Размерно-возрастные различия биоэкологии Mesobuthus eupeus (C.L. Koch, 1839) (Arachnida, Scorpiones, Buthidae) в Азербайджане
 (Arachnida, Scorpiones, Buthidae) в Азербайджане")
Автор: Новрузов Низами Энвер Оглы
Журнал: Вестник Нижневартовского государственного университета @vestnik-nvsu
Рубрика: Экология животных
Статья в выпуске: 2, 2021 года.
Бесплатный доступ
Исследовано биотопическое распределение, размерно-возрастная структура популяции, ночной ритм активности и питание пестрого скорпиона (Mesobuthus eupeus). Исследования проводились на левобережье р. Пирсагат (Ширванская равнина, восточный Азербайджан). Все обнаруженные особи скорпионов по длине метасомы были условно распределены на 5 размерно-возрастных групп. Биотопическое распределение пестрого скорпиона носило выраженный агрегированный характер с образованием локальных скоплений (поселений), в которых было выделено 5 типов соотношений размерно-возрастных групп. Установлено, что особи I и II групп имеют унимодальный характер ритма ночной активности, III и IV групп - бимодальный, а V группы - тримодальный. С увеличением размера скорпионов широта спектра питания увеличивается, а интенсивность питания снижается. Индекс элективности в I и II группах имел отрицательные значения, а с III по V группаы по некоторым таксонам жертв - положительные значения. Ведущую роль в питании особей всех размерно-возрастных групп играют представители отрядов Isopoda (Crustacea), Aranei (Arachnida), Hymenoptera и Lepidoptera (Insecta). С увеличением размера скорпионов доля представителей отрядов Hymenoptera, Blattodea и Colembola в их питании снижается. Представители отрядов Coleoptera и Orthoptera отмечены в питании скорпионов III-V групп. Представители отрядов Pulmonata (Gastropoda), Lumbricidae (Oligochaeta), Scolopendromorpha (Chilopoda), Solifugae (Arachnida) присутствуют в питании только у особей IV и V групп.
Пестрый скорпион, размерно-возрастные группы, биотопическое распределение, ритм активности, спектр питания
Короткий адрес: https://sciup.org/14120890
IDR: 14120890 | УДК: 595.461 | DOI: 10.36906/2311-4444/21-2/09
Size-age differences in the bioecology of Mesobuthus eupeus (C.L. Koch, 1839) (Arachnida, Scorpiones, Buthidae) in Azerbaijan
The biotopical distribution, size and age structure of the population, nocturnal activity rhythm and feeding of the mottled scorpion (Mesobuthus eupeus) were studied. The research was conducted on the left bank of the Pirsagat river (Shirvan plain, Istern Azerbaijan). All discovered individuals of scorpions were divided into 5 size and age groups by the length of the metasome. The biotopical distribution of the mottled scorpion had a pronounced aggregated character with the formation of local clusters (settlements) in which 5 types of ratios of size and age groups were identified. It was found that individuals of II groups and I have unimodal, III and IV groups - bimodal, and V group - trimodal character of the rhythm of nocturnal activity. As the size of scorpions increases, the breadth of the nutrition spectrum increases, and the intensity of nutrition decreases. The index of electivity in II groups and I had negative values, and from III to V groups for some taxa of preys - positive values. Representatives of the orders Isopoda (Crustacea), Aranei (Arachnida), Hymenoptera and Lepidoptera (Insecta) play a leading role in the nutrition of individuals of all size- age groups. As the size of scorpions increases, the proportion of members of the orders Hymenoptera, Blattodea and Collembola in their diet decreases. Representatives of the orders Coleoptera and Orthoptera are noted in the diet of scorpions from group III to V groups. Representatives of the orders Pulmonata (Gastropoda), Lumbricidae (Oligochaeta), Scolopendromorpha (Chilopoda), and Solifugae (Arachnida) are present in the diet only in individuals of IV and V groups.
Текст научной статьи Размерно-возрастные различия биоэкологии Mesobuthus eupeus (C.L. Koch, 1839) (Arachnida, Scorpiones, Buthidae) в Азербайджане
К настоящему времени систематика, морфология, физиология, состав и свойства яда пестрого скорпиона очень подробно исследованы. Однако биология и экология этого вида на всем протяжении его ареала изучена недостаточно. Отсутствуют данные о возрастной структуре популяции, принципах биотопического распределения разных возрастных групп, характере их ночного ритма активности, интенсивности питания и таксономическом спектре поедаемых жертв в естественных условиях. Это препятствует составлению объективной картины участия вида в пищевых и топических связях с другими беспозвоночными в биоценозе. Цель данной работы – изучение биотопического распределения, ночного ритма активности и питания разновозрастных групп пестрого скорпиона в естественных условиях.
Материалы и методы Изучение биоэкологии пестрого скорпиона осуществлялось в общем объеме плановых исследований таксономической структуры и экологии сообществ наземных членистоногих, проводимых в 2012–2020 гг. В статье частично использованы данные разных лет, но в ее основе лежат материалы полевых исследований 2019 г. на территории левобережья р. Пирсагат (Ширванская равнина, восточный Азербайджан).
Возраст скорпионов устанавливался условно по длине метасомы [13], измерения которой проводилось с помощью прозрачной трубки и цифрового штангенциркуля с точностью до 0,1 мм. За период исследований сняты промеры с 376 экз. скорпионов, которых затем распределили на 5 размерно-возрастных групп (табл. 1).
Таблица 1
Длина метасомы исследованных экземпляров M. eupeus разных размерно-возрастных групп (n=376)
|
Размерная группа |
Предположительная стадия онтогенеза |
N |
Длина метасомы, мм |
|
|
Lim |
M±SD |
|||
|
I |
1-2-я линька |
133 |
6,11–8,08 |
7,00±0,18 |
|
II |
3-я линька |
57 |
12,06–16,87 |
14,39±0,64 |
|
III |
4-я линька |
66 |
17,35–23,87 |
20,14±0,97 |
|
IV |
5-я линька |
81 |
24,18–27,70 |
25,20±0,34 |
|
V |
6-я линька (Imago) |
39 |
29,23–33,29 |
31,18±0,80 |
Примечание. lim – минимальное и максимальное значение; M±SD – среднее значение и стандартное отклонение
Размерно-возрастная структура и распределение особей в поселениях скорпионов были изучены методом учетных площадок [22]. Для этого был выбран сравнительно малопосещаемый и незначительно антропогенно трансформированный участок территории площадью около 20 га, сочетающий полупустынный и сухостепной ландшафты. На схеме участка отмечались места обнаружения скорпионов, определялись размерно-возрастные группы, затем наложением масштабной сетки со стороной квадрата 5 м устанавливалась площадь участка, преимущественная позиция и состав поселения. Для определения характера пространственного распределения объектов использовался индекс агрегированности [7]: 1 Д = S/4M, где S - дисперсия, M - среднее арифметическое. Агрегированному распределению особей соответствовали значения индекса I A > 1.
Для определения степени дискретности возрастных групп в поселении расчитывался индекс дискретности по формуле: 1 D = 1 — М/С, где M - количество разобщенных возрастных групп; C -общее количество групп.
Оценка размерно-возрастного разнообразия проводилась с использованием показателя возрастной гетерогенности, вычисляемого по формуле: V= 1/£Р2, где V - показатель возрастной гетерогенности, P i – доля особей i-той возрастной группы. Высокое разнообразие свидетельствовало о стабильности популяции, т.к. различные стадии жизненного цикла обладают неодинаковой устойчивостью к экологическим факторам [3].
Площадь территории, занимаемой каждым поселением, условно делилась на 9 приблизительно равных частей (квадратов). На плане участка отмечалось расположение поселения и «квадрат» обнаружения (позиция) размерной группы (рис. 1). Исследуемая территория имела пространственную неоднородность растительного покрытия и характера микрорельефа поверхности. Неоднородность поверхности почвы биотопа оценивалась по доле (%) травянисто-кустарникового покрытия и перепадам микрорельефа [10]. Особенности распределения микросредовых условий на участке биотопов в местах скоплений скорпионов отражены на фоновых диаграммах уровней (рис. 2). Для построения фоновых диаграмм микрорельефа, отражающей перепады высот местности, применена условная шкала измерения: (-1) – низина, (0) – равнина, (1) – возвышение.
Зависимость распределения и численности скорпионов от микросредовых параметров устанавливалась при помощи коэффициента линейной корреляции Пирсона ®. Динамика ритма ночной активности устанавливалась путем непрерывных ночных наблюдений за объектами в местах их скоплений с предварительным выяснением их численности, возрастной структуры и мечением укрытий. Наблюдения велись после 18.00 ч вечера и до 8.00 ч утра. Общее количество наблюдаемых особей составило 103 экз. Всего зафиксировано 408 ночных выходов скорпионов из укрытий на поверхность, различающихся по своей продолжительности.
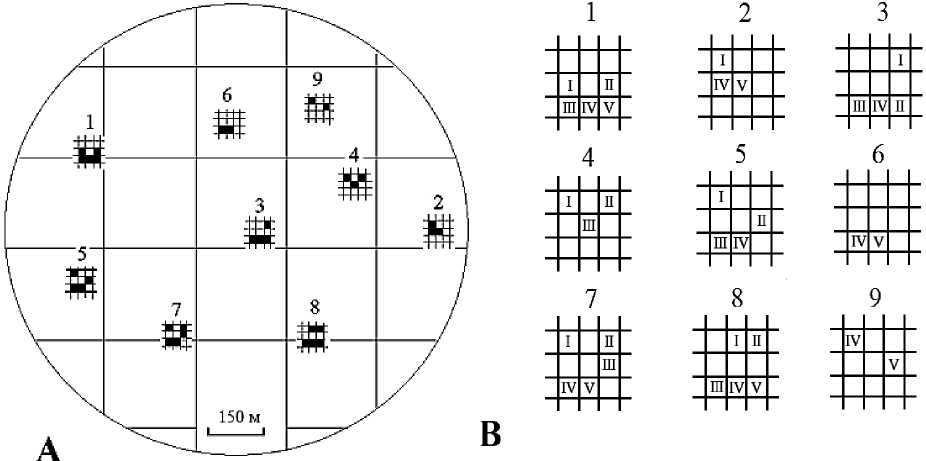
Рис. 1. Расположение поселений пестрого скорпиона на план-схеме участка (А). Позиции размерновозрастных групп в пространствах поселений (В): 1-9 – поселения; I–V – размерно-возрастные группы
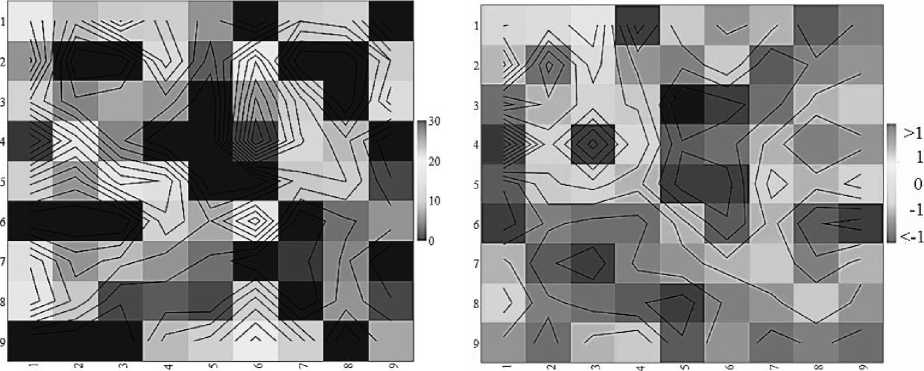
Растительное покрытие Микрорельеф
Рис. 2. Распределение микросредовых условий в местах скоплений скорпионов
Питание M. eupeus изучалось во все сезоны активности (с мая по сентябрь). Фиксировались все случаи обнаружения скорпионов с добычей на хелицерах на поверхности и в укрытиях. Дополнительно проводился сбор и идентификация хитиновых фрагментов их жертв, обнаруженных в укрытиях хищников. Проанализированы хитиновые остатки 840 экз. членистоногих, пригодных для идентификации (преимущественно насекомых). Объекты в основном определялись до отряда и семейства. Проведены визуальные наблюдения за 164 особями скорпионов в процессе охоты и поедания добычи. Для выяснения интенсивности питания объекты исследования взвешивались до и после ночной активности на портативных электронных весах с точностью до 0,01 г. Всего проведено 328 взвешиваний.
Определялся таксономический состав и частота встречаемости беспозвоночных в биотопах с использованием стандартных энтомологических методов учета: маршрутного, ручного сбора, биоценометрии [6; 15]. Визуальный учет на пеших маршрутах использовали для учета отдельных групп (прямокрылые, равнокрылые, сетчатокрылые). Протяженность маршрутов – 200 м, ширина маршрутной ленты – 2 м. Ручной сбор проводился на площадках 25 м2 с поверхности почвы и в укрытиях. Биоценометрия проводилась следующим образом: исследуемый участок поверхности

накрывался деревянной рамкой площадью 1 м2 со съемным мешком из плотной ткани, стянутым в верхней части. Через горловину мешка проводился сбор всех обнаруженных беспозвоночных. Ловушки (пластиковые стаканы объемом 250 мл) устанавливались в количестве 20 шт на 100 м (по одной на каждые 5 м) с экспозицией 5 дней. Объекты определялись в основном до отрядов и семейств, реже до рода и вида.
Значимость тех или иных представителей фауны беспозвоночных в питании скорпионов устанавливалась вычислением индекса элективности [9]: 1Е = (г — р)/(г + р), где r - процентное значение объекта в составе пищи; p – процентное содержание объекта в биотопе. Первичные данные обработаны методами вариационной статистики в программе Microsoft Excel 2010. Построение фоновых диаграм с линиями уровней распределения и кластерный анализ проведены в программе PAST 3.26.
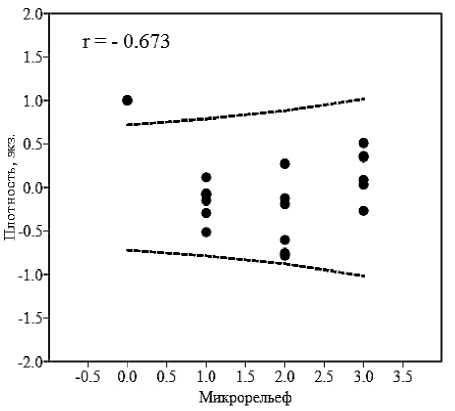
Результаты. На выбранной для исследований незначительно антропогенно трансформированной и не испытывающей пастбищной нагрузки части территории пестрый скорпион встречался на сероземных участках с ксерофитной, сухостепной, степной и псаммофитной растительностью. Скорпионы отмечались преимущественно агрегировано, образуя разные по численности и размерно-возрастному составу скопления (поселения) – 21–68 экз. (40,67±5,73), занимающие небольшие по площади участки пространства поверхности почвы – 29–85 м2 (45,07±2,80). Агрегированность их распределения (I A ) варьировала от псевдоагрегированной (1,03–1,09) до агрегированной (1,21–1,63; M=1,35±0,10). Существенной зависимости плотности населения скорпионов от растительного покрытия и микрорельефа биотопов не выявлено (рис. 3).
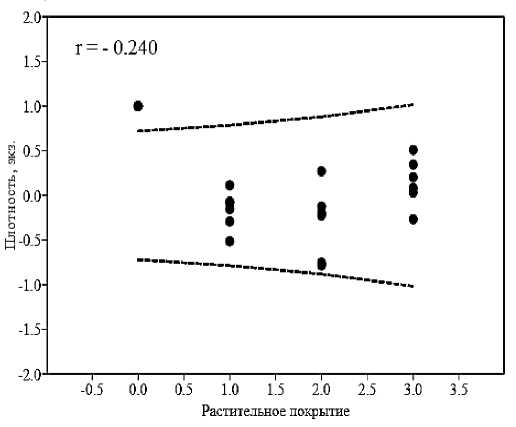
Рис. 3. Корреляция плотности населения скорпионов с микросредовыми условиями биотопа
На обследованном участке было отмечено 9 скоплений (поселений) пестрого скорпиона, дистанцированных друг от друга расстоянием не менее 130 м. Всего за время исследований отмечено 376 экз. скорпионов пяти размерно-возрастных групп (I–V). Численность размерных групп в каждом из обнаруженных поселений представлена в таблице 2, а их распределение на рисунке 4.
Таблица 2

Численность и соотношение возрастных групп M. eupeus
|
Поселение |
Общая численность |
Численность M. eupeus разных возрастных групп (абс./отн. %) |
Показатель возрастной гетерогенности |
||||
|
I |
II |
III |
IV |
V |
|||
|
1 |
68 |
19/27,9 |
9/13,2 |
22/32,3 |
11/16,1 |
7/10,2 |
5,74 |
|
2 |
44 |
25/56,8 |
– |
– |
15/34,0 |
4/9,1 |
4,89 |
|
3 |
34 |
13/38,2 |
6/17,6 |
8/23,5 |
7/20,5 |
– |
4,07 |
|
4 |
54 |
30/55,5 |
18/33,3 |
6/11,1 |
– |
– |
4,21 |
|
5 |
53 |
12/22,6 |
7/13,2 |
18/33,9 |
16/30,1 |
– |
4,30 |
|
6 |
21 |
– |
– |
– |
13/61,9 |
8/38,1 |
1,60 |
|
7 |
44 |
16/36,3 |
7/15,9 |
9/20,4 |
7/15,9 |
5/11,3 |
5,57 |
|
8 |
37 |
18/48,6 |
10/27,0 |
3/8,1 |
3/8,1 |
3/8,1 |
8,31 |
|
9 |
21 |
– |
– |
– |
9/42,8 |
12/57,1 |
1,51 |
|
Всего |
376 |
133 |
57 |
66 |
81 |
39 |
40,2 |
Для оценки сходства и различия структуры поселений скорпионов на сравниваемых участках территории был использован кластерный анализ сходства Брея-Кертиса. Полученные результаты представлены в виде дендрограммы (рис. 5).
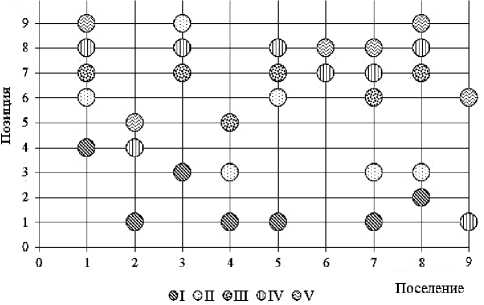
Рис. 4. Распределение размерно-возрастных групп в поселениях M. eupeus
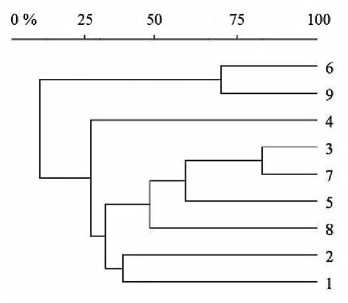
Рис. 5. Дендрограмма сходства и различия структуры сравниваемых поселений (1-9) пестрого скорпиона
По трем критериям оценки (размерно-возрастной состав, численное доминирование групп и степень их разобщенности (индекс I A ) выделено 5 типов поселений пестрого скорпиона: 1) присутствие всех 5 групп с доминированием I, II (48,6 и 27,0%) или I, III (27,9 и 32,3%), индекс разобщенности – 0,20–0,60; 2) присутствие I–IV групп с доминированием I, III (23,5 и 38,2%) или III, IV (30,1 и 33,9%), индекс разобщенности – 0,25; 3) присутствие I, IV, V групп с доминированием I, IV (56,8 и 34,0%), индекс разобщенности – 0,33; 4) присутствие I, II, III групп с доминированием I, II (55,5 и 33,3%), индекс разобщенности – 0,33; 5) присутствие IV, V групп с доминированием IV (61,9%) или V (57,1%), индекс разобщенности 0–0,50.
Полученная дендрограмма демонстрирует наибольшее сходство поселений 3 и 7 (81,2%), наименьшее – поселений 1 и 2 (38,4%).
В ночное время скорпионы ведут активный образ жизни на поверхности, совершают дальние прогулки, охотятся на других беспозвоночных, занимаются поиском брачных партнеров в период размножения. Исключение могут составлять животные, проходящие процесс линьки и самки, вынашивающие потомство, вынужденные на протяжении нескольких недель прятаться в облюбованных заранее убежищах. В мае-июне ввиду отсутствия пополнения (сравнивались только II– V возрастные группы) сбор данных осуществлялся раз в 5 дней и ограничивался фиксированием количества активных особей на поверхности (рис. 6). Среднесуточная температура в мае-июне на поверхности почвы менялась от 16 до 49°С, в укрытиях от 18 до 21°С. Пик максимума среднесуточной температуры на поверхности почвы в мае приходился на 15–16 часов, минимума – на 02 ч, а в июне поверхность почвы максимально прогревалась к 13 часам, а минимум температуры приходился на 04 ч. В укрытиях за все время наблюдений температура достигалась своего максимума только к 17 часам, а минимума к 06 часам. Ввиду этого продолжительность ночной активности проходила по сокращенному сценарию: около 3 ч – у скорпионов II, III групп и 4 ч – у скорпионов IV, V групп.
Исследование почасовой динамики ночной активности скорпионов было начато с появлением пополнения (2–3 декада июля) и проводилось в августе-сентябре путем непрерывных ночных наблюдений за объектами в местах их скоплений.
Ночная активность M. eupeus по данным на август отмечалась с 19 ч и до 06 ч. Для особей I и II групп отмечен унимодальный (19–23 ч), для III и IV групп бимодальный (20–01ч; 03–07 ч), а для особей V группы – тримодальный (21–23 ч; 01–02 ч; 04–06 ч) характер ночной активности (рис. 7). Продолжительность ночной активности в I и II группах была 3–4 ч, III группе – до 8 ч, IV группе – 6– 7 ч, в V группе – 5–6 ч. По общей продолжительности ночной активности самцы IV и V групп незначительно превосходили самок, но по мобильности и дистанцированности удалений от дневного укрытия значительно (в 2–3 раза) превалировали над ними.
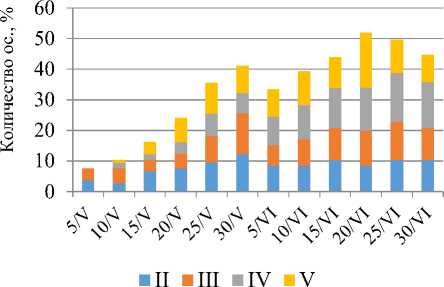
Рис. 6. Ночная активность скорпионов в мае-июне
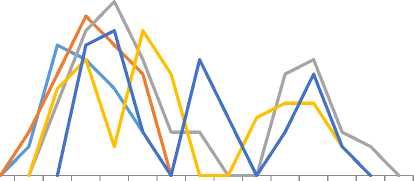
18 19 20 21 22 23 24 1 2 3 4 5 6 7 8
I II III Время, час
Рис. 7. Возрастные различия ритма ночной активности M. eupeus (I-V - размерно-возрастные группы)
Спектр питания M. eupeus в обобщенном виде включал представителей 5 классов, 16 отрядов и 26 семейств. При этом, с увеличением длины тела особей, увеличивалась и широта спектра их питания. Спектр питания I группы представляли 8 отрядов, 12 семейств; II группы – 9 отрядов, 14 семейств; III группы – 13 отрядов, 18 семейств; IV группы – 16 отрядов, 26 семейств; V группы – 15 отрядов (исчезают Hymenoptera), 25 семейств.
По частоте встречаемости объектов в пище общим сходством для всех размерно-возрастных групп было преобладание представителей отрядов Isopoda (15,24±1,71%) и Aranei (11,48±0,55%). С увеличением размеров скорпионов сокращается частота встречаемости в пище представителей Hymenoptera (с 22,2 до 2,7%), Collembola (с 15,3 до 1,8%), Zygentoma (с 11,1 до 5,1%), и возрастает частота встречаемости Isopoda (с 12,8 до 20,5%). Встречаемость Aranei изменяется в небольших пределах (10,2–12,9%).Представители отрядов Opiliones, Orthoptera и Coleoptera начинают встречаться только с III возрастной группы и старше, а отрядов Pulmonata, Lumbricidae, Scolopedromorpha,
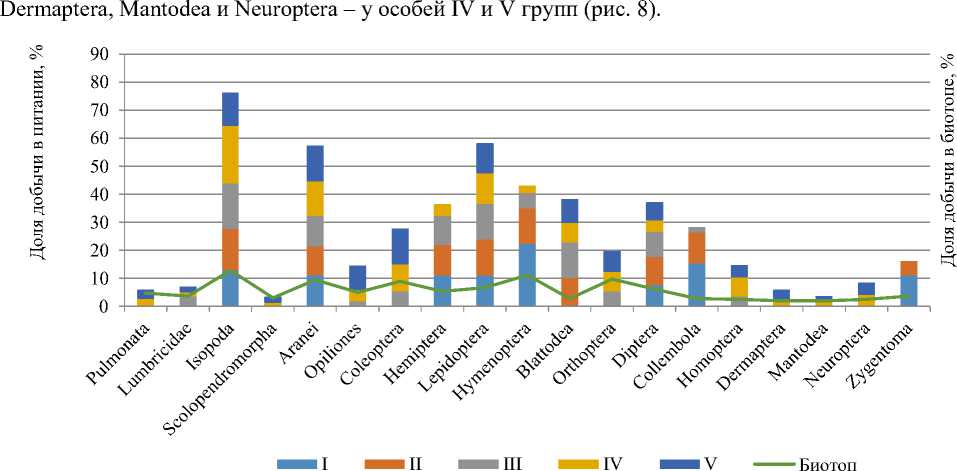
Рис. 8. Состав жертв в пище M. eupeus разных возрастных групп и в биотопе, %
Сравнение интенсивности питания разных размерно-возрастных групп, установленное по массе съеденной пищи, также показало значительные различия. По массе съеденной пищи лидировали скорпионы II и III групп, в среднем потребляя за сутки 39,1 и 47,2 % от собственной массы, скорпионы I группы за сутки потребляли в пищу в среднем 19,7 %, а IV и V групп – 17,4 и 17,1 % (табл. 3).

Таблица 3
Масса тела M. eupeus разных возрастных групп до и после ночной активности (n=164)
|
Размерная группа |
n |
Масса тела, г |
% от массы Тела |
|
|
До охоты |
После охоты |
|||
|
I |
35 |
0,012–0,020/0,015 |
0,019–0,026/0,022 |
19,7 |
|
II |
28 |
0,040–0,074/0,054 |
0,053–0,081/0,065 |
39,1 |
|
III |
40 |
0,240–0,560/0,342 |
0,340–0,710/0,452 |
47,2 |
|
IV |
36 |
0,750–1,203/0,975 |
0,901–1,370/1,121 |
17,4 |
|
V |
25 |
0,950–2,704/1,990 |
1,031–3,160/2,213 |
17,1 |
При сравнении таксономического спектра питания разных размерных групп в разные сезоны активности установлено, что спектр питания определялся как составом и численностью доступных жертв, так и возрастом и отчасти полом скорпиона. Сопоставление состава питания молодых и половозрелых скорпионов выявило значительное изменение таксономического спектра пищи и размеров добычи в разные месяцы. В мае по ширине спектра питания лидировали скорпионы III и IV групп (30,2 и 29,5% соответственно), в июне – IV группы (28,9%), в июле – V группы (31,3%), в августе – II группы (29,1%), а в сентябре – II и IV групп (по 26,3%). Скорпионы I группы обнаруживались в биотопе не ранее середины июля, поэтому сезонность их питания не рассматривалась. Рацион остальных размерно-возрастных групп в основном формировался за счет беспозвоночных, имеющих сходную с ними ритмику сезонной активности (рис. 8).
В летние месяцы трофическая активность скорпионов на поверхности отмечалась преимущественно в сумеречно-ночные часы. В мае и сентябре пищевая активность на поверхности сокращалась по продолжительности и смещалась на вечерне-ночные часы (18–22 ч). По мере увеличения размера скорпионов их пищевой спектр таксономически менялся, прежде всего, за счет потребления более массовых и доступных по размерам видов добычи. Максимальные размеры жертв у взрослых особей не превышали 48 мм, основу же питания при этом составляли относительно мелкие виды, длина которых составляла 15–20 мм.
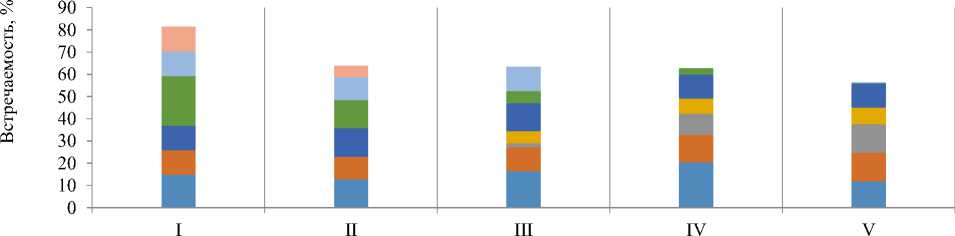
Как уже отмечалось, спектр жертв у скорпионов IV и V групп почти в два раза шире, чем у особей I и II групп. Существенно различается у разных возрастных групп скорпионов удельная доля отдельных отрядов их жертв (рис. 9). Виды семейство. Acrididae (Ortopthera), занимающие одно из доминирующих положений в энтомофауне биотопа (9,7%), почти не входят в рацион взрослых скорпионов (7,5%) несмотря на не слишком крупные размеры (2,5–3 см) и относительную мякость покровов. В то же время Coleoptera отмечены в рационе практически всех взрослых особей (95%). Изменение спектра жертв пестрого скорпиона с увеличением возраста наиболее ярко отражается в увеличении удельной доли и числа видов жесткокрылых – от 1,8% (III группа) и 9,5% (IV группа) до 12,9% у половозрелых особей (V группа).
■ Isopoda ■ Aranei ■ Coleoptera ■ Orthoptera ■ Lepidoptera ■ Hymenoptera ■ Hemiptera ■ Zygentoma
Рис. 9. Изменение спектра питания M. eupeus разных возрастных групп по основным таксонам жертв
В рационее скорпионов I группы доминируют Hymenoptera (22,2%, в основном Formicidae) и Isopoda (14,8%, в основном Armadillididae и Porcellionidae). Реже отмечаются Diptera (7,4%). В рационе скорпионов II группы обнаружено 9 крупных таксонов кормовых объектов. Доминирующее положение занимают Collembola (15,3%). Часто отмечались ракообразные Isopoda (12,8%) и личинки Lepidoptera (12,7%). В рационе данной группы впервые появляются Blattodea (10,2%), представленные в основном личинками Blatta orientalis и Phylodromica sp . Пищевой спектр скорпионов III и IV групп более
Вестник НВГУ. № 2(54) / 2021 ЭКОЛОГИЯ ЖИВОТНЫХ / ANIMAL ECOLOGY разнообразен и представлен 13–16 крупными таксонами кормовых объектов. Наиболее часто встречались Isopoda (16,3 и 20,5% соответственно) и личинки Lepidoptera (12,7 и 10,9% соответственно). В этих возрастных группах уже появляются, хотя не являются доминантами в рационе Coleoptera (до 9,5%) и Orthoptera (5,4 и 6,8% соответственно). Значительную долю в рационе занимают Aranei (10,9 и 12,3%), Blattodea (12,7 и 6,8%). Остальные пищевые объекты в рационе скорпионов составляли менее 4%.
Примечательно, что доминирующую группу жесткокрылых (12,9%), собранных в укрытиях взрослых особей, в основном самок V группы, составляют достаточно крупные виды семейства Tenebrionidae (15–25 мм) с плотными хитиновыми покровами. У половозрелых скорпионов значительно увеличивается, и доля Orthoptera (с 5,4 до 7,5%). У скорпионов III, IV групп они представлены личинками Acrididae, а у половозрелых особей V группы к ним присоединяются Gryllotalpidae – обитатели верхних слоев почвы, выходящие на поверхность исключительно в сумеречно-ночное время. Отметим, что большинство объектов, представленых в питании взрослых скорпионов (V группы), относятся к видам с сумеречной и ночной активностью Armadillididae и Porcellionidae (Isopoda), Gryllotalpidae, Oecanthidae (Orthoptera), Blattidae, Blattelidae (Blattodea). Основу рациона ювенильных особей (I группа) примерно в равной степени составляют виды с дневной и сумеречно-ночной активностью – Porcellionidae, Armadillididae (Isopoda), Miridae (Hemiptera), Formicidae (Hymenoptera). Увеличение доли Lepidoptera (11,1–12,8%) в пище особей M. eupeus I–III групп происходит исключительно за счет личиночных стадий, имаго отмечены только у единичных особей IV и V групп. По мере увеличения размера скорпионов доминирующую позицию теряют представители отрядов Hemiptera (с 11,1 до 4,2%) и Hymenoptera (с 22,2 до 2,7%). Последние в основном за счет Formicidae составляли основу рациона особей I группы (22,2%), а у половозрелых – всего 2,7%. Практически неизменной остается доля Aranei (11,1–12,9%), представленных 4 семействами. Не составляют исключения и сами скорпионы, остатки которых единично (0,6%) отмечались в основном в укрытиях предадультных и адультных самок.
Для выявления избирательной способности скорпионов (индекс элективности) было проведено количественное и качественное сравнение встречаемости беспозвоночных в их пище и в биотопе. По индексу элективности определена положительная избирательность к некоторым группам жертв. Индекс элективности в I и II группах имел отрицательные значения (за исключением Collembola во II группе), а в III, IV и V группах по 4–7 таксонам жертв – положительные значения (табл. 4).
Таблица 4
Значения индекса элективности для таксонов добычи M. eupeus
|
Таксон добычи |
Индекс элективности размерновозрастных групп |
Таксон добычи |
Индекс элективности размерновозрастных групп |
||||||||
|
I |
II |
III |
IV |
V |
I |
II |
III |
IV |
V |
||
|
Lumbricidae |
– |
– |
-0,20 |
-0,50 |
-0,20 |
Diptera |
-0,55 |
-0,27 |
-0,16 |
-0,40 |
-0,07 |
|
Pulmonata |
– |
– |
– |
-0,55 |
-0,40 |
Coleoptera |
– |
– |
-0,25 |
0,16 |
0,41 |
|
Isopoda |
-0,20 |
-0,09 |
0,20 |
0,42 |
0,29 |
Lepidoptera |
-0,33 |
-0,09 |
0,07 |
0,14 |
0,25 |
|
Chilopoda |
– |
– |
– |
-0,60 |
-0,33 |
Blattodea |
– |
-0,20 |
0,07 |
-0,09 |
0,14 |
|
Aranei |
-0,33 |
-0,20 |
0 |
0,20 |
0,33 |
Collembola |
-0,14 |
0,20 |
-0,60 |
- |
- |
|
Opiliones |
– |
– |
-0,60 |
-0,14 |
0,33 |
Mantodea |
– |
– |
– |
-0,20 |
-0,50 |
|
Dermaptera |
– |
– |
– |
-0,33 |
-0,14 |
Zygentoma |
-0,33 |
-0,50 |
– |
– |
– |
|
Orthopterа |
– |
– |
-0,33 |
-0,09 |
0,07 |
Neuroptera |
– |
– |
– |
-0,25 |
-0,11 |
|
Hemiptera |
-0,25 |
-0,11 |
0,09 |
– |
– |
Homoptera |
– |
– |
-0,50 |
-0,09 |
-0,20 |
|
Hymenoptera |
-0,07 |
-0,16 |
-0,40 |
-0,55 |
– |
||||||
Обсуждение. Биотопическое распределение и размерно-возрастная структура популяции. Пестрый скорпион относится к числу типичных обитателей пустынных, полупустынных и степных биотопов. Своей нишей избрал сероземные, песчаные и каменистые полупустынные участки, в которых предпочитает готовые укрытия в виде крупных и средних камней, трещин в почве, нор мышевидных грызунов и т.п. В сочетании с небольшими самостоятельно вырытыми норами все эти укрытия вполне обеспечивают ему защиту от естественных врагов и неблагоприятных климатических факторов в дневное время и в период зимовки [5].
Для M. eupeus не отмечены периодические резкие увеличения численности, чередующиеся с периодами депрессии, характерные у некоторых других паукообразным, например, Solifugae, Aranei [2; 14]. Однако некоторые колебания численности все же отмечены, что предположительно связано с изменением гигротермических условий среды, подъемом и депрессией численности некоторых базисных кормовых объектов в биотопе (Orthoptera, Lepidoptera, Homoptera) [1; 18; 20]. Так, проведение учета численности в разные годы показало изменчивую плотность поселений скорпионов: в 2013 г/ – 30,0±4,34 экз./100 кв.м., в 2014 г/ – 28,33±5,93 экз./100 кв.м., в 2016 г/ – 22,17±5,09 экз./100 кв.м. Разобщенность возрастных групп в послениях, вероятно, связана с онтогенетическими особенностями распределения разновозрастных особей [26]. Так, например, собственными наблюдениями было отмечено, что нимфы пестрого скорпиона вплоть до 2-ой линьки держатся вместе, т. к. в течение некотого времени посре рождеия у них сохраняется «эффект группы» (Allee effect) [21]. Заметное разобщение особей происходит только после 3-ей линьки.
О сроках появления потомства у пестрого скорпиона сведения в литературе разноречивы и охватывают период с мая по сентябрь [11; 12; 14]. Появление новорожденных скорпионов в условиях равнинного Азербайджана происходит с конца мая до середины июля [20; 19]. Ввиду этого изучение размерно-возрастной структуры поселений скорпионов проводилось в августе-сентябре с тем, чтобы во время исследований в материале присутствовали все возрастные категории. Биотопическое распределение пестрого скорпиона имело выраженный агрегированный характер с образованием локальных скоплений (поселений). Выделено 5 типов поселений по соотношениям и степени дискретности разных размерно-возрастных групп. От степени дискретности обитания разных возрастных групп в поселениях, предположительно, может зависеть пространственное распределение вида, т. к. при этом варьирует острота внутривидовой конкуренции в силу различия их ритма активности и пищевых предпочтений [26]. При достаточно изолированном обитании разных возрастных групп (I D > 0), в каждой из них чаще должны выживать особи менее сходные с особями другой группы. Поэтому с увеличением дивергенции между группами ослабляется конкуренция [17].
Теоретически можно предположить и другие типы биотопического распределения вида, для их выделения необходимы дополнительные данные, на сбор которых потребуется больший объем исследований с охватом значительных территорий. Но даже обнаружение 5 указанных типов распределения позволяет предположить участие в механизме формирования структуры поселений скорпионов ряда факторов. Например, таких, как разные типы почвы и микроклиматические условия в укрытиях, неравномерность распределения кормовых объектов в биотопе, прессинг хищников, индивидуальность онтогенетического развития скорпионов и их естественная элиминация.
Суточный ритм активности . Принято считать, что скорпионы совершенно не выносят света и в естественных условиях проводят светлую часть суток в различных укрытиях, появляясь на поверхности только в сумеречно-ночное время. Их ритмическая активность имеет строго 24-часовую периодичность, которая тесно связана с суточной сменой дня и ночи [26]. В литературе по активности пестрого скорпиона в условиях Азербайджана сведения ограничиваются указанием, что они активны исключительно в ночное время и выходят из убежищ после захода солнца [19]. В данных по другим частям ареала их обитания указывается общая продолжительность ночной активности без указания возраста или размеров отмеченных особей [12; 16]. Для получения собственных данных по динамике ночной активности разных возрастных групп проводились непрерывные ночные наблюдения за особями с фиксированием маршрутов их передвижения и основных укрытий. По данным на август-сентябрь, первыми на поверхности (преимущественно возле укрытий) в 19 часов отмечались скорпионы I и II групп. В промежутках между 21–23 ч, 01–02 ч и 04–06 ч встречались взрослые особи. Период их ночной активности независимо от результатов охоты, завершался к 06 часам утра. Скорпионы III группы из дневных укрытий выходили ближе к заходу солнца и в сумерках (20 ч) и отмечались на поверхности в промежутках 20–01 ч и 04–07 часов. Таким образом, к периоду появления на поверхности взрослых скорпионов активность молодых (II группы) была снижена, а представители I группы практически не встречались. По продолжительности ночной активности лидирует III группа (8 ч), что согласуется с их лидерством по интенсивности питания. Несколько уступают ей особи IV и V группы (6–7 и 5–6 ч). Продолжительность ночной активности скорпионов I и II групп не превышала 4 ч.
Сравнение питания размерно-возрастных групп пестрого скорпиона . Исследования разных авторов, проведенные в лабораторных условиях, показывают лишь размерные различия жертв у пестрого скорпиона в зависимости от возраста особей [8; 19]. По таксономическим различиям спектров питания возрастных групп данные отсутствуют. В ходе собственных исследований в их рационе установлено от 36 до 50 видов, в число которых входили преимущественно представители Isopoda, Insecta и Aranei. Все размерно-возрастные группы данного вида объединяет отсутствие специализации к каким-либо определенным видам добычи. В то же время отмечено предпочтение одних групп таксонов членистоногих и избегание других. Наибольшая частота встречаемости Hymenoptera отмечена у скорпионов I группы (22,2%). Так, например, из 16 видов Formicidae, обнаруженных в районе исследования, в рационе скорпионов I–III групп отмечены только три – Tapinoma sp., Messor
sp., Tetramorium sp. Причем при поедании муравьев скорпионы всегда ограничивались головной и гудной частями жертвы, оставляя нетронутым брюшко. Из насекомых в равной степени предпочитались отряды Diptera, Homoptera, Hemiptera, Collembola, Ephemeroptera, личинки Coleoptera, Lepidoptera, Blattodea и Ortoptera. В меньшей степени использовались в пищу имаго Hymenoptera и Coleoptera. Из отряда Aranei можно отметить представители семейств Tetragnathidae, Theridiidae, Clubionidae, Araneidae, Linyphiidae, Lycosidae, Thomisidae и Salticidae.
На предпочтение жертв определенных таксонов указывает, и доля их остатков в укрытиях скорпионов. Так, например, доля остатков Opiliones, Coleoptera, Lepidoptera (larve), Hymenoptera и Isopoda по отношению к другим группам в укрытиях скорпионов превышала долю этих беспозвоночных по отношению к другим группам в биотопе. В укрытиях с большей частотой отмечались остатки Orthoptera, Aranei, Coleoptera, Diptera и Hymenoptera. По коэффициенту элективности определена положительная избирательность к указанным группам жертв у скорпионов старших возрастных групп и отрицательная у младших групп. По интенсивности питания скорпионов в литературе приводятся данные, полученные в условиях лаборатории, и указывающие, что она подвержена изменениям в зависимости от температурных условий, прохождения жизненно важных биологических циклов (линька, размножение, гибернация) [19]. Наиболее благоприятный температурный градиент для пищевой активности M. eupeus, указанный в литературе, составлял 25– 30° С [19]. Большинство скорпионов предпочитают температуры от 18 до 35° C, но для каждого из видов отмечен свой диапазон температур, при котором трофическая активность проявляется чаще всего [25]. По собственным данным, пищевая активность пестрого скорпиона в естественных условиях проявлялась в более широком диапазоне температур – 12–35° С, достигая своего максимума при температурах 26–28º С. Интенсивность питания скорпионов, исследованная в естественных условиях, отличалась у особей разных размерно-возрастных групп с явным лидерством особей II и III групп (39,1 и 47,2%), что предположительно объясняется большей динамикой процессов роста на этих стадиях развития.
Заключение. Исследование битопического распределения, ночного ритма активности и питания 5 размерно-возрастных групп пестрого скорпиона ( Mesobuthus eupeus ) выявило разобщенность возрастных групп в поселениях, различия продолжительности и почасового ритма активности и пищевых предпочтений. Биотопическое распределение пестрого скорпиона имело выраженный агрегированный характер с образованием локальных скоплений (поселений). Выделено 5 типов поселений по соотношениям и степени разобщения разных размерно-возрастных групп. Установлена некоторая асинхронность в продолжительности и динамике ночного ритма активности особей разных размерно-возрастных групп. Особи I и II групп имели унимодальный характер ночной активности, III и IV групп – бимодальный, а V группы – тримодальный. По продолжительности ночной активности лидируют особи III группы. С увеличением размера скорпионов интенсивность питания уменьшается, а широта спектра питания и индекс элективности увеличивается. Индекс элективности в I и II группах имел отрицательные значения, а с III по V группах по некоторым таксонам жертв – положительные значения. Ведущую роль в питании скорпионов играют представители классов Crustacea (отряд Isopoda), Arachnida (отряд Aranei), Insecta (отряды Hymenoptera, Lepidoptera). С увеличением размеров скорпионов доля Hymenoptera, Blattodea и Collembola в их питании снижается. Представители отрядов Coleoptera, Orthoptera встречаются у особей III и IV групп, с длиной метасомы более 27,0 мм. Представители отрядов Lumbricidae (Oligochaeta), Pulmonata (Gastropoda), Chilipoda (Miriapoda), Solifugae (Arachnida) присутствуют в питании только особей IV, V групп.
Автор выражает искреннюю признательность своим коллегам по работе за помощь в определении материала и ценные советы, и замечания, сделанные в процессе подготовки рукописи.
Список литературы Размерно-возрастные различия биоэкологии Mesobuthus eupeus (C.L. Koch, 1839) (Arachnida, Scorpiones, Buthidae) в Азербайджане
- Алиев Х.А., Гусейнов Э.Ф., Топчиева Ш.А., Мусаева А.Н. Влияние экологических факторов на размеры скорпионов // Ekologiya, Fəlsəfə, Mədəniyyət. Elmi məqalələr məcmuəsi, 38-ci buraxılış, Bakı, 2004. C. 154-155.
- Алиев Ш.И. Сольпуги (Arachnida, Solifugae) Азербайджана: Автореф. дис…канд. биол. наук. Баку. 1984.
- Бигон М., Харпер Дж., Таузенд К. Экология. Особи, популяции и сообщества. Т. 1. М.: Мир. 1989.
- Бялыницкий-Бируля А.А. Arachnoidea Arthrogastra Caucasica. Ч. I. Scorpiones. Записки Кавказского музея, Тифлис, A (5). 1917. 253 с.
- Гаджиев А.Т. Скорпионы // Животный мир Азербайджана. Т. 2. Баку: Элм. 1996.
- Гиляров М.С. Методы почвенно-зоологических исследований. М.: Наука. 1975.
- 7.Гиляров М.С. Фауна и экология почвенных беспозвоночных Московской области. М.: Наука. 1983.
- Гусейнов Э.Ф., Алиев Х.А., Топчиева Ш.А., Мусаева Н.Н. Сравнение пищи пауков (Araneae) и скорпионов (Scorpiones) // Экология. Философия. Культура. Вып. 38. Баку. 2004. С. 25-26.
- Ивлев В.С. Экспериментальная экология питания рыб. М.: Пищепромиздат. 1955.
- Кашкаров Д.Н. Среда и сообщество (основы синекологии). М. 1933.
- Кобахидзе Д.Н. К нахождению пестрого скорпиона на обращенных к Черному морю склонах Главного Кавказского хребта // Зоологический журнал. 1960. Т. 39. №10. С. 1573–1574.
- Малинина О.В. Mesobuthus eupeus, образ жизни // Природа. 1953. №10. С. 82–85.
- Мариковский П.И. Материалы по биологии и ядовитости скорпиона Buthus eupeus (C. Koch) // Труды Института зоологии АН Казахской ССР. 1981. T.II. С. 160–166.
- Мариковский П.И. Тарантул и каракурт. Фрунзе. 1956.
- Тихомирова А.Л. Учет напочвенных беспозвоночных // Методы почвенно-зоологических исследований. М.: Наука. 1975. С. 73–85.
- Фет В.Я. К экологии скорпионов (Arachnida, Scorpiones) юго-восточных Каракумов // Энтомологическое обозрение. 1980. LIX. T. 1. С. 223–228.
- Шмальгаузен И.И. Факторы эволюции (теория стабилизирующего отбора). M.: Наука. 1968.
- Юсифов Э.Ф., Мамедов А.А., Новрузов Н.Э., Халилова В.С. Модель динамики численности паукообразных в спектре их межвидовых конкурентных отношений // Математическое моделирование. 2019. Т. 31. №4. С. 131–144.
- Юсубов Э.Б. Скорпионы (Arachnidae, Scorpiones) Азербайджана: Автореф. … канд. биол. наук. Баку. 1984.
- Юсубов Э.Б., Гаджиев А.Т. О численности скорпионов в Азербайджане // Полезные насекомые и их охрана в Азербайджане: Материалы симпозиума. Баку: Элм. 1983. С. 33–34.
- Allee W.C., Bowen E.S. Studies in animal aggregations: mass protection against colloidal silver among goldfishes // Journal of Experimental Zoology. 1932. Vol. 61. №2. P. 185-207. https://doi.org/10.1002/jez.1400610202
- Don B.A. C., Rennolls K. A home range model incorporating biological attraction points // The Journal of Animal Ecology. 1983. P. 69-81. https://doi.org/10.2307/4588
- Fet V. A catalogue of scorpions (Chelicerata: Scorpiones) of the USSR // Rivista del Museo Civico di Scienze Naturali “Enrico Caffi”. 1989. Vol. 13. P. 73-171.
- Fet V., Sissom W.D., Lowe G., Braunwalder M.E. Catalog of the scorpions of the world (1758-1998). New York Entomological Society, 2000.
- Hadley N.F. Adaptational biology of desert scorpions // Journal of Arachnology. 1974. P. 11-23.
- Polis G. A. Seasonal patterns and age-specific variation in the surface activity of a population of desert scorpions in relation to environmental factors // The Journal of Animal Ecology. 1980. P. 1-18. https://doi.org/10.2307/4275
- Teruel R. First record of Mesobuthus eupeus (Koch, 1839) from western Turkey (Scorpiones: Buthidae) // Revista Ibérica de Aracnologia. 2002. Vol. 5. P. 75-76.
