Размещение подроста в микрогруппировках и окнах северных широколиственно-кедровых лесов

Автор: Возмищева А.С., Лонкина Е.С., Крестов П.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 3, 2012 года.
Бесплатный доступ
Рассмотрены особенности влияния верхних ярусов на подрост древесных видов - потенциальных доминантов широколиственно-кедровых лесов. По отношению к состоянию верхнего полога подрост деревьев подразделен на 3 группы: 1) виды, возобновляющиеся преимущественно в окнах; 2) виды, успешно возобновляющиеся и под пологом, и в окнах; 3) виды, возобновляющиеся преимущественно под пологом. Установлено что, распределение и высота подроста зависят от типа микрогруппировки, а также от местоположения внутри окна. Условия возобновления большинства видов в окне оптимальны на расстоянии первых метров от края крон.
Древостой, подрост, полог, пространственное распределение, кедр корейский, теневыносливость
Короткий адрес: https://sciup.org/14082262
IDR: 14082262 | УДК: 630.182.2
Young growth distribution in the microcommunities and openings of the northern broadleaved-korean pine forests
The peculiarities of overwood influence on the young growth of woody species which are the potential dominants of the broadleaved-Korean pine forests are considered. In relation to a shelterwood state the young growth can be divided into three groups: 1) species which are mainly regenerated in the openings, 2) species which are successfully regenerated both under a shelterwood and in the openings, and 3) species which are mainly regenerated under a shelterwood. It is determined that young growth distribution and height depends on the microcommunity type, as well as on the location within the opening. The regeneration conditions of most species in the opening are optimal at the distance of the first meters from the crown edge.
Текст научной статьи Размещение подроста в микрогруппировках и окнах северных широколиственно-кедровых лесов
Введение. Широколиственно-кедровые леса Восточной Азии представляют один из самых сложных по видовому составу и динамическим процессам типов экосистем в широтных пределах 40–50° с.ш. Их древостой сформирован видами с различными экологическими характеристиками и с разной продолжительностью жизненного цикла [16, 20]. Для экосистем характерна большая пространственная неоднородность и выраженная вертикальная сомкнутость.
Успешность возобновления различных по теневыносливости эдификаторов древостоя широколиственно-кедровых лесов зависит от множества факторов биологической и экологической природы. При выпадении из состава первого и второго яруса крупных деревьев в пологе леса образуются окна, способствующие возобновлению светолюбивых видов. Оконная динамика – основной естественный механизм поддержания развивающихся длительное время без катастрофических нарушений листопадных широколиственных и смешанных лесов Европы [1, 9, 13], Азии [19, 22] и Северной Америки [14, 18].
Для прогноза развития сложных лесных сообществ актуальны исследования внутриценотической организации лесного сообщества.
Цель работы – изучение влияния элементов горизонтальной структуры верхнего полога и световых окон древостоя на развитие и распределение подроста в смешанных широколиственно-кедровых лесах.
Материал и методы. В работе использованы данные, полученные в результате детальных исследований на 4 постоянных пробных площадях (ПП) размером 0,8–1,0 га, заложенных на экологически равноценных местообитаниях в заповеднике «Бастак» [10]. Выбор участков для постоянных пробных площадей произведен с учетом критериев, рекомендованных для биогеоценотических исследований [8]. На каждой ПП проведено детальное картирование положений стволов деревьев и подроста, а также проекций их крон с помощью программы Crown 0.3.1 [2]. У каждого дерева измерена высота, диаметр, высота начала кроны и высота самой широкой части кроны. На основе этих данных были выделены микрогруппировки по доминантам ярусов древостоя, кустарников и травяного покрова и окна [3]. Микрогруппировки были классифицированы в типы по признакам сходства состава ярусов. Для каждого индивидуума подроста, находящегося в окне, было рассчитано расстояние до края окна.
При анализе успешности возобновления подрост был классифицирован в три функциональные группы: низкий (менее 0,5 м, развитие происходит в пределах травяного яруса), средний (0,51–1,5 м, развитие происходит внутри кустарникового яруса) и высокий (выше 1,51 м, выход подроста из кустарникового яруса).
Весьма полезной характеристикой при определении благоприятных условий развития оказалась максимальная высота подроста.
На четырех постоянных ПП было измерено 12763 особи подроста 21 вида. Для достоверного выявления закономерностей характера распределения подроста были проанализированы только виды, представленные на ПП более чем 100 особями.
Для оценки достоверности зависимости количественных характеристик подроста от условий его развития представленными категорийными переменными был использован однонаправленный дисперсионный анализ [12] с последующим тестом Tukey [21]. Статистический анализ реализован с помощью пакета Statistica 9.0.
Результаты исследований. Исследованные сообщества относятся к типу леса «холодно-влажные лещинные кедровники с пихтой, липой и желтой березой» [4] и ассоциации Ribesi maximowicziani-Pinetum koraiensis [17]. Доминантами древостоя в первом ярусе являются Pinus koraiensis и комплекс умеренных листопадных широколиственных и мелколиственных видов: Tilia amurensis , Fraxinus mandshurica , Betula costata . В формировании яруса всегда участвует Picea ajanensis . Анализ горизонтальной структуры позволил выделить 125 микрогруппировок, классифицированных в 6 типов (табл. 1). Выделено и описано 119 световых окон.
Таблица 1
Характеристика типов микрогруппировок
|
Тип микрогруппировки |
Доминанты первого яруса |
Доминанты подчиненных ярусов древостоя |
Доминанты кустарникового яруса |
|
Темнохвойная |
Abn , Pia |
Abn , Pia |
– |
|
Широколиственнокедровая |
Bec , Bel , Tia , Pik |
Abn , Acm , Act , Pik , Tia |
Corylus mandshurica, Eleuthe-rococcus senticocus |
|
Кедровая |
Pik |
Abn , Acm |
– |
|
Ясенево-кедровая |
Frm , Pik |
Abn , Pik |
Sorbaria sorbifolia |
|
Широколиственнохвойная |
Abn , Bec , Pia |
Abn , Acm , Act , Pia |
Acer ukurunduense , Actinidia kolomikta |
|
Широколиственная |
Bec , Bel , Bep, Tia |
Abn , Act , Pia, Pik |
Corylus mandshurica , Phila-delphus tenuifolius |
Примечание: Abn – Abies nephrolepis; Acm – Acer mono; Bec – Betula costata; Frm – Fraxinus mandshurica; Pia – Picea ajanensis; Pik – Pinus koraiensis; Tia – Tilia amurensis; Ull – Ulmus laciniata; Bel – Betula lanata, Bep – Betula platyphylla; Act – Acer tegmentosum.
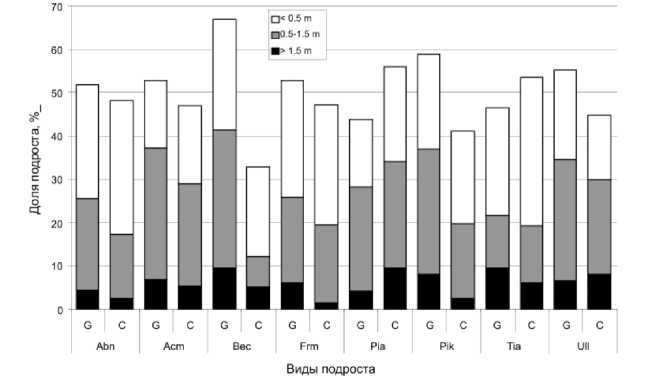
Рис. 1. Численность особей подроста в световых окнах (G) и под пологом леса (С) (пояснение сокращений названий типов микрогруппировок приводится в примечании к табл. 1)
Выделяется три типа распределения подроста разного размера. 1-й тип – возобновление преимущественно в окнах. Типичный пример – Betula costata (рис. 1). Мелкий подрост этого вида в массе встречается и под пологом, и в окнах. На стадии среднего подроста происходит его существенная дифференциация – под сомкнутым пологом он малочисленный, основная его масса сосредоточена в окнах, та же тенденция наблюдается и в распределении крупного подроста. Другими характерными представителями этого типа являются Pinus koraiensis и Fraxinus mandshurica. Однако наиболее резкая дифференциация по численности в окнах и под пологом происходит у этих видов уже на стадии крупного подроста. Abies nephrolepis и Tilia amurensis также принадлежат к этому типу, однако их мелкого подроста больше под пологом, чем в окнах. 2-й тип – нейтральный по отношению к пологу. Ближе к этому типу распределение подроста у Acer mono и Ulmus lacin-iata. 3-й тип представлен Picea ajanensis, у которой под пологом широколиственно-кедрового леса крупного подроста существенно больше, чем в окнах.
Анализ встречаемости подроста в микрогруппировках показывает, что теневыносливые виды, потенциальные строители подчиненного яруса древостоя, Abies nephrolepis и Acer mono наиболее часто встречаются в микрогруппировках, слагающих основу верхнего яруса широколиственно-кедрового леса (рис. 2). В то же время большая часть подроста этих видов сосредоточена в окнах. Наиболее успешно возобновление Abies nephrole-pis под пологом происходит под пологом широколиственных пород, там пихта имеет наибольшую долю крупного подроста. В то же время жизнеспособный подрост Acer mono встречается с равной вероятностью, как под кронами лиственных, так и хвойных деревьев. Подобный тип распространения характерен для Ulmus laciniata , основная часть крупного подроста которого приходится на темнохвойные микрогруппировки.
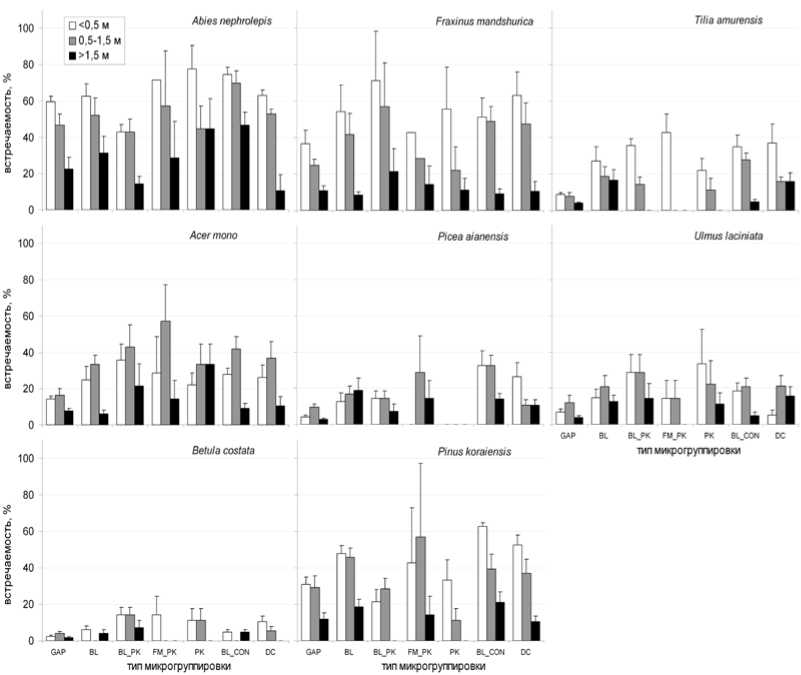
Рис. 2. Встречаемость подроста в различных типах микрогруппировок (Gap – окно; микрогруппировки: BL – широколиственная; BL_PK – широколиственно-кедровая; FM_PK – ясенево-кедроваяа;
BL_C – широколиственно-хвойная; PK – кедровая; DC – темнохвойная)
Виды широколиственных деревьев Betula costata , Fraxinus mandshurica и Tilia amurensis при возобновлении проявляют сильную зависимость от состояния верхнего яруса. Подрост Betula costata практически не встречается в микрогруппировках с участием Pinus koraiensis в верхнем пологе, а его основная часть сосредоточена в окнах и в смешанных микрогруппировках с доминированием широколиственных и темнохвойных видов, сформировавшихся также на месте окон. Для Fraxinus mandshurica характерно широкое распространение семян; его мелкий подрост распространен во всех типах микрогруппировок. Однако по мере роста он последовательно выпадает из состава микрогруппировок с участием хвойных, а затем и лиственных. Большая часть крупного подроста ясеня сосредоточена в окнах. Исключительно большой избирательностью по отношению к состоянию верхнего полога характеризуется Tilia amurensis . Ее мелкий подрост встречается повсеместно, однако встречаемость среднего подроста резко уменьшается в микрогруппировках с кедром, а основная доля крупного подроста приходится на микрогруппировки широколиственных и темнохвойных пород.
Распределение подроста Picea ajanensis и Pinus koraiensis также неравномерно и зависит от типа микрогруппировки. У ели максимальная встречаемость мелкого подроста – в микрогруппировках, в которых ель присутствует в первом ярусе древостоя. Подрост ели отсутствует в микрогруппировках кедра. Наиболее жизнеспособный подрост (по числу крупных экземпляров) отмечен в широколиственных микрогруппировках, а его количество в окнах и под пологом хвойных существенно ниже.
Анализ распределения подроста внутри окна (рис. 3) показывает, что подрост большинства видов деревьев – потенциальных эдификаторов, концентрируется на дистанции первых метров от края окна, что, с одной стороны, отражает потребности подроста в освещении, а с другой стороны, защитную функцию древесного полога, ослабляющего развитие покрова светолюбивых кустарников.
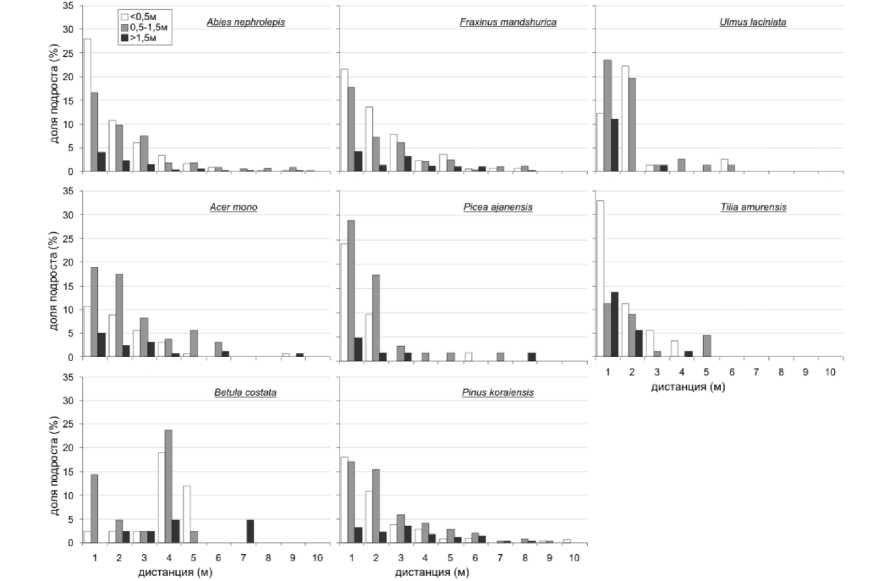
Рис. 3. Зависимость численности подроста от дистанции до края светового окна
Обсуждение результатов исследований. Pinus koraiensis принадлежит к группе видов, требующих окон для возобновления и образующих окна при выпадении из древостоя. В то же время, распространение кедра, облигатного зоохорного вида, зависит от предпочтений животных, создающих кладовки с его семенами в старовозрастных микрогруппировках, где существует сомкнутый верхний полог и отсутствуют густые подлесок и травостой [7]. Первые годы (десятилетия) жизни кедра обычно проходят под сомкнутым пологом, где он достигает высоты 4–6 (8) м. Если за 60–80 лет не происходит осветления полога, подрост кедра погибает. Если окно формируется, кедр начинает активно расти, давая в этот период максимальные в своем жизненном цикле приросты по диаметру [15]. В последующие 80–120 лет он достигает верхнего полога древостоя. Для поддержания непрерывного потока поколений [11] кедра в сообществе всегда должны формироваться окна, поэтому задаваемая ими мозаичная структура является характерной для широколиственно-кедровых лесов.
Возобновление Picea ajanensis происходит преимущественно под пологом, особенно интенсивно – в микрогруппировках с большим участием широколиственных пород. Высокая степень сгруппированности подроста ели в широколиственно-кедровых лесах объясняется его высокой требовательностью к влагообеспеченности местообитания. Подрост выживает только на участках аккумулирования влаги: выворотах, валеже, пнях, микропонижениях рельефа [6]. Ель не нуждается в окнах для возобновления, более того, избегает их, по-видимому, по причине чрезмерной инсоляции, но формирует окна при отмирании. При значительном участии ели в широколиственно-кедровом фитоценозе массовое возобновление ели отмечалось в периоды между пиками возобновления кедра и если стадии возобновления ели предшествовала стадия возобновления Acer mono [15].
Положение подроста Betula costata в окне ближе к центру и к северной границе окна указывает на его большую требовательность к условиям освещения. По отношению к окнам в пологе этот вид следует рассматривать как требующий окон для возобновления и образующий окна при выпадении. Tilia amurensis чаще возобновляется и успешно развивается под относительно светлым пологом, образованным широколиственными видами. Также оптимальными являются условия на краю окна.
В северных широколиственно-кедровых лесах Abies nephrolepis обычно формирует подчиненный ярус древостоя. Характеризуется коротким жизненным циклом (180–200 лет), сильной теневыносливостью и то-лерантна к осветлению полога. Ее подрост обилен в окнах и в микрогруппировках с большим участием лиственных деревьев. В случае масштабных нарушений полога (ветровалы, рубки) формирует сомкнутый ярус, существенно осложняющий возобновление других древесных видов.
Acer mono – наиболее теневыносливый широколиственный вид, по отношению к окнам в пологе характеризуется как не требующий окон для возобновления и не создающий окон при выпадении из древостоя.
Выводы
Таким образом, происходящие асинхронно возобновление, рост и отмирание деревьев являются необходимым условием для существования полидоминантного сообщества: возобновление видов различных по теневыносливости может происходить только в условиях полного или частичного осветления полога, а следовательно, и отмирания части деревьев, его слагающих, либо под сомкнутым пологом. Стадийность в развитии древостоя может проявляться в масштабе микрогруппировки, и значительно менее вероятно – в масштабе всего лесного сообщества.
