Разложение сидератов и высвобождение питательных элементов в условиях субарктики, Кольский полуостров

Автор: Кашулина Г.М., Чуева Н.В., Литвинова Т.И., Никерова К.М.
Журнал: Бюллетень Почвенного института им. В.В. Докучаева @byulleten-esoil
Рубрика: Статьи
Статья в выпуске: 119, 2024 года.
Бесплатный доступ
В полевом эксперименте изучали интенсивность разложения и высвобождения питательных элементов для шести видов растительных остатков - листьев, метелок и соломы овса (Avena sativa), а также целых растений люпина узколистного (Lupinus angustifolius), фацелии пижмолистной (Phacelia tanacetifolia) и вики посевной (Vicia sativa) в пахотном горизонте агроподзола на Кольском полуострове. Образцы в сетчатых капроновых мешочках извлекались по истечении 8, 12 и 24 месяцев экспозиции в почве. В растительных остатках определяли потерю веса, содержание Сорг и Nорг, а также зольных элементов - Si, Al, Fe, K, Ca, Mg, P, S и Mn. Как показали результаты, несмотря на низкие температуры, разложение сидератов в агроподзоле протекало с высокой интенсивностью, сопоставимой с таковой в южной черноземной зоне: после года экспозиции в почве растительные остатки теряли от 47 (солома овса) до 75% (люпин) своего исходного беззольного веса. После двух лет - потери веса достигли 68% у соломы овса и 90% у люпина. Распределение потерь Cорг, Nорг и P между видами растительных остатков и по срокам отбора достоверно (при p≤0.01) коррелировало с распределением потерь веса. Изменения содержания других зольных элементов не зависели от потери веса: в зависимости от вида материала и стадии разложения образцы могли не только терять их (K - все виды; Mn - люпин; S, Ca и Mg - все, кроме соломы и метелок овса), но и накапливать с превышением исходного уровня (Fe - все виды, Mn - все виды, кроме люпина; Ca и Mg - солома и метелки овса). Накопление элеменов выше исходного уровня означает, что транформированные растительные остатки могут удерживать дополнительные количества элементов из почвенного раствора, а нерегулярные смены увеличения и снижения содержания Ca, Mg, S, Fe и Mn свидетельствуют об обратимом характере этого процесса. Благодаря значительным различиям между видами сидератов по химическому составу, скорости и динамике разложения и высвобождения питательных элементов их применение предоставляет большие возможности для управления плодородием почв и интенсивностью круговорота элементов в агроценозах за счет подбора культур.
Сидераты, углерод, азот, зольные элементы, разложение растительных остатков, агроподзол, кольский полуостров
Короткий адрес: https://sciup.org/143183305
IDR: 143183305 | DOI: 10.19047/0136-1694-2024-119-30-65
Decomposition of green manure and nutrients release in subarctic, Kola peninsula, Russia
In the field experiment, the intensity of decomposition and release of nutrients was studied for six types of plant residues - leaves, panicles and straw of oats (Avena sativa), as well as whole plants of lupine (Lupinus angustifolius), phacelia (Phacelia tanacetifolia) and vetch (Vicia sativa) in the arable horizon of Anthrosols on the Kola Peninsula. Samples in mesh nylon bags were sampled after 8, 12 and 24 months of exposure in soil. The loss of weight, the content of Corg and Norg, as well as ash elements - Si, Al, Fe, K, Ca, Mg, P, S and Mn were determined in all plant residue samples. As the results showed, despite the low temperatures, the decomposition of green manure in soil occurred with a high intensity, comparable to the southern chernozem zone: after a year of exposure in the soil, plant residues lost from 47% (oat straw) to 75% (lupine) of their initial ash-free weight. After 2 years, weight loss reached 68% in oat straw and 90% in lupine. The distribution of losses of Corg, Norg, and P between the types of plant residues and decomposition periods correlated significantly (at p≤0.01) with the distribution of weight losses. Changes in the content of other ash elements did not depend on weight loss: depending on the type of material and the stage of decomposition, the samples could not only lose them (K - by all plant residues; Mn - by lupine; S, Ca and Mg - by all plant residues, except straw and oat panicles), but also accumulate them in quantities exceeding the initial ones (Fe - by all species, Mn - by all species except lupine; Ca and Mg - by straw and oat panicles) at certain periods. The accumulation of elements above initial level means that the transformed plant residues can accumulate additional amounts of elements from the soil solution, while the irregular alternation of increases and decreases in Ca, Mg, S, Fe and Mn indicates the reversible nature of the process. Due to the significant differences between the types of green manure in terms of chemical composition, rate and dynamics of decomposition and release of nutrients, their use provides great opportunities for managing soil fertility and the intensity of elements cycling in agrocenoses through the species selection.
Текст научной статьи Разложение сидератов и высвобождение питательных элементов в условиях субарктики, Кольский полуостров
184209, Апатиты, Мурманская обл., ул. Ферсмана, 18а, *, e-mail:
18a Fersman st., Murmansk reg., Apatity, 184209, Russian Federation, *, e-mail:
Разложение растительных остатков является важнейшим экосистемным процессом, который, в том числе, регулирует круговорот питательных элементов и плодородие почв (Александрова, 1980; Гришина, 1986). Лесные почвы на Кольском полуострове – подзолы – характеризуются очень низким естественным плодородием (Переверзев, 2004) и даже после окультуривания для поддержания их плодородия необходимо ежегодное внесение очень больших доз органических (порядка 40–90 т/га) и минеральных (N120P80K80) удобрений (Переверзев, 1993; Елсаков и др., 2001 и др.). Зеленые удобрения считаются наиболее экологичным видом органических удобрений (например, Довбан, 2009; Green manure…, 2011; Лошаков, 2018). Однако исследований по эффективности их применения на Кольском полуострове не проводилось. Одним из факторов, лимитирующих возможности применения сидератов в агроподзолах, могут быть суровые климатические условия. Характерные для региона низкие температуры и недостаток влаги в отдельные периоды (Семко, 1982) могут замедлять разложение, поскольку температура и влажность являются основными внешними факторами, определяющими интенсивность разложения растительных остатков в почве (Zhang et al., 2008; Prescott, 2010; Ларионова и др., 2017; Семенов и др., 2022). Изучение процессов трансформации растительных остатков в окультуренных подзолах на Кольском полуострове представляет не только практический, но и большой теоретический интерес, поскольку для данных почв характерно специфическое сочетание внешних природных факторов, которые контролируют интенсивность разложения: холодный гумидный климат, аэробные условия, кислая среда, легкий гранулометрический состав и кислый состав почвообразующих пород. В данной статье представлены результаты изучения интенсивности разложения и динамики высвобождения питательных элементов в процессе трансформации нескольких видов сидератов в субарктическом агроподзоле при таком специ-фичесоком сочетании природных факторов.
ОБЪЕКТЫ И МЕТОДЫ
Эксперимент по разложению растительных остатков (РО) сидератов проводился в пахотном горизонте агроподзола на опытном участке Полярно-альпийского ботанического сада-института
(ПАБСИ) КНЦ РАН в окрестностях г. Апатиты (67°34'46.0" N; 33°18'01.2" E). Для региона характерен арктически-умеренный климат, смягченный теплым Северо-Атлантическим течением Гольфстрим. Поэтому на большей части территории вечная мерзлота отсутствует, и почвы являются длительно-сезонно-промерзающими. Преобладающими почвообразующими породами здесь являются ледниковые песчаные отложения кислого состава (Переверзев, 2004). Агроподзол сформирован на рыхлых озерноледниковых отложениях.
По данным предшествующего обследования 44 площадок (Кашулина и др., 2021; Чуева и др., 2021), пахотный горизонт окультуренного подзола на экспериментальном участке ПАБСИ характеризовался низким содержанием органического углерода (здесь и далее – среднее ± стандартное отклонение: 2.2 ± 0.7%) и азота (0.14 ± 0.03%), слабокислой реакцией среды (рНKCl = 5.6 ± 0.2), низким содержанием обменных форм Ca (5.6 ± 1.7 смоль(экв)/кг) и Mg (0.39 ± 0.33 смоль(экв)/кг), а также низкой ЕКО (8.1 ± 2.3 смоль(экв)/кг). Пахотный горизонт также характеризуется легким гранулометрическим составом (доля физической глины в мелкоземе составляла 10 ± 2.4%), высокой каменистостью (доля камней и гравия составила 18 ± 3%), рыхлым сложением (V = 0.92 ± 0.13 г/см3) (Кашулина и др., 2021; Чуева и др., 2021), низкой влагоемкостью (ПВ = 58.5%, НВ = 24%) (Семко, 1982) и промывным водным режимом (Семко, 1982; Переверзев, 1987; Переверзев, 1993). Коэффициент увлажнения за 9 лет (с 1971–1979 гг.) составил 1.6 ± 0.33 (Семко, 1982). Погодные условия во время проведения эксперимента, по данным метеостанции Апатиты, расположенной в 4 км от места проведения исследований , представлены на рисунке 1.
Участок, где непосредственно проводили эксперимент, 2 предшествующих года не использовался под посадки. Во время эксперимента сорняки регулярно удаляли. Пахотный горизонт на этом участке характеризовался низким содержанием органического вещества (потеря при прокаливании при t 600 °С = 6.4 ± 0.3% (N = 6) приблизительно соответствует 1% С орг ), слабокислой реакцией среды (рНKCl = 5.5 ± 0.04, рН Н2О = 6.4 ± 0.08), очень высоким содержанием подвижного фосфора по Кирсанову (P 2 O 5 =
62.3 ± 14.1 мг/100 г), средним – подвижного калия (K 2 O =
10 ± 3 мг/100 г), очень низким – аммиачного азота (N-NH 4 = 0.24 ± 0.08 мг/100 г, нитратный азот отсутствовал.
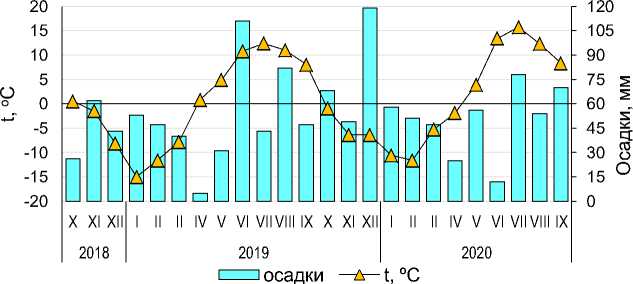
Рис. 1. Количество осадков и средняя за месяц температура воздуха во время проведения эксперимента по разложению.
Fig. 1. The amount of precipitation and the average air temperature for the month during the decomposition experiment.
Влажность пахотного горизонта агроподзола перед началом эксперимента составила 18.5 ± 1.2 весовых %. Однако в процессе трансформации РО поддерживали значительно более высокую влажность по сравнению с окружающей их почвой (рис. 2).
Воздушно-сухие образцы листьев (ОвЛ), метелок (ОвМ) и соломы овса (ОвС) (Avena sativa, сорт Хибинский) на стадии молочной спелости, а также целых растений люпина узколистного (Lupinus angustifolius, сорт Белозерный 110) – Лп, фацелии пижмолистной (Phacelia tanacetifolia, сорт Рязанская) – Фц и вики посевной (Vicia sativa, сорт Льговская-2) – Вк на стадии начала цветения разрезали на кусочки по 5 см и помещали в ячеистые капроновые мешочки размером 15 × 15 см. Мешочки с растительными остатками были заложены горизонтально на глубине 10 см пахотного горизонта 23 сентября 2018 г. и извлекались через 8 месяцев (14 мая 2019 г., I срок), через год (18 сентября 2019 г., II срок) и через 2 года (23 сентября 2020 г., III срок) в 3–10-кратной повтор- ности.
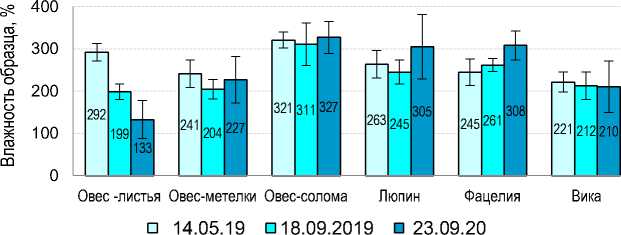
Рис. 2. Влажность экспонировавшихся в почве образцов растительных остатков.
Fig. 2. Moisture level of plant residue exposed in the soil.
Подготовка и химический анализ растительных остатков . Для химического анализа индивидуальные образцы одного вида РО и одного срока отбора объединялись в один смешанный образец. В исходном и в экспонировавшихся смешанных образцах растений определяли зольность (озоление при температуре 450 °С), зольные элементы (Si, Al, Fe, Ca, Mg, K, P, Mn и S) по прописи Поповцевой (Поповцева, 1974), а также С орг по методу Тюрина и N орг по Кьельдалю. Концентрация Si в использованной методике представляла сумму свободных (обработка золы 5%-ной HCl) и аморфных (обработка остатка 5%-ным раствором KOH) форм.
Углеводный состав листьев и стеблей, был определен на научном оборудовании Центра коллективного пользования ФИЦ “Карельский научный центр РАН”. Лигнин и целлюлозу определяли гравиметрическим методом после выделения лигнина 72%-ной серной кислотой, целлюлозы – по методу Кюршнера-Хоффера смесью концентрированной азотной кислоты и этилового спирта в соотношении 1 : 4 (по объему). Глюкозу определяли глюкозоокси-дазным методом со спектрофотометрическим окончанием; фруктозу, сахарозу – по реакции с резорцином со спектрофотометрическим окончанием; крахмал – по количеству образовавшейся глю- козы глюкозооксидазным методом.
Обработка данных была проведена с использованием Microsoft Excel 2010. В тексте приведены только достоверные при p≤0.01 коэффициенты корреляции. Для оценки достоверности различий между наборами данных использовали U-критерий Манна– Уитни . При обсуждении материала были использованы 2 понятия: концентрация элемента в % или мг/кг на абсолютно-сухой вес (а. с. в.) и содержание элемента в образце в мг (концентрация * вес образца).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Элементный и биохимический состав исходных образцов
Элементный и биохимический состав растений определяется их видовыми особенностями (Ильин, 1985; Markert, 1992; Nelson, Cox, 2005). Сидераты, использованные для эксперимента, значительно различались по химическому (рис. 3, табл. 1) и биохимическому составу (рис. 4), которые в значительной степени влияют на интенсивность их разложения (Александрова, 1980; Гришина, 1986; Шлегель, 1987).
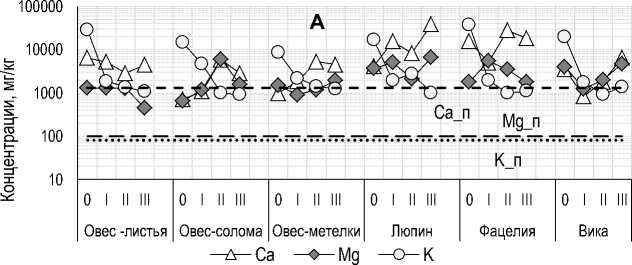
Элементный состав растений . Концентрация C орг (% а. с. в) варьировала от 39 (листья овса) до 44% (солома овса и люпин), N орг – от 2.8 (фацелия) до 6.1% (люпин). Межвидовые различия по содержанию зольных элементов были более значительными (табл. 1): от трех раз по P до 100 раз и более по Mn. Преобладающим зольным элементом во всех видах РО был K. За счет дополнительного извлечения аморфных форм второе место во всех частях овса занял Si. При этом доля его аморфных форм составила 80–90%. У травянистых растений общая концентрация Si была значительно ниже, по сравнению с K, Ca, Mg, P и S, а доля его аморфных форм варьировала от 20% в вике до 50% в люпине, т. е. с растительными остатками в почву поступают не только значительные количества важных питательных – N, K, Ca, Mg, P и S, – но и геогенных элементов, особенно Si.
А
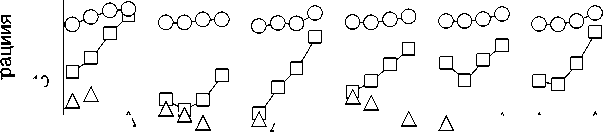
ф
△△△
△
0 I II III
0 I II III
0 I II III
0 I II III
0 I II III
АГ ”\Г
△
0 I II III
Ф
-20
-40
-60
-80
-100
-120
Люпин
Фацелия
Вика
Сорг
А Nорг
Овес -листья
Овес-метелки
Овес-солома Зольность
Б
I II III
I II III
I II III
I II III
I II III
Овес-листья
I II III ес- лки
Овес-солома
Зола
Люпин
Фацелия
Вика
Б/з вес
О Сорг
Nорг
Рис. 3. А – Изменение зольности (% на а. с. в.) и концентрации С орг и N орг (% на беззольный вес); Б – потери общего веса, золы и С орг и N орг (% от исходного) в процессе трансформации растительных остатков в почве: О – исходные образцы, I – после первых 8 месяцев экспозиции в почве, II – после 1 года экспозиции и III – после 2 лет.
Fig. 3. A – Change in ash content (% of dry weight) and concentration of C org and N org (% of ashless weight); Б – loss of total weight, ash and content of C org and N org (% of initial) in the process of transformation of plant residues in the soil: O – initial samples, I – after the first 8 months of exposure in the soil, II – after 1 year of exposure and III – after 2 years.
Таблица 1. Зольность (% на а. с. в.) и концентрация зольных элементов (г/кг или мг/кг на а. с. в.) в растительном материале, используемом для эксперимента
Table 1. Ash content (% of a. d. w.) and concentration of ash elements (g/kg or mg/kg of a. d. w.) in the plant material used for the experiment
|
Образец |
Зола, % |
г/кг |
мг/кг |
|||||||
|
K |
Ca |
Mg |
P |
S |
Si |
Al |
Fe |
Mn |
||
|
Овес-листья |
12.8 |
28.4 |
6.4 |
1.3 |
2.4 |
1.8 |
12.7 |
5.0 |
267 |
0.5 |
|
Овес-солома |
6.1 |
14.7 |
0.7 |
0.6 |
1.2 |
0.3 |
4.4 |
2.0 |
51 |
23.6 |
|
Овес-метелки |
4.2 |
8.7 |
1.0 |
1.5 |
2.9 |
0.3 |
7.9 |
0.8 |
88 |
44.0 |
|
Люпин |
7.5 |
16.9 |
3.5 |
4.1 |
2.9 |
1.5 |
0.7 |
2.2 |
278 |
18.6 |
|
Фацелия |
16.1 |
37.0 |
15.8 |
1.8 |
3.6 |
1.4 |
1.0 |
3.5 |
134 |
66.0 |
|
Вика |
10.1 |
19.8 |
4.1 |
3.7 |
3.3 |
1.1 |
1.0 |
1.5 |
607 |
2610 |
|
Макс/мин |
4 |
4 |
22 |
7 |
3 |
7 |
19 |
5.9 |
12 |
>100 |
Углеводный состав. Трудно разлагаемые компоненты – целлюлоза и лигнин – были преобладающими компонентами и в стеблях, и в листьях, всех обследованных сидератов (рис. 4). Для всех компонентов, кроме крахмала, было свойственно более высокое содержание в стеблях по сравнению с листьями: целлюлозы – от 1.3 раза (овес и вика) до 2.6 раз (люпин); лигнина – от 1.3 раза (овес) до 3 раз (фацелия); фруктозы – от приблизительно равного (люпин, фацелия и вика) до 3 раз (овес); и глюкозы – от приблизительного равного (вика) до 3 раз (фацелия). Содержание крахмала в листьях фацелии, овса и люпина было выше, чем в стеблях в 10, 3 и 2 раза соответственно. У вики его содержание в стеблях было, наоборот, в 2.5 раза выше, чем в листьях.
Различия между видами сидератов по содержанию некоторых углеводов в стеблях и листьях были более значительны, чем между листьями и стеблями одного вида. Особенно сильно виды различались по содержанию в стеблях фруктозы (в 27 раз) и крахмала (300 раз). Для листьев эти различия составили 5 и 50 раз для фруктозы и крахмала, соответственно. По содержанию преобладающих, трудноразлагаемых компонентов виды сидератов разли- чались в меньшей степени: по целлюлозе – в 1.2 раза в стеблях и в ≈2 раза в листьях, по лигнину – в 1.4 раза в стеблях и в 2.4 раза в
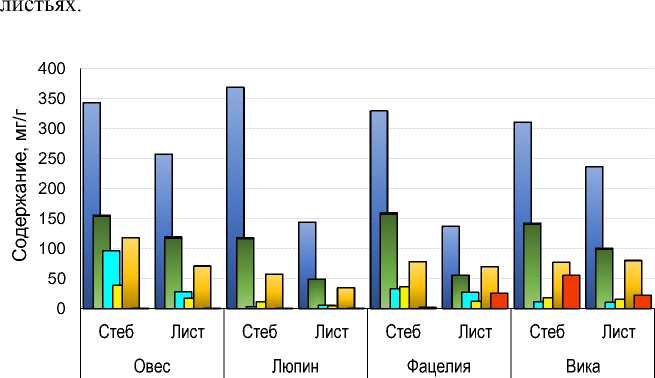
□ Целлюлоза □ Лигнин □ Фруктоза □ Глюкоза □ Сахароза □ Крахмал
Рис. 4 . Углеводный состав листьев и стеблей 5 видов сидератов.
Fig. 4. Carbohydrate composition of leaves and stems of 5 types of green manure.
Трансформация растительных остатков в почве
Из-за специфики биохимического состава различных органов, тканей и компонентов клеток изменение внешнего вида РО в процессе их трансформации в почве зависит от морфологического и анатомического сложения растений (Александрова, 1980; Гришина, 1986). Внешние изменения некоторых видов РО в процессе их трансформации в почве представлены на риcунках 5–8. Листья, характеризующиеся менее прочным строением стенок клеток и более низким содержанием трудноразлагаемых компонентов – целлюлозы и лигнина – разлагались значительно быстрее с постепенным увеличением степени фрагментации до мелкой пыли после 2 лет экспозиции (рис. 7 и 8). Тогда как стебли всех растений сохраняли целостность и объем все 2 года (рис. 5, 7 и 8). При значительной потере веса (рис. 3) это может означать, что объем сохранялся за счет каркаса из стенок клеток, состоящих из более устойчивых компонентов (Смирнова, Кочетов, 2015), и что, как минимум, преобладающая часть остающегося в почве Сорг продолжает находиться в составе органического вещества крупных неразложившихся фрагментов растений.
Наиболее значительное потемнение и разрушение стеблей было приурочено к концам отрезков (рис. 5, 7 и 8). Очевидно, что защита от проникновения патогенов, созданная живым растением, – эпидермис со специфическим биохимическим составом стенок клеток (Смирнова, Кочетов, 2015) – затрудняет проникновение микроорганизмов-деструкторов внутрь растительных остатков при их попадании в почву.

Рис. 5. Внешний вид образцов стеблей овса ( Avena sativa , Khibiny variety), экспонировавшихся в почве в течение 8 месяцев, 1 года и 2 лет . Fig. 5. Appearance of oats stems samples ( Avena sativa , Khibiny variety) exhibited in the soil for 8 months, 1 year and 2 years .

Рис. 6. Внешний вид образцов метелок овса ( Avena sativa , сорт Хибинский), экспонировавшихся в почве в течение 8 месяцев, 1 года и 2 лет.
Fig. 6. Appearance of samples of oat panicles ( Avena sativa , Khibiny variety) exhibited in the soil for 8 months, 1 year and 2 years.

Рис. 7. Внешний вид образцов люпина узколистного ( Lupinus angustifolius , сорт Белозерный 110), экспонировавшихся в почве в течение 8 месяцев, 1 года и 2 лет.
Fig. 7. Appearance of the samples of lupine ( Lupinus angustifolius, Belozerniy 110 variety) exhibited in the soil for 8 months, 1 year and 2 years.

Рис. 8. Внешний вид образцов фацелии пижмолистной ( Phacelia tanacetifolia , сорт Рязанская), экспонировавшихся в почве в течение 8 месяцев, 1 года и 2 лет.
Fig. 8. Appearance of samples of phacelia tanacetifolia ( Phacelia tanacetifolia , Ryazanskaya variety) exhibited in the soil for 8 months, 1 year and 2 years.
Измельчение и нарушение целостности растений перед их внесением в почву может быть одним из наиболее эффективных механизмов управления интенсивностью их разложения. Так, увеличение степени измельчения от 20-сантиметровых отрезков до фрагментов размером менее 1 мм в эксперименте с соломой зерновых культур (Ловчиков и др., 2016) сопровождалось 4-кратным увеличением скорости ее разложения в почве, тогда как виды растений по интенсивности разложения различались всего в 2 раза (см. ни- же).
Изменение зольности растительных образцов в процессе разложения было индивидуальным для каждого вида растительного материала (рис. 3) и зависело от соотношения потерь одних элементов, особенно K (доля K 2 O в золе исходных образцов составляет 25–30%), от иммобилизации на месте высвободившихся при разложении элементов (Si, Al, Fe и Ca) и накопления почвенных частиц со временем. В большинстве экспонировавшихся образцов, кроме соломы овса (низкая исходная концентрация K, слабая способность накапливать почвенные частицы из-за обтекаемой формы), фацелии (самая высокая исходная концентрация K) и вики, после первых 8 месяцев экспозиции в почве зольность была значительно ниже исходной.
Потеря веса . Из-за значительного изменения зольности снижение общего веса образцов не совсем адекватно характеризовало интенсивность разложения органического вещества РО. Поэтому в нашем эксперименте скорость его разложения была оценена по беззольной навеске (рис. 3). Согласно результатам многочисленных исследований (Переверзев, 1987; Ушакова, 1983; Cornwell et al., 2008; Дедов и др., 2016; Семенов и др., 2019 и др.), при экспонировании РО в одинаковых условиях вид растений становится ведущим фактором, определяющим скорость их разложения. В данном эксперименте потери беззольного веса соломой и метелками овса во все сроки отбора образцов, согласно U -критерию Манна–Уитни (при p≤0.01), были значимо ниже по сравнению с листьями овса и целыми растениями люпина, фацелии и вики. Межвидовые различия наиболее четко проявились на начальных стадиях разложения и сглаживались со временем. Так, за первые 8 месяцев экспозиции в почве потери беззольного веса варьировали от 30% у соломы овса, до 60% у листьев овса и целых растений фацелии и вики (т. е. в 2 раза). К концу 2-го года потери достигли 68% у соломы овса, и около 90% у листьев овса, люпина и вики, т. е., по кумулятивным потерям за 2 года различия между видами уменьшились до 1.3 раза.
Несмотря на низкие температуры, интенсивность разложе- ния РО в субарктическом агроподзоле оказалась сопоставима с таковой в южных регионах. Например, солома озимой пшеницы в сходном по методике эксперименте (капроновые мешочки, отрезки 5–7 см) после года разложения в типичном черноземе Воронежской области потеряла только 25% своего исходного веса, люцерна – 66%, а потери после двух лет экспозиции составили 72 и 88% для соломы пшеницы и люцерны, соответственно (Дедов и др., 2016).
Из-за специфичного для каждого вида растений соотношения различных по устойчивости к разложению групп органического вещества (Семенов и др., 2019) виды РО значительно различались и по скорости изменения интенсивности разложения со временем. Как следует из убывающих рядов потерь беззольного веса, (в пересчете на месяц для каждого периода экспозиции, в % от исходного) интенсивность разложения органического вещества между видами РО во все периоды отличалась в 2 раза:
I этап:
ОвЛ (7.8) > Фц (7.2) > Вк (7.1) > Лп (6.9) > ОвМ (4.4) > ОвС (4.0) II этап:
ОвМ (7.2) > Лп (5.0) > ОвЛ (4.7) > Вк (3.8) ≥ ОвС (3.7) = Фц (3.7) III этап:
ОвС (1.8) > ОвМ (1.4) > Вк (1.3) > Лп (1.2) > Фц (1.0) > ОвЛ (0.8)
Однако из-за различий между видами в сочетании трудно- и легкоразлагаемых компонентов их распределение по этому показателю менялось со временем. Наиболее интенсивно разложение во всех видах РО, кроме метелок овса, протекало в первые 8 месяцев экспозиции с самыми высокими потерями беззольного веса у листьев овса и травянистых растений, а с наименьшими – у соломы и метелок овса. В последующие четыре летних месяца у листьев овса, целых растений фацелии, вики и люпина интенсивность разложения снизилась незначительно (в 1.5–2 раза), у соломы овса – практически осталась на том же уровне, а у метелок овса, наоборот, увеличилась в 1.6 раза. Последнее было обусловлено тем, что легкоразлагаемые компоненты зерна стали доступны для микроорганизмов только после частичного разложения прочной семенной оболочки в первые 8 месяцев (рис. 6). В летний период самая высокая интенсивность разложения была свойственна метелкам овса, самая низкая – соломе овса и фацелии.
Наиболее значительное снижение интенсивности разложения, по сравнению с первым периодом, произошло на второй год экспозиции: в 9 раз – у листьев овса, в 7 раз – у фацелии, около 6 раз – у вики и люпина и в 2 раза – у соломы овса. В этот период наиболее высокая интенсивность разложения была свойственна медленно разлагаемым РО (солома и метелки овса), наиболее низкая – быстро разлагающимся (листья овса и травянистые растения).
Изменение концентрации элементов в РО . Эффективность применения сидератов для повышения плодородия почв и экологические последствия применения сидератов зависят от поведения конкретного элемента в процессе трансформации РО в конкретной почвенно-геохимической обстановке. В отличие от лабораторных экспериментов, концентрация элемента в РО в полевом эксперименте зависит не только от свойств элемента и изменения его подвижности, как в связи со сменой окислительновосстановительных условий, так и со способностью взаимодействовать с продуктами трансформации РО, но и от многих других факторов: например, таких как накопление почвенных частиц и микробной биомассы, значительно отличающихся по химическому составу от РО, а также вынос/поступление элементов с фильтрующимися почвенными растворами. Некоторое представление о поведении элементов при разложении сидератов в пахотном горизонте агроподзола дает анализ изменения их концентрации со временем.
Сорг. Из-за значительного повышения зольности направленность и степень изменения концентрации Cорг и Nорг по мере трансформации РО в почве была охарактеризована в % на беззольный вес (рис. 3). В исходных образцах концентрация Сорг варьировала от 42.8 (метелки овса) до 49.3% (фацелия) на б/з вес. Степень и динамика изменения концентрации Сорг были индивидуальными для каждого вида РО. Однако в большинстве случаев концентрация Сорг в экспонировавшихся образцах РО была выше исходной. После первых 8 месяцев экспозиции в почве концентрация Сорг значительно возросла, по сравнению с исходной, только в листьях и метелках овса (на 10 и 4% на б/з вес, соответственно). На более поздних сроках экспозиции концентрация Сорг была выше исходной во всех видах РО, особенно после второго года экспозиции. Всего за 2 года экспозиции концентрация Сорг увеличилась на 4% у соломы овса, на 18% у метелок овса и вики и до 22% у листьев овса. Причинами общей тенденции концентрации Сорг к увеличению может быть относительное обогащение РО в процессе их трансформации трудно разлагаемым лигнином и вновь образованными гуминовыми кислотами, которые характеризуются более высоким содержанием Сорг (Шлегель, 1987; Гришина, 1986).
N орг . Различия по исходной концентрации N орг (рис. 3) между видами РО (варьировала от 3.3 у фацелии до 6.6% на б/з вес у люпина) и ее изменения в процессе их трансформации в почве были более значительными (от ≈ 1.5 раз у видов с низкой исходной концентрацией азота – метелки овса, фацелия, вика, до более 2 раз для видов с его высокой исходной концентрацией – люпин, листья и солома овса) по сравнению с С орг . После первых 8 осенне-зимних месяцев концентрация N орг незначительно снизилась во всех видах РО, кроме листьев овса: на 0.3% у вики и на 1% у люпина. Во всех частях овса концентрация N орг продолжала уменьшаться до конца эксперимента с наиболее резким снижением (в 2.4 раза) у листьев овса за 4 теплых летних месяца, а у соломы овса (в 1.8 раза) за время второго года экспозиции. У травянистых растений концентрация N орг снизилась до минимума после 1-го года экспозиции и снова увеличилась к концу 2-го года, превысив исходно низкие значения у вики и фацелии. На нерегулярные изменения концентрации азота в РО могло оказать влияние чередование ассимиляции микроорганизмами высвобождающегося при разложении растений азота и его последующее высвобождение при их отмирании (Кузнецова и др., 2006).
Снижение концентрации Nорг и обеднение органического вещества РО азотом в процессе их трансформации в почве обусловлено более быстрым разложением белков с высоким содержа- нием азота и увеличением доли трудно разлагаемых компонентов – лигнина и гуминовых кислот – с высоким содержанием углерода.
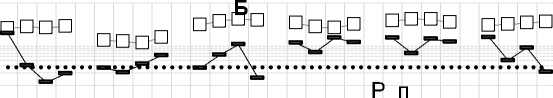
Зольные элементы: P и S также входят в состав молекул органического вещества растений, но в процессе трансформации РО в почве эти элементы вели себя по-разному (рис. 9). Направленность изменения концентрации P зависела от вида РО и срока отбора: в 12 образцах из 18 концентрация была равна или выше исходной (во все сроки отбора у вики, листьев и метелок овса, в отдельные сроки у соломы овса и фацелии) и в 6 случаях – ниже (во все сроки отбора у люпина, в отдельные сроки – у соломы овса и фацелии). Однако степень изменения концентрации P относительно исходной была незначительной: максимальное повышение было в 1.4 раза (метелки овса после 1-го года экспозиции), максимальное снижение – в 1.3 раза (у люпина после 1-го года).
Изменение концентрации S в РО по мере их трансформации в почве были более значительными по сравнению с P, а направленность изменений чаще всего зависела от периода экспозиции (рис. 9). После первых 8 осенне-зимних месяцев экспозиции в почве концентрация S снизилась во всех видах РО, кроме метелок овса. Наиболее значительное ее снижение произошло у быстро разлагающихся видов с высоким исходным содержанием S: в 6 раз у листьев овса, почти в 4 раза у вики и в 2.4 раза у фацелии. За последующие четыре летних месяца во всех видах РО, кроме листьев овса, концентрация S, наоборот, существенно увеличилась по сравнению с предыдущим сроком отбора образцов. Наиболее значительное повышение в этот период было отмечено у видов с низким исходным содержанием: в 4 раза в метелках овса и в 2 раза в соломе. В большинстве видов РО, кроме листьев и соломы овса, за время 2-го года экспозиции концентрация S вновь снизилась, достигнув минимума в метелках овса и вике.
Наиболее значительно в процессе разложения в образцах РО снижалась концентрация главного зольного элемента – калия (K) .
г 1000
0 I II III
Овес -листья
0 I II III
Овес-солома
0 I II III
Овес-метелки
□ P
0 I II III
0 I II III
Фацелия
0 I II III
Вика
Люпин S
г 1000
В
0 I II III
Овес -листья

0 I II III
Овес-солома
0 I II III
Овес-метелки
0 I II III
Люпин
0 I II III
Фацелия
0 I II III
Вика
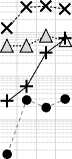
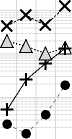
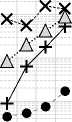
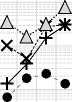
X Si —I Fe ......• △ Al • Mn
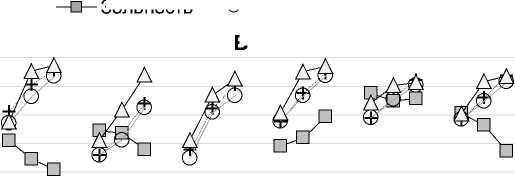
Рис. 9. Изменение концентрации зольных элементов в процессе трансформации растительных остатков в почве: А – Ca, Mg и K; Б – P и S; В – Si, Al, Fe, Mn. Обозначения на оси Х см. на рис. 3.
Fig. 9. Changes in the concentration of ash elements during the transformation of plant residues in the soil: A – Ca, Mg и K; Б – P и S; В – Si, Al, Fe, Mn. For the designations of the X-axis, see Fig. 3.
Уже после первых 8 месяцев экспозиции в почве его концентрация в РО снизилась в 3–4 раза у медленно разлагающихся видов с низкой исходной концентрацией (солома и метелки овса) и в 15–19 раз у быстро разлагающихся видов с высокой исходной концентрацией (листья овса и фацелия) (рис. 9). Такое поведение K обусловлено ионной формой его присутствия в растениях и отсутствием механизма у трансформированных РО к его удержанию.
В отличие от K, концентрация Ca и Mg в разлагающихся РО могла быть как выше, так и ниже исходной. Наиболее значительное снижение концентрации Са было отмечено у вики (в 4 раза) и у фацелии (в 3 раза) после первых 8 месяцев экспозиции в почве. В 1.5–2.3 раза ниже исходной она была в листьях овса на протяжении всего эксперимента. Наоборот, в большинстве экспонировавшихся в почве образцов РО (в 12 из 18) концентрация Ca была значительно выше исходной: от 1.5 до 8 раз в соломе овса, от 2 до 5 раз в метелках овса и от 2 до 9 раз в люпине.
Снижение концентрации Mg относительно исходной было отмечено в отдельные сроки отбора образцов у вики (в 2–3 раза), у метелок овса (в 1.3–1.6 раза) и у люпина (в 1.7 раза). Наоборот, значительное повышение его концентрации (в 9 раз) было свойственно соломе овса и фацелии (в 3 раза) после первых 8 месяцев экспозиции.
Нерегулярные изменения концентрации и содержания Ca и Mg в процессе трансформации РО предполагают наличие механизма их временного закрепления в образцах. Таким механизмом для этих элементов может быть обменное связывание карбоксильными группами вновь образующихся при трансформации РО гумусовых веществ (Шлегель, 1987; Мальцева и др., 2013; Столбовой, 2022). Способность трансформированных РО связывать Ca и Mg, наиболее четко проявившаяся у соломы овса, предотвращает вынос этих важных питательных элементов и сохраняет их в доступной для растений форме.
Сравнение уровней концентраций элементов в РО и в почве участка (рис. 9, пунктирные линии для почв) показало, что даже после 2 лет экспозиции в почве концентрации важных питательных элементов в РО находились на уровне, значительно превосхо- дящем концентрации подвижных форм этих элементов в почве: Nорг – от 10 раз у листьев и соломы овса до ≈20 раз у люпина; доступного P – от 5 у соломы овса до 13 раз у вики; доступного K – от 11 раз у соломы овса ≈17 раз у вики; обменного Ca – от 2 у соломы овса до ≈30 раз у фацелии; обменного Mg – от 15 раз у соломы овса до >60 раз у фацелии. Однако внесения 200 г/м2 воздушно-сухих РО (соответствует средней урожайности с 1 м2 в регионе) при массе пахотного горизонта ≈200 кг/м2 не было бы достаточно, чтобы оказать какое-либо ощутимое влияние на показатели плодородия почвы.
Марганец (Mn) – важный и дефицитный микроэлемент c самым низким содержанием из всех зольных элементов в обследованных растениях, кроме люпина (табл. 1). Учитывая высокую подвижность Mn в большинстве типов почв (Kabata-Pendias, Mukherjee, 2007), также и в почвах региона (Манаков, 1969), предполагалось, что разложение растительного материала в почвах с промывным водным режимом будет сопровождаться снижением его концентрации. Однако во всех образцах, кроме соломы овса, после первых 8 месяцев экспозиции, она была выше исходной (рис. 9). Наиболее значительное повышение концентрации Mn было свойственно растениям с его низкой исходной концентрацией: более чем в 100 раз в листьях овса и в 4–25 раз в вике. У люпина с самой высокой исходной концентрацией это повышение было незначительным – в 1.1–1.3 раза. Только для соломы и метелок овса было характерно постепенное повышение концентрации Mn cо временем. У остальных видов РО изменения его концентрации со временем не имели регулярного характера. Что свидетельствует о наличии механизма временной иммобилизации Mn в трансформированных остатках. Повышение концентрации Mn в процессе трансформации РО в почвах региона уже наблюдалось ранее в эксперименте по разложению листьев березы в подстилке подзолов (Ушакова, 1983).
Изменение концентрации Si и Al по мере трансформации РО в почве для большинства видов, кроме Si в люпине и Al в люпине и метелках овса, не имело регулярного характера (рис. 9). Относительно небольшое снижение концентрации Si было отмечено в метелках овса (в 1.4 раза) и фацелии (в 2 раза) после первых 8 месяцев экспозиции в почве. В образцах остальных видов РО или при большей продолжительности их нахождения в почве концентрация Si была выше исходной: в 5 раз в вике к концу 1-го года и в 3 раза в соломе овса, люпине, фацелии и вике к концу 2-го года. Концентрация Al была ниже исходной в образцах вики (в 1.6 раза) и фацелии (в 2 раза) после первых 8 месяцев экспозиции в почве, а также во всех образцах соломы овса (в 1.3–2 раза). В двух видах РО с низким исходным содержанием Al (табл. 1) его концентрация значительно возрастала с увеличением продолжительности экспозиции: от 2.4 до 11 раз в метелках овса и от 1.5 до 3 раз в люпине.
Для Fe было характерно значительное повышение концентрации в РО относительно исходной с увеличением продолжительности экспозиции образцов в почве: от 2 (листья овса) до 7 раз (метелки овса) после первых 8 месяцев экспозиции и от 7 (люпин) до ≈60 раз (метелки овса) к концу 2-го года экспозиции. Такое значительное повышение концентрации Fe в РО может быть обусловлено несколькими обстоятельствами: иммобилизацией на месте разложения за счет окисления Fe2+ до малоподвижных оксидов и гидроксидов Fe3+ в окислительных условиях, преобладающих в агроподзолах; связыванием железа, высвобождающегося при разложении, и, возможно, дополнительного количества из почвенного раствора в комплексы со вновь образуемыми гумусовыми веществами; “загрязнением” образцов почвенными частицами при очень низкой его исходной концентрации в растениях и более значительным контрастом с уровнями железа в почве.
Потери элементов в процессе трансформации
Из-за разной направленности и/или степени изменения концентрации потери элементов РО при их трансформации в почве не совпадали (рис. 3 и 9) с потерями общего веса для большинства элементов, кроме С орг , N орг и фосфора (рис. 3, табл. 2).
Распределение потерь Сорг между видами растений и сроками отбора образцов достоверно коррелировало с потерями общего и беззольного веса – r = 0.99 (рис. 3). Однако из-за увеличения концентрации Сорг в РО в процессе их трансформации в почве, по- тери Сорг относительно исходного содержания были ниже по сравнению с беззольным весом, особенно в более поздние сроки разложения. После 1-го года экспозиции в почве РО потеряли от 42 (солома овса) до 73% (листья овса и люпина) Сорг от его исходного содержания в образце. К концу 2-го года они возросли до 65% у соломы овса и до 87% у листьев овса и люпина.
Таблица 2. Потери зольных элементов в процессе трансформации растительных остатков в почве, % от исходного содержания
Table 2. Losses of ash elements in the process of transformation of plant residues in the soil, % of the initial content
|
Сроки |
K |
Ca |
Mg |
P |
S |
Mn |
Si |
Al |
Fe |
|
|
6 3 о g |
I |
97 |
67 |
60 |
55 |
93 |
-618 |
-24 |
60 |
13 |
|
II |
99 |
89 |
75 |
74 |
98 |
-203 |
19 |
57 |
-226 |
|
|
III |
99 |
88 |
94 |
80 |
98 |
-225 |
51 |
77 |
-388 |
|
|
4) 2 о § |
I |
82 |
-1 |
-24 |
38 |
48 |
59 |
-18 |
48 |
-236 |
|
II |
97 |
-337 |
-394 |
55 |
35 |
12 |
45 |
73 |
-520 |
|
|
III |
98 |
-34 |
18 |
60 |
31 |
-172 |
3 |
76 |
-785 |
|
|
О £ S |
I |
95 |
-26 |
59 |
16 |
-40 |
18 |
52 |
-60 |
-384 |
|
II |
98 |
-114 |
68 |
45 |
-48 |
33 |
20 |
-100 |
-708 |
|
|
III |
99 |
-21 |
65 |
65 |
85 |
-3 |
54 |
-184 |
-1479 |
|
|
я я я |
I |
97 |
-72 |
36 |
63 |
73 |
48 |
39 |
32 |
-25 |
|
II |
97 |
45 |
84 |
79 |
64 |
64 |
40 |
48 |
-19 |
|
|
III |
100 |
-23 |
76 |
88 |
86 |
83 |
62 |
60 |
8 |
|
|
и я л е |
I |
97 |
87 |
-20 |
56 |
83 |
-7 |
81 |
82 |
-14 |
|
II |
99 |
51 |
45 |
69 |
73 |
3 |
22 |
75 |
-216 |
|
|
III |
99 |
79 |
82 |
84 |
85 |
64 |
47 |
61 |
-309 |
|
|
я а |
I |
97 |
90 |
87 |
55 |
88 |
-541 |
32 |
73 |
-20 |
|
II |
99 |
84 |
85 |
66 |
83 |
-30 |
-61 |
66 |
-98 |
|
|
III |
99 |
67 |
79 |
78 |
97 |
-358 |
46 |
2 |
-365 |
В абсолютном выражении величина потери образцами Сорг определялась его исходной концентрацией в РО. По оценкам, сде- ланным на основе результатов этого эксперимента (рис. 10), из Сорг, связанного за вегетационный период при создании 200 г сухой биомассы (средняя урожайность с 1 м2) и внесенного в почву осенью, к началу следующего сезона солома овса теряла 28 г, люпин – 49 г. При этом большая часть высвободившегося углерода, предположительно, возвращается в атмосферу в газообразной форме. Поскольку, согласно лизиметрическим исследованиям, потери Сорг за счет выноса с водорастворимыми органическими соединениями за пределы профиля в окультуренном подзоле даже при применении органических удобрений составляли только 1–2 г с 1 м2/год (Переверзев, 1993).
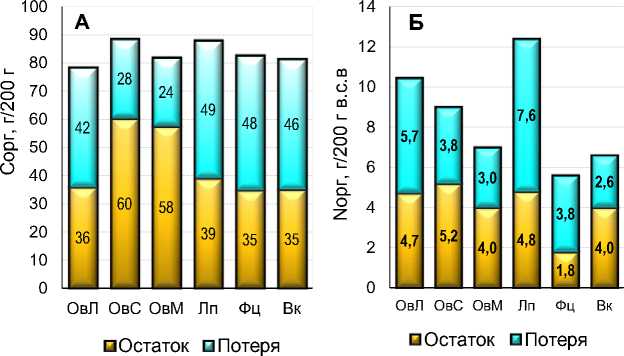
Рис. 10. Распределение между потерей и остатком С орг ( А ) и N орг ( Б ) при разложении сидератов в агроподзоле, в г на каждые 200 г внесенных в почву абсолютно сухих растительных остатков.
Fig. 10. Distribution between loss and residue of C org ( A ) and N org ( Б ) during the decomposition of green manure in the soil, in g per every 200 g of absolutely dry plant residues applied to the soil.
Вместе с тем, несмотря на высокую интенсивность разложения, значительная часть связанного при формировании биомассы сидератов Сорг все же долгое время остается в почве. В годовом балансе (рис. 10) от каждых 200 г внесенных воздушно-сухих РО в почве остается от 35 г (фацелия и вика) до 60 г (солома овса) Сорг. И даже через 2 года его остаток в почве составляет от 10 (люпин) до 30 г (солома овса). При этом, как следует из внешнего вида (рис. 5–8), большая часть остающегося в почве Сорг продолжает находиться в составе крупных неразложившихся фрагментов РО.
Распределение потерь N орг по видам РО и срокам отбора достоверно коррелировало с потерей C орг и беззольного веса ( r = 0.93). Но из-за более резкого снижения концентрации (рис. 3) потери N орг (в % от исходного содержания) для большинства видов растительного материала были выше по сравнению с беззольным весом и С орг . Уже к концу 1-го года экспозиции в почве РО потеряли от 63% (солома овса) до более 90% (листья овса и люпин) N орг от его исходного содержания. За 2 года потери достигли 88% у соломы овса и 94% у листьев овса и люпина. Однако в абсолютном выражении и потери N орг , и его остаток в почве были значительно ниже, по сравнению с С орг , из-за значительно более низкой исходной концентрации (рис. 10).
Поскольку концентрация P изменялась незначительно, его потери при разложении РО (табл. 2) были близки потерям общего веса и С орг по величине и достоверно коррелировали в распределении между видами материала и сроками отбора образцов с этими показателями ( r = 0.92 для общего веса и 0.91 для С орг ). Наиболее значительные потери P были свойственны травянистым растениям и листьям овса (быстроразлагающимся видам с высоким исходным содержанием), а самые низкие – соломе овса (сочетание низкой скорости разложения и низкой исходной концентрации).
Несмотря на то, что S, как и фосфор, входит в состав органического вещества, ее потери по величине и динамике не согласовались с потерями беззольного веса, Cорг и P (табл. 2). Как и для концентрации, для содержания S в образцах РО было характерно не только снижение, но и незначительное увеличение в отдельные периоды их трансформации. Однако к концу 2-го года экспозиции во всех РО, кроме соломы овса, содержание S в образцах резко снизилось на 85% у метелок овса и на 98% у листьев овса относительно исходных значений. Интенсивный вынос S из окультурен- ного подзола (до 20 г/м2 в год) подтверждается лизиметрическими исследованиями (Переверзев, 1993).
Из-за высокой подвижности и отсутствия механизмов фиксации в данных условиях калий ( K ) наиболее интенсивно выносится из растительного материала в процессе разложения в почве. С учетом потери общего веса уже после 8 месяцев экспозиции образцы потеряли от 82 (солома овса) до 97% (листья овса, люпин, фацелия, вика) K от его исходного содержания. За два года экспозиции все растительные образцы потеряли практически весь K.
Кальций (Ca) и магний (Mg) являются элементами, для которых характерны наиболее значительные различия между видами РО и стадиями разложения по направленности и степени изменения содержания в процессе трансформации. По сравнению с исходным, содержание Ca в листьях овса, в фацелии и вике к концу 1-го года экспозиции в почве снизилось на 70–90%, в соломе овса оно, наоборот, повысилось в 4 раза. Содержание Mg в РО также могло не только снизиться (например, в листьях овса оно снизилось на 99% к концу 2-го года экспозиции), но и увеличиться (например, в соломе овса оно увеличилось в 5 раз к концу 2-го года). Нерегулярные изменения содержания Ca и Mg по мере трансформации РО в почве с превышением исходного уровня свидетельствуют не только о наличии механизма их обратимой фиксации трансформированными остатками (предположительно, карбоксильными группами вновь образуемого гумуса), но и о дополнительном их поступлении из окружающей почвы. Таким источником могут быть мигрирующие вниз по профилю растворы с высоким содержанием Ca и Mg (Переверзев, 1993).
Из-за повышения концентрации Mn (рис. 9) разложение РО не всегда сопровождалось снижением его содержания в образцах (табл. 2). Возрастающие со временем потери Mn были свойственны только для люпина с его очень высокой исходной концентрацией. В листьях овса и вике, характеризующихся низкой исходной концентрацией Mn, его содержание в течение всего эксперимента было значительно (на 600% в листьях овса и на 500% в растениях вики после первых 8 месяцев экспозиции) выше исходного. Более высокое содержание Mn, по сравнению с исходным, предполагает его дополнительное поступление из почвы, а свойственные боль- шинству видов РО нерегулярные смены накопления/снижения свидетельствуют об обратимом механизме его фиксации в разлагающихся РО. Сложность поведения Mn после высвобождения из РО обусловлена не только изменением его подвижности за счет резких изменений окислительно-восстановительных условий (восстановительные в растениях и окислительные в почве), но также и активной ролью микроорганизмов и органического вещества в процессах его трансформации в почве (Водяницкий, 2005).
Изменения содержания Si и Al в процессе разложения РО не имели регулярного характера (табл. 2). Так же, как и для Mn, в процессе трансформации оно могло увеличиваться (иногда с превышением исходного) или снижаться.
Для Fe , с более низкой исходной концентрацией в растениях и значительно большим контрастом с окружающей почвой, было характерно значительное (до 1 500% выше исходного в метелках овса к концу 2-го года экспозиции) повышение содержания со временем во всех видах РО, кроме люпина. В люпине накопленное в первые 8 месяцев экспозиции Fe было вымыто из образцов в процессе дальнейшего разложения РО.
Высвобождение элементов в первый вегетационный период. Как показали результаты, при осеннем внесении сидератов количество высвобождаемых при их разложении питательных элементов в первый вегетационный период после их внесения в почву зависело не только от исходной концентрации в растениях, но и от их поведения в первые 8 месяцев экспозиции РО в почве. В большинстве случаев, за четыре летних месяца первого вегетационного периода высвобождается лишь небольшая доля (табл. 2, разница между II и I отбором) от потребности растений в элементах для формирования нового урожая. Из-за значительной потери в предшествующий осенне-зимний период летом высвободилось только от 0.5 (люпин) до 15% (солома овса) от потребности растений в K. Высвобождающийся летом P мог обеспечить от 11 (вика) до 30% (метелки овса) от потребности сидератов, N – от 12 (фацелия) до 36% (листья овса). Кальций высвобождался летом только при разложении листьев овса и люпина. Трансформированные остатки других видов РО, наоборот, его накапливали. Однако, как обсуж- далось выше, Ca может находиться в них в обменной, т. е. доступной для растений форме.
Только в единичных случаях высвобождающегося летом количества элемента было достаточно для формирования нового урожая того же вида растений: например, Ca – для люпина и Mn – для листьев овса и вики. Причем это было обусловлено способностью продуктов трансформации этих видов РО связывать высвобождающиеся при разложении и/или мигрирующие с почвенными растворами элементы в течение предыдущих 8 месяцев.
Вместе с тем сидераты с высоким исходным содержанием элемента (табл. 1), и/или чьи продукты трансформации способны накапливать дополнительные его количества в предшествующий холодный период (табл. 2), могут существенно снизить дефицит или даже полностью обеспечить потребности для формирования нового урожая видов с более низкими потребностями. Например, люпин может существенно снизить дефицит N для других видов сидератов, полностью обеспечить потребности в Ca для соломы овса, в Mg – для всех частей овса, в Mn – для всех других видов растений.
Однако на основе эксперимента по разложению можно увидеть только некоторые моменты “работы” сидератов в качестве источников минерального питания для растений. Для более полной картины необходимы полевые эксперименты с внесением сидератов и контролем за уровнями доступных элементов в почвах.
ЗАКЛЮЧЕНИЕ
Как показали результаты исследования, при специфическом сочетании холодного гумидного климата, легкого гранулометрического состава почв и аэробных условий, процессы разложения сидератов в агрогенно преобразованных подзолах Субарктики протекали с высокой интенсивностью, сопоставимой с таковой в южной черноземной зоне: после года экспозиции в почве РО теряли от 47 (солома овса) до 75% (люпин) своего исходного беззольного веса. После 2 лет потери веса достигли 68% у соломы овса и 90% у люпина.
Виды сидератов по потерям беззольного веса после первых 8 месяцев экспозиции отличались в 2 раза: от 32% у соломы овса до ≈60% у фацелии. Межвидовые различия со временем сглаживались и по кумулятивным потерям за 2 года варьировали от 68% у соломы овса до около 90% у люпина, т. е. отличались в 1.3 раза.
Распределение потерь С орг ( r = 0.99) и N орг ( r = 0.93) между видами растительного материала и по срокам отбора достоверно коррелировало с потерями беззольного веса. Однако потери С орг были существенно ниже, по сравнению с потерей веса, из-за увеличения концентрации С орг в РО в процессе их разложения, особенно в более поздние сроки экспозиции. За 2 года потери С орг варьировали от 65% (солома овса) до 87% (люпин). Наоборот, потери N орг были выше потерь веса и углерода из-за существенно снижения концентрации, особенно после первого года разложения. За 2 года потери N орг составили 83% у фацелии и 94% у люпина.
Снижение концентрации N орг и обеднение органического вещества РО азотом в процессе их трансформации в почве может быть обусловлено более быстрым разложением белков с высоким содержанием азота, а повышение концентрации С орг – увеличением со временем доли трудно разлагаемых компонентов – лигнина и гуминовых кислот – с высоким содержанием углерода.
Как показали расчеты, несмотря на высокую интенсивность разложения, из каждых выращенных летом и внесенных в почву осенью 200 г воздушно-сухих РО сидератов (средняя урожайность с 1 м2), к началу следующего лета в почве может оставаться от 35 г (фацелия и вика) до 60 г (солома овса) С орг и от 2 г (фацелия) до 5 г (солома овса) N орг . То есть применение сидератов может иметь значительный положительный эффект для баланса C и N в субарктическом агроценозе. При этом большая часть оставшегося в почве С орг входит в состав крупных неразложившихся фрагментов РО.
Поведение зольных элементов в процессе трансформации РО зависело не только от свойств элемента, но и вида растительного материала. Потери K превысили 90% уже после первых 8 месяцев, S и P – достигли 80–98% у быстроразлагающихся видов РО (листья овса, люпин, фацелия и вика) к концу 2-го года экспозиции. В поведении Ca и Mg при разложении некоторых видов РО были выявлены нерегулярные изменения их содержания с периодическими превышениями исходного уровня. Дополнительным источником этих элементов могут быть фильтрующиеся по профилю растворы, а возможным механизмом фиксации – обменное поглощение вновь образуемым гумусом.
Благодаря значительным межвидовым различиям по биохимическому и химическому составу сидератов, по интенсивности и динамике разложения, а также по поведению элементов в процессе трансформации РО в почве, применение сидератов предоставляет большие возможности для управления не только плодородием почв, но и круговоротом элементов в агроценозах за счет подбора культур.
Список литературы Разложение сидератов и высвобождение питательных элементов в условиях субарктики, Кольский полуостров
- Александрова, Л.Н. Органическое вещество почвы и процессы его трансформации. Л.: Наука, 1980. 288 с.
- Водяницкий Ю.Н. Оксиды марганца в почвах. М.: ГНУ Почвенный институт им. В.В. Докучаева РАСХН, 2005. 95 с.
- Гришина Л.А. Гумусообразование и гумусное состояние почв. М.: МГУ, 1986. 244 с.
- Дедов А.А., Дедов А.В., Несмеянова М.А. Динамика разложения растительных остатков в черноземе типичном и продуктивность культур севооборота // Агрохимия. 2016. № 6. С. 3-8.
- Довбан К.И. Зеленое удобрение в современном земледелии: вопросы теории и практики. Минск: Белорусская наука, 2009. 404 с.
- Елсаков Г.В., Горелова А.П., Миронова Р.А. Роль удобрений при возделывании лекарственных культур в Заполярье // Агрохимия. 2001. № 2. С. 40-45.
- Ильин В.Б. Элементарный химический состав растений. Новосибирск: Наука, 1985. 130 с.
- Кашулина Г.М., Коробейникова Н.М., Вирачева Л.Л., Чуева Н.В. Агрохимическая характеристика почв коллекционного питомника интродуцированных лекарственных травянистых растений Полярно-альпийского ботанического сада // Агрохимия. 2021. № 9. C. 13-21. https://doi.org/10.31857/S0002188121090076.
- Кузнецова Т.В., Ходжаева А.К., Семенова Н.А., Иванникова Л.А., Семенов В.М. Минерализационно-иммобилизационная оборачиваемость азота в почве при разной обеспеченности разлагаемым органическим веществом // Агрохимия. 2006. № 6. С. 5-12.
- Ларионова А.А., Мальцева А.Н., Лопес де Гереню В.О., Квиткина А.К., Быховец C.С., Золотарева Б.Н., Кудеяров В.Н. Влияние температуры и влажности на минерализацию и гумификацию лиственного опада в модельном инкубационном эксперименте // Почвоведение. 2017. № 4. С. 438-448.
- Ловчиков А.П., Ловчиков В.П., Поздеев Е.А. Биологизация земледелия в ресурсосберегающих технологиях возделывания зерновых культур // Международный научно-исследовательский журнал. 2016. № 1(43). https://doi.org/10.18454/IRJ.2016.43.094.
- Лошаков В.Г. Зеленое удобрение как фактор повышения плодородия почвы, биологизации и экологизации земледелия // Плодородие. 2018. № 2. С. 26-29.
- Мальцева А.Н., Золотарева Б.Н., Пинский Д.Л. Трансформация растительных остатков кукурузы в суглинистом и песчаном субстратах // Почвоведение. 2013. № 10. С. 1239-1252.
- Манаков К.Н. Водная миграция минеральных элементов в северо таежных ландшафтах Кольского полуострова // Почвенные режимы на Полярном Севере. Л.: Наука, 1969. C. 104-117.
- Переверзев В.Н. Культурное почвообразование на Крайнем Севере. Апатиты: Изд-во Кольского научного центра РАН, 1993. 156 с.
- Переверзев В.Н. Биохимия гумуса и азота почв Кольского полуострова. Л.: Наука, 1987. 303 с.
- Переверзев В.Н. Лесные почвы Кольского полуострова. М.: Наука, 2004. 232 с.
- Поповцева А.А. Методическое руководство по ускоренному анализу золы растений. Сыктывкар: Коми филиал АН СССР. 1974. 84 с.
- Cеменов В.М., Лебедева, Т.Н., Зинякова Н.Б., Хромычкина Д.П., Cоколов Д.А., Лопес де Гереню В.О., Кравченко И.К., Х. Ли, Семенов М.В. Зависимость разложения органического вещества почвы и растительных остатков от температуры и влажности в длительных инкубационных экспериментах // Почвоведение. 2022. № 7. С. 860-875. https://doi.org/10.31857/S0032180X22070085.
- Семко А.П. Агротермический режим почв лесной зоны Кольского полуострова. Апатиты: Изд-во Кольского филиала АН СССР, 1982. 142 с.
- Смирнова О.Г., Кочетов А.В. Клеточная стенка растений и механизмы устойчивости к патогенам // Вавиловский журнал генетики и селекции. 2015. № 19(6). С. 715-723. https://doi.org/10.18699/VJ15.109.
- Столбовой В.С. Влияние потепления климата на баланс углерода в лесных почвах России // Бюллетень Почвенного института имени В.В. Докучаева. 2022. Вып. 111. С. 5-29. https://doi.org/10.19047/0136-1694-2022-111-5-29.
- Шлегель Г. Общая микробиология. М.: Мир, 1987. 566 с. URL: https://hlebspb.ru/uf/editor/files/Ne.
- Ушакова Г.И. Скорость высвобождения химических элементов из опада и подстилки в двух типах еловых лесов Кольского полуострова // Почвоведение и агрохимия в Мурманской области. Апатиты: Изд-во Кольского филиала АН СССР, 1983. С. 16-27.
- Чуева Н.В., Кашулина Г.М., Коробейникова Н.М. Агрохимические свойства почв коллекционных питомников Полярно-альпийского ботанического сада // Труды Кольского научного центра РАН. Серия Прикладная экология Севера. 2021. Вып. 9. С. 287-291. https://doi.org/10.37614/2307-5252.2021.6.12.9.043.
- Cornwell W.K., Cornelissen J.H.C., Amatangelo K., Dorrepaal E., Eviner V.T., Godoy O., Hobbie S.E., Hoorens B., Kurokawa H., Pérez-Harguindeguy N., Quested H.M., Santiago L.S., Wardle D.A., Wright I.J., Aerts R., Allison S.D., van Bodegom P., Brovkin V., Chatain A., Callaghan T.V., Díaz S., Garnier E., Gurvich D.E., Kazakou E., Klein J.A., Read J., Reich P.B., Soudzilovskaia N.A., Vaieretti M.V., Westoby M. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide // Ecology Lett. 2008. Vol. 11. P. 1065-1071. https://doi.org/10.1111/j.1461-0248.2008.01219.
- Green manure/cover crops and crop rotation in conservation agriculture on small farms // Integrated Crop management. Rome. 2011. 97 p.
- Kabata-Pendias A., Mukherjee A.B. Trace Elements from Soil to Human. Springer, Berlin Heidelberg, New York, 2007. 550 p.
- Markert B. Presence and significance of naturally occurring chemical elements of the periodic system in the plant organism and consequences for future investigations on inorganic environmental chemistry in ecosystems // Vegetatio. 1992. No. 103. P. 1-30.
- Nelson D.L., Cox M.M. Lehninger. Principles of Biochemistry. Fourth Edition. W.H. Freeman, 2005. 1130 p. URL: https://handoutset.com/wp-content/uploads/2022/03/Lehninger-Principles-of-Biochemistry-Fourth-Edition-by-David-L.-Nelson-Michael-M.-Cox.pdf.
- Prescott C.E. Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? // Biogeochemistry. 2010. Vol. 101. P. 133-149. https://doi.org/10.1007/s10533-010-9439-0.
- Zhang D., Hui D., Luo Y., Zhou G. Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors // J. Plant Ecology. 2008. Vol. 1. No. 2. P. 85-93. https://doi.org/10.1093/jpe/rtn002.
