедника:
-
1 .0.0.0. Класс Phragmito-Magnocaricetea Klika in Klika et Novak 1941.
-
1.1. 0.0. Порядок Phragmitetalia W. Koch 1926.
-
1.1.1. 0. Союз Phragmition W. Koch 1926.
-
1.1.1.1. Асс. Glycerio spiculosae-Phragmitetum australis Akht. 1987.
-
-
-
1.2. 0.0.Порядок Magnocaricetalia Pignatti 1953
-
1.2.1. 0.Союз Caricion appendiculatae Akhtyamov et al. 1985.
-
1.2.1.1. Асс. Caricetum vesicato- appendiculatae Akhtyamov et al. 1985.
-
-
-
-
2 .0.0.0. Класс Scheuchzerio-Caricetea fuscae R.Tx. 1937.
-
2.1. 0.0. Порядок Caricetalia meyerianae Akhtyamov 1987.
-
2.1.1. 0. Союз Caricion meyerianae Akht. et al. 1987.
-
2.1.1.1. Асс. Menyantho trifoliatae-Caricetum meyerianae ass. nov.
-
2.1.1.2. Асс. Caricetum lasiocarpo-meyerianae Akht. 1987.
-
-
-
-
3 .0.0.0. Класс Calamagrostietea langsdorffii Mirkin in Akhtyamov et al. 1985.
-
3.1. 0.0. Порядок Calamagrostietalia langsdorffii Akhtyamov et al. 1985.
-
3.1.1. 0. Союз Calamagrostion langsdorffii Akhtyamov et al. 1985.
-
3.1.1.1. Асс. Lathyro pilosi-Calamagrostietum purpureae ass. nov.
-
-
3.1.2. 0. Cоюз Caricion schmidtii Akht. et al. 1985.
-
3.1.2.1. Асс. Calthii-Caricetum schmidtii Akhtymov 1987.
-
-
-
-
4 .0.0.0. Класс Arundinello anomalae-Agrostietea trinii Ermakov et Krestov 2009.
-
4.1. 0.0. Порядок Carici schmidtii-Agrostietalia trinii Ermakov et Krestov 2009.
-
4.1.1. 0. Союз Agrostion trinii Akhtyamov et al. 1985.
-
4.1.1.1. Асс. Carici schmidtii - Hemerocallidetum minoris ass. nov.
-
-
-
4.2. 0.0. Порядок Artemisietalia mandshuricae Akhtyamov et al. 1985.
-
4.2.1. 0. Союз Arundinellion anomalae Akht. et al. 1985
-
4.2.1.1. Асс. Cariceto schmidtii - Arundinelletum anomalae Akhtyamov et al. 1985.
-
-
-
-
5 .0.0.0. Класс Querco mongolicae-Betuletea davuricae Ermakov et Petelin 1997.
-
5.1. 0.0. Порядок Querco mongolicae-Betuletalia davuricae Ermakov 1997.
-
5.1.1. 0. Союз Ligulario fischeri - Betulion davuricae Ermakov 1997.
-
5.1.1.1. Асс. Carici schmidtii - Betuletum platyphyllae ass. nov.
-
5.1.1.2. Асс. Rubo saxatilis - Betuletum platyphyllae ass. nov.
-
-
5.1.2. 0. Союз Kitagawio terebinthaceae-Betulion davuricae Ermakov 1997.
-
5.1.2.1. Асс. Corylo heterophylae - Quercetum mongolicae Stupnikova 2002.
-
5.1.2.2. Асс. Artemisio gmelinii - Quercetum mongolicae ass. nov.
-
-
-
Заросли гигрофитов ( Zizania latifolia, Thypha latifolia и др.) и крупноосочники класса PHRAGMITI-MAGNOCARICETEA Klika in Klika et Novak 1941 произрастают на самых низких уровнях высокой поймы – глубоких мочажинах прарусловых понижений и околостаричных экотопах. Мезотрофные мейроосоковые ( C. meyeriana ) болота, входящие в состав класса SCHEUCHZERIO-CARICETEA FUSCAE R.Tx. 1937, занимают основные части плоских прарусловых понижений на высокой пойме. Класс CALAMAGROSTETEA LANGSDORFII Mirkin in Achtiamov et al. 1985 на территории исследования представлен сырыми и заболоченными вейниковыми (Calamagrostis purpurea) и шмидтосоково (Carex schmidtii) – вейниковыми лугами, которые формируются на нижних частях склонов, а также в неглубоких понижениях террас. Свежие и сухие разнотравные «лилейные» и разнотравно-злаковые ( Koeleria cristata, Arundinella anomale, Spodiopogon sibiricus и др.) луга класса ARUNDINELLO ANOMALAE- AGROSTIETEA TRINII cl. nov. hoc loco [9] занимают основные пространства плоских террасовидных поверхностей и определяют физиономический облик всей долины. На относительно автоморфных частях долины, вышедших из-под непосредственного влияния аллювиального фактора, последовательно сменяют друг друга различные типы мелколиственных ( Betula plathyphylla, Populus tremula ) и смешанных ( Quercus mongolica, Ulmus laciniata, Tilia amurensis, Betula davurica ) лесов класса QUERCO MONGOLICAE-BETULETEA DAVURICAE Ermakov et Petelin 1997.
Оценка варьирования различных показателей биоразнообразия по градиентам среды невозможна без проведения экологической ординации синтаксонов. Для экологической характеристики синтаксонов в пространстве ведущих факторов среды используются экологические шкалы Л.Г. Раменского [18, 19], а также европейские шкалы Г. Элленберга [24] и Э. Ландольта [25]. Пространственная координация синтаксонов проводилась в условном экологическом поле по двум факторам (увлажнение и богатство почв). Показатели по каждому фактору для видов растений брались из региональных таблиц, составленных И.А. Цаценкиным и др. [22] и В.П. Селедцом [19] для Дальнего Востока РФ.
Все синтаксоны долины располагаются в условном экологическое поле, в котором условия увлажнения (ось X) меняются от увлажнения сильнообводненных болот (105) до лугово-степного или влажностепного (48), а условия богатства почв (ось Y) от бедных почв (4) до богатых бурых, луговых, достаточно обеспеченных элементами минерального питания почв (16). В результате проведенного анализа получено 3 экологических ряда ассоциаций: болотный ряд – от ассоциации околоводных зарослей ( Glycerio spiculosae– Phragmitetum australis Akht. 1987) до мейроосоковых болот ( Caricetum lasiocarpo-meyerianae Akht. 1987) на высокой пойме, ряд луговых ассоциаций – от сыролуговых шмидтосоково-вейниковых лугов ( Lathyro pilosi-Calamagrostietum purpureae ass. nov.) до остепненных разнотравно-злаковых лугов ( Cariceto schmidtii-Arundinelletum anomalae Akhtyamov et al. 1985) на относительно выположенных поверхностях надпойменных террас и ряд лесных ассоциаций – от белоберезовых олуговелых ( Carici schmidtii - Betuletum platyphyllae ass. nov.) до остепненных дубрав ( Artemisio gmelinii - Quercetum mongolicae ass. nov. ) на древних прирусловых гривах на террасах.
Видовое богатство (ВБ) ассоциаций растительности долинных комплексов меняется в диапазоне 20 до 169 (рис.). Анализ изменений видового богатства ассоциаций в поле ведущих факторов среды показал, что оно связано с изменением условий увлажнения и богатства почв, достигая наибольших значений для растительных ассоциаций грив и террас (луговой и лесной ряды), а минимальные – для растительных ассоциаций обводненных прарусловых понижений на пойме (болотный ряд). Если на прарусловых понижениях поймы могут произрастать лишь 98 видов растений, постепенно по профилю долины экологическая емкость среды увеличивается: общее богатство луговых сообществ составляет 221, а лесных сообществ на гривах – 258 видов.
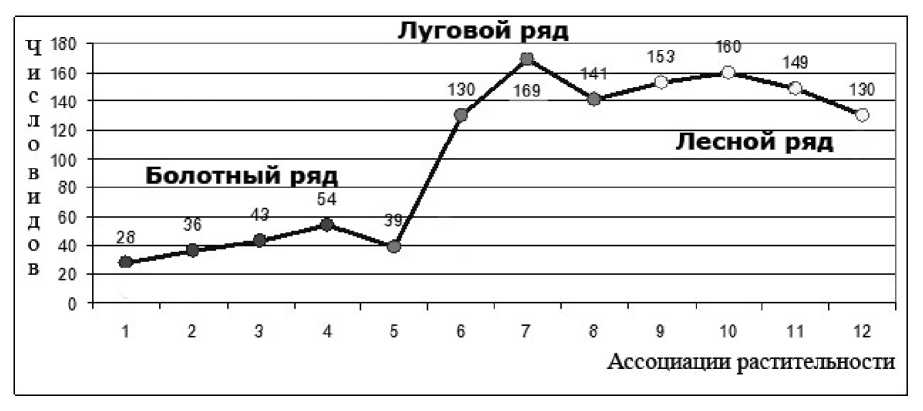
График изменения видового богатства: (1 – Glycerio spiculosae-Phragmitetum australis Akht. 1987;
-
2 – Caricetum vesicato- appendiculatae Akhtyamov et al. 1985; 3 – Menyantho trifoliatae-Caricetum meyerianae ass. nov.; 4 – Caricetum lasiocarpo-meyerianae Akht. 1987; 5 – Lathyro pilosi-Calamagrostietum purpureae ass. nov.; 6 – Calthii-Caricetum schmidtii Akhtymov 1987; 7 – Carici schmidti – Hemerocallidetum minoris ass. nov.; 8 – Cariceto schmidtii-Arundinelletum anomalae Akhtyamov et al. 1985; 9 – Carici schmidtii-Betuletum platyphyllae ass. nov.; 10 – Rubo saxatilis-Betuletum platyphyllae ass. nov.; 11 – Corylo heterophylae-Quercetum mongolicae Stupnikova 2002; 12 – Artemisio gmelinii-Quercetum mongolicae ass. nov.
Болотный ряд. Минимальные показатели видового богатства соответствуют ассоциации прибрежноводных сообществ (Glycerio spiculosae-Phragmitetum australis Akht. 1987). Такой низкий показатель связан с крайними экологическими условиями экотопа по фактору увлажнения, в которых может произрастать лишь ограниченное число видов. В связи с уменьшением увлажнения от ассоциации околоводных зарослей сообществ к ассоциации мезотрофных болот ( Caricetum lasiocarpo-meyerianae Akht. 1987) происходит постепенный рост ВБ. Наибольшие показатели ВБ (54) в пределах болотного ряда соответствуют ассоциации мейроосоковых кровохлебковых болот ( Caricetum lasiocarpo-meyerianae Akht. 1987). Здесь высокая мозаичность (кочки и микровалы), а также относительно благоприятные условия увлажнения приводят к росту видового богатства.
Луговой ряд. В ряду луговых ассоциаций на террасах происходит быстрый рост ВБ по градиенту увлажнения и богатства почв: от 39 в сырых шмидтосоково-вейниковых лугах до 169 в свежих «лилейных» лугах ( Carici schmidtii - Hemerocallidetum minoris ass. nov.). На первой ступени ряда увлажнение еще избыточно для луговых, но уже недостаточно для болотных видов. В связи с этим здесь формируется специфическая ассоциация монодоминантных сообществ Lathyro pilosi-Calamagrostietum purpureae ass. nov с низким видовым богатством (36). На следующей ступени ряда (влажные луга Calthii-Caricetum schmidtii Akhtymov 1987) происходит резкое увеличение видового богатства, связанное с улучшением экологической среды. Максимальные показатели видового богатства в связи с относительно благоприятными условиями увлажнения и богатства почв, как для этого ряда, так и среди всех ассоциаций долинных комплексов, имеет ассоциация свежих лугов ( Carici schmidtii - Hemerocallidetum minoris ass. nov.). В составе флоры ассоциации следующей ступени экологического ряда ( Cariceto schmidtii-Arundinelletum anomalae Akhtyamov et al. 1985), сообщества которой формируются на плоских поверхностях наиболее высоких и древних террас, в условиях сухолугового увлажнения выпадает большинство мезогигрофитов и гигромезофитов, появляются немногочисленные ксерофиты, что сказывается на показателе ВБ: он падает до 141.
Лесной ряд. Локальный максимум (160 видов) внутри ряда наблюдается в ассоциации мелколиственно-широколиственных лесов ( Rubo saxatilis - Betuletum platyphyllae ass. nov.), во флоре которой смешиваются комплексы видов субклимаксовых дубрав и луговолесных видов березняков и осинников. Локальный минимум соответствует остепненным дубравам на крутых южных склонах грив ( Artemisio gmelinii - Quercetum mongolicae ass. nov.). В условиях эдафической сухости из состава фитоценозов выпадает большое число мезофитных видов и таким образом снижается видовое богатство ассоциации (130).
Анализ важнейших показателей разнообразия по градиенту увлажнения и богатства почв позволил установить закономерности распределения отдельных типологических категорий, выявить связи различных элементов с определенными генетическими частями долины. Согласно полученным результатам, ядро флоры околоводных и болотных сообществ прарусловых понижений на пойме составляют широкоареальные лугово-болотные и водно-болотные виды, также довольно значимыми оказываются маньчжурские водно-болотные и лугово-болотные виды. В структуре влажных и сырых лугов на нижних уровнях террас доминируют маньчжурские гигрофильно-луговые виды. Маньчжурские луговые виды находят свой оптимум в различных типах лугов (от влажных до сухих) на плоских поверхностях террас. Виды, связанные с маньчжурскими неморальными экосистемами, начинают играть значимую роль в лесных ассоциациях на гривах террас, а максимальных показателей они достигают в субклимаксовых черноберезово-дубовых лесах на гривах 2 НТ. Дауро-маньчжурский степисто-лесной элемент, объединяющий виды, ценотически и в своем распространении связанные с континентальными дубравами Даурии и Среднего Амура, насыщает флоры субклимаксовых смешенных березово-дубовых лесов и остепненных дубрав на гривах. Южносибирские степистолуговые виды проникают и доминируют в сухих лугах на высоких террасах, а также находят свой оптимум в структуре флоры остепненных дубрав ( Artemisio gmelinii - Quercetum mongolicae ass. nov.). Широкоареальные лугово-лесные виды – одна из наиболее распространенных групп видов для флор свежих, сухих лугов, а также лесных фитоценозов на гривах террас.
Выводы
Изучение ботанического разнообразия долинных комплексов – одна из актуальных задач современной географии биоразнообразия. Долинная часть Хинганского заповедника может быть использована в качестве модельной по отношению ко всей Архаринской низменности, в силу слабой нарушенности растительного покрова она является очень ценным объектом для выявления закономерностей дифференциации биоразнообразия.
Флора Архаринской низменности включает в себя 410 видов растений, относящихся к 91 семейству. Систематизация растительности Антоновского лесничества на принципах флористической классификации по Браун-Бланке позволила выделить 12 ассоциаций, относящихся к 5 классам и 7 порядкам.
Согласно результатам экологической ординации синтаксонов по методу шкал Раменского, они располагаются в условном экологическое поле, в котором условия увлажнения меняются от увлажнения сильнообводненных болот до лугово-степного или влажностепного, а условия богатства почв от бедных почв до богатых бурых, луговых, достаточно обеспеченных элементами минерального питания почв. Видовое богатство и видовая насыщенность растительности взаимосвязаны с изменением условий увлажнения и богатства почв, достигают наибольших значений для растительных ассоциаций грив и террас (луговой и лесной ряды), а минимальные – для растительных ассоциаций обводненных прарусловых понижений (болотный ряд) на пойме. Анализ важнейших показателей разнообразия по градиенту увлажнения и богатства почв позволил установить закономерности распределения отдельных типологических категорий, выявить связи различных элементов с определенными генетическими частями долины.
