Размножение крапивника Troglodytes troglodytes в Ленинградской области: откладка и инкубация яиц, выкармливание птенцов, успешность размножения

Автор: Дорофеева М.Ю.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 87 т.8, 1999 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149032
IDR: 140149032
Breeding biology of the wren Troglodytes troglodytes in Leningrad region: the laying, incubation and nestling periods, nesting success
Текст статьи Размножение крапивника Troglodytes troglodytes в Ленинградской области: откладка и инкубация яиц, выкармливание птенцов, успешность размножения
Статья продолжает серию публикаций по биологии крапивника Troglodytes troglodytes в Ленинградской области, основанную на результатах стационарных исследований, проводимых нами с 1995 года в окрестностях дер. Коваши (Дорофеева 1996, 1999а,б; Дорофеева, Кудрявцева 1998, 1999).
Район исследования, материал и методика
Работу проводили в 1995-1999 в окрестностях дер. Коваши (59°55' с.ш., 29° 15' в.д., Ломоносовский р-н, Ленинградская обл.) на верхней террасе пред-глинтовой равнины, протянувшейся вдоль южного берега Финского залива. Терраса сложена валунными суглинками и глинами, на которых развиваются торфяно-подзолисто-глеевые и торфяные почвы, и занята долгомошными и сфагновыми сосняками и ельниками, а также вторичными берёзовыми и берёзово-осиновыми лесами. Здесь постоянно ведётся просечная рубка леса. Благодаря этому образуются обширные участки, сильно захламлённые отходами рубки и ветровалом, где и предпочитают селиться крапивники.
Основной нашей целью было изучение социодемографической системы крапивника, поэтому мы вели постоянные отлов, индивидуальное мечение и длительные наблюдения за опознаваемыми особями (всего помечено 24 самца, 56 самок и 364 птенцов и слётков). На территориях самцов отыскивали по возможности все гнёзда. Всего нам удалось найти и описать 460 гнёзд, из них 151 выводковое — т.е. выбранное самками для размножения.
Результаты и обсуждение
Откладка яиц
Привлечённая территориальным самцом самка, приняв его ухаживания, спаривается с ним и останавливает свой выбор на построенном и рекламируемом им гнездовом сооружении. После первых копуляций она начинает делать внутри постройки самца собственно гнездо, выстилая лоток мхом, затем шерстью и перьями. Деятельностью по выстилке лотка самка занимается одна, без помощи полового партнёра. На это уходит 3-4 дня, затем самка приступает к откладыванию яиц. Самец в это время активно поёт и сооружает новые гнёзда, пытаясь привлечь на свою территорию следующую самку. Первое время все гнездовые заботы лежат только на самке. Самец не кормит её во время откладки яиц и насиживания, хотя время от времени интересуется гнездом и, как правило, по-
Таблица 1. Сроки начала кладок у крапивника на стационаре “Коваши” (число кладок, начатых в каждой декаде)
В районе наших исследований крапивники обычно приступают к откладке яиц в начале мая. В самых ранних кладках первое яйцо появилось 2 мая 1995, в начале первой декады мая 1996, 16 мая 1997, 4 мая 1998 и 30 апреля 1999. Сроки откладки яиц сильно растянуты (табл. 1). Самая поздняя из найденных нами кладок была начата 20 июля 1998 и состояла из 4 яиц. Такие же сроки откладки яиц — начало мая-конец июля — приводят для Северо-Запада России и другие исследователи (Зимин 1972; Носков и др. 1981; Мальчевский, Пукинский 1983). В Ленинградской обл. наиболее поздняя откладка первого яйца зарегистрирована 1 августа 1977 в юго-восточном Приладожье (Носков и др. 1981).
Хорошо известно, что крапивники могут размножаться дважды в течение одного репродуктивного периода. В районе нашего исследования возможность второго размножения после успешного воспитания птенцов первого выводка подтверждено наблюдениями за самками, окольцованными цветными кольцами. Обычность нормальных вторых кладок у крапивника косвенно подтверждается и выраженностью второго пика в сезоном распределении времени начала кладок (табл. 1). Второй цикл размножения начинается во второй декаде июня. Сроки начала первых и вторых кладок в разных частях ареала представлены в таблице 2.
Судя по срокам начала кладок, количество первых и вторых кладок у крапивника в районе наших исследований примерно одинаково (табл. 3). Во всяком случае, различия статистически незначимы (U-критерий Уил-коксона: Uf = И > ий; п\ = и2 = 5; Р = 0.01). Это позволяет высказать предположение, что практически все самки выводили два выводка за сезон. Подтвердить это предположение наблюдениями за окольцованными особями довольно трудно, поскольку второй раз самка как правило размножается на территории другого самца и при смене партнёров часто уходит за пределы контролируемой нами территории. В большинстве случаев после окончания забот о первом выводке самки уходили с территории самца, а на их месте появлялись и гнездились новые самки, предположительно, вырастившие первые выводки на других территориях. В
1998 на участке исследования второй раз размножалась только 1 самка из 12 окольцованных при первом выводке, в 1999 — 5 из 13. Из этих 6 самок только 1 размножалась второй раз с тем же самцом и на той же гнездовой территории. Минимальное расстояние между гнёздами одной самки на нашем участке составило 32 м, максимальное — 360 м. По данным А.Ф.Ковшаря (1979), вторые гнёзда располагались в 0.15-60 м от первых. По наблюдениям М.Даллманна (Dallmann 1987) в Германии, это расстояние варьировало от 20 до 3000 м.
Сделанные разными авторами оценки доли самок крапивника, участвующих во втором цикле размножения, существенно различаются. В Ленинградской обл., по мнению А.С.Мальчевского и Ю.Б.Пукинского (1983), ко второй кладке приступает не более четверти особей. Примерно такую же оценку приводят Е.С.Птушенко и А.А.Иноземцев (1968) для Московской обл. По данным М.Даллманна (Dallmann 1987), в Германии дважды размножается 60-70% самок.
Таблица 2. Сроки начала первых и вторых кладок у крапивника в разных частях ареала
|
Регион Первая кладка |
Вторая кладка |
Источник |
|
Великобритания Конец марта-май |
Начало июня- |
Garson 1980 |
|
Нидерланды Конец апреля-май |
середина июля 2-я декада июня |
Kluijver et al. 1940 |
|
Германия Апрель- |
2-я декада июня |
Dallmann 1987 |
|
1 -я декада июня Ленинградская обл. Начало мая |
3-я декада июня |
Носков и др. 1981 |
|
Ленинградская обл. Начало мая |
Середина июня |
Мальчевский, Пукинский. 1983 |
|
Ленинградская обл. Начало мая |
2-я декада июня |
Наши данные |
|
Московская обл. Начало мая |
3-я декада июня |
Птушенко, Иноземцев 1968 |
|
Тянь-Шань Начало мая |
2-я половина июня |
Родионов 1968 |
|
Тянь-Шань Начало мая |
2-я половина июня |
Ковшарь 1979 |
Таблица 3. Число и средняя величина первых и вторых кладок у крапивника на стационаре “Коваши”
|
Год |
Число кладок |
Среднее число яиц в кладке* |
||
|
1 -я кладка |
2-я кладка |
1 -я кладка |
2-я кладка |
|
|
1995 |
11 |
16 |
7.1 (8) |
5.6 (8) |
|
1996 |
20 |
22 |
7.5 (10) |
6.2 (18) |
|
1997 |
15 |
8 |
6.6 (8) |
6.0 (4) |
|
1998 |
14 |
10 |
6-4 (7) |
5.8 (10) |
|
1999 |
14 |
11 |
6.7(11) |
5.8 (8) |
|
В среднем |
12.4 |
13.4 |
6.8 (44) |
5.8 (48) |
* В скобках — число кладок, использованных для вычисления среднего.
Величина кладки и размеры яиц
В районе исследования кладки крапивника содержат 4-8, чаще всего 6-7 яиц. Средняя величина кладки составляет 6.3 яйца (табл. 4 и 5). Величина кладки уменьшается к концу репродуктивного сезона. Отрицательная связь между началом кладки и её величиной статистически значима: г = - 0.54±0.1; п = 84; Р < 0.001. Влияние фактора значимо, но не велико — 3.6% (табл. 6). Попарное сравнение средних выявило значимые различия между величиной кладок, начатых в мае и июне, в мае и июле. Аналогичные результаты получены П.Гарсоном (Garson 1980) на английских крапивниках: г = -0.224; п = 444; Р < 0.001. Влияние фактора “время сезона” значимо, но также относительно невелико. Средняя величина июльских кладок составила 4.5 яйца (и = 46), тогда как апрельских и майских — 5.9 яйца (п = 804). Уменьшение средней величины кладки в течение репродуктивного сезона отмечено и другими авторами (табл. 6).
По нашим данным, величина вторых кладок значимо ниже, чем первых (tf = 5.53 > tst = 1.96, Р < 0.05; табл. 3).
Данные о величине кладки крапивника в разных точках ареала, представленные в таблицах 7 и 8, свидетельствуют о существовании выражен-
Таблица 4. Частотное распределение числа яиц в полной кладке крапивника на стационаре “Коваши”
|
Год |
Число яиц в кладке |
Z |
||||
|
4 |
5 |
6 |
7 |
8 |
||
|
1995 |
— |
4 |
5 |
4 |
3 |
16 |
|
1996 |
— |
4 |
9 |
15 |
1 |
29 |
|
1997 |
— |
1 |
5 |
6 |
— |
12 |
|
1998 |
1 |
4 |
6 |
5 |
1 |
17 |
|
1999 |
— |
4 |
8 |
6 |
2 |
20 |
|
S |
1 |
17 |
33 |
36 |
7 |
94 |
Таблица 5. Средняя величина кладки крапивника в зависимости от сроков откладки первого яйца. Стационар “Коваши”
|
Год |
Апрель |
Май |
Июнь |
Июль |
Весь сезон |
|
1995 |
— |
7.3 (6) |
6.0 (7) |
5.3 (3) |
6.4 (16) |
|
1996 |
— |
6.9 (10) |
6.3 (16) |
6.0 (3) |
6.4 (29) |
|
1997 |
— |
6.6 (8) |
6.0 (4) |
— |
6.4(12) |
|
1998 |
— |
7.0 (5) |
6.0 (8) |
5.0 (4) |
6.1 (17) |
|
1999 |
7.0 (2) |
6.7 (9) |
5.6 (7) |
7.0(1) |
6.3 (19) |
|
В среднем |
7.0 (2) |
6.9 (38) |
6.0 (42) |
5.5 (11) |
6.3 (93) |
В скобках — число кладок ной географической изменчивости этого показателя. Корреляционный анализ выявил наличие значимой положительной связи средней величины кладки с широтой (г = 0.61±0.06; п = 10; Р < 0.001) и долготой местности (г = 0.75±0.21; п = 10; Р < 0.05). Увеличение величины кладки с юга на север и с запада на восток в пределах Европы характерна для многих птиц разных отрядов (Lack 1968). Крапивник не является исключением из этого правила.
Таблица 6. Сезонная изменчивость величины кладки крапивника в разных точках ареала
|
Регион |
Средняя величина кладки* |
Источник |
|||
|
Апрель |
Май |
Июнь |
Июль |
||
|
Великобритания |
5.4 (14) |
5.8 (60) |
5.2 (10) |
4.0 (4) |
Armstrong 1955 |
|
Великобритания |
5.9 (160) |
5.9 (644) |
5.4 (202) |
4.5 (46) |
Garson 1980 |
|
Швейцария |
6.1 (16) |
6.2 (40) |
5.9 (18) |
5.8 (4) |
Glutz 1962 |
|
Рейнланд, Германия |
5.9 (18) |
6.2 (120) |
5.5 (91) |
5.0 (23) |
Mildenberger 1984** |
|
Вюртемберг, Германия |
6.3 (35) |
6.2 (379) |
5.9 (191) |
5.0 (84) |
Dallmann 1987 |
|
Беловежская Пуща, Польша |
7.0 (8) |
6.8 (30) |
6.0 (6) |
Wesolowsky 1983 |
|
|
Ленинградская обл. |
— |
6.5 (4) |
6.1 (23) |
5.6 (9) |
Мальчевский, Пукинский 1983 |
|
Ленинградская обл. |
7-0 (2) |
6.9 (38) |
6.0 (42) |
5.5 (11) |
Наши данные |
|
Тянь-Шань |
— |
5.8 (20) |
5.3 (9) |
5.0 (4) |
Ковшарь 1979 |
* В скобках — число кладок
** цит. по: Glutz, Bauer 1985.
Таблица 7. Распределение числа яиц в кладке у крапивника в разных точках ареала
|
Регион |
Число яиц в кладке |
N |
Источник |
||||||
|
<3 |
4 |
5 |
6 |
7 |
8 |
9 |
|||
|
Великобритания |
— |
— |
30 |
50 |
12 |
1 |
— |
93 |
Armstrong 1955 |
|
Великобритания |
40 |
93 |
280 |
487 |
153 |
40 |
13 |
1068 |
Garson 1980 |
|
Нидерланды |
2 |
5 |
27 |
45 |
11 |
— |
— |
90 |
Kluijver et al. 1940 |
|
Вюртемберг, Германия |
— |
— |
44 |
386 |
212 |
12 |
— |
654 |
Dallmann 1987 |
|
Восточная Германия |
— |
— |
10 |
18 |
10 |
4 |
— |
42 |
Makatsch 1976 |
|
Беловежская Пуща, Польша |
— |
1 |
2 |
9 |
28 |
4 |
— |
44 |
Wesolowski 1983 |
|
Финляндия |
— |
— |
3 |
4 |
13 |
2 |
1 |
23 |
Haartmann 1969 |
|
Ленинградская обл. |
— |
3 |
9 |
23 |
14 |
6 |
1 |
56 |
Мальчевский, Пукинский 1983 |
|
Ленинградская обл. |
— |
1 |
17 |
33 |
36 |
7 |
— |
94 |
Наши данные |
|
Тянь-Шань |
— |
3 |
7 |
23 |
— |
— |
— |
33 |
Ковшарь 1979 |
Таблица 8. Средняя величина кладки крапивника в разных точках ареала
|
Географические |
Средняя величина кладки |
lim |
/V |
Источник |
|
|
к |
оординаты |
||||
|
52° |
с.ш., 00° в.д. |
5.6 |
5-8 |
88 |
Armstrong 1955 |
|
52° |
с.ш., 01° в.д. |
5.7 |
2-9 |
1052 |
Garson 1980 |
|
47° |
с.ш., 08° в.д. |
6.1 |
— |
78 |
Glutz 1962 |
|
50° |
с.ш., 05° в.д. |
5.6 |
2-7 |
90 |
Kluijver et al. 1940 |
|
50° |
с.ш., 07° в.д. |
5.8 |
— |
252 |
Mildenberger 1984* |
|
48° |
с.ш., 09° в.д. |
6.0 |
5-8 |
689 |
Dallmann 1987 |
|
51° |
с.ш., 14° в.д. |
5.5 |
3-7 |
10 |
Pannach 1990 |
|
52° |
с.ш., 13° в.д. |
6.2 |
5-8 |
42 |
Makatsch 1976 |
|
52° |
с.ш., 23° в.д. |
6.7 |
4-8 |
44 |
Wesolowski 1983 |
|
65° |
с.ш., 25° в.д. |
6.7 |
5-9 |
23 |
Haartmann 1969 |
|
60° |
с.ш., 30° в.д. |
6.3 |
4-9 |
56 |
Мальчевский, Пукинский 1983 |
|
60° |
с.ш., 29° в.д. |
6.3 |
4-8 |
94 |
Наши данные |
|
62° |
с.ш., 35° в.д. |
8.5 |
7-10 |
10 |
Зимин 1972 |
* цит. по: Glutz, Bauer 1985.
Данные о средних размерах яиц крапивника в окрестностях стационара “Коваши” в сравнении с размерами яиц в других точках ареала представлены в таблице 9.
Таблица 9. Средние размеры яиц крапивника в разных точках ареала
|
Регион |
Подвид |
Средние размеры яиц, мм |
N |
Источник |
|
о-в Сент-Килда |
hirtensis |
18.55x13.93 |
100 |
in: Armstrong 1955 |
|
Шетландские о-ва |
zetlandicus |
18.58x13.58 |
25 |
in: Armstrong 1955 |
|
Великобритания |
troglodytes |
16.66x12.77 |
100 |
in: Dallmann 1987 |
|
Средняя Европа |
troglodytes |
16.42x12.45 |
72 |
Rey 1905 |
|
Бельгия |
troglodytes |
16.50x12.50 |
100 |
in: Glutz, Bauer 1985 |
|
Швеция |
troglodytes |
16.46x12.56 |
131 |
in: Makatsch 1976 |
|
Западная Германия |
troglodytes |
16.60x12.52 |
657 |
Dallmann 1987 |
|
Восточная Германия |
troglodytes |
16.42x12.63 |
170 |
Makatsch 1976 |
|
Ленинградская обл. |
troglodytes |
16.47x12.24 |
14 |
Дорофеева 1996 |
|
о-в Сахалин |
fumigatus |
17.56x13.12 |
11 |
Нечаев 1991 |
|
Тянь-Шань |
tianschanicus |
17.20x12.80 |
29 |
Родионов 1968 |
Насиживание
Плотное насиживание начинается после откладки последнего яйца. Насиживает только самка. Самец не принимает участия в насиживании и не кормит самку в этот период. Однако он может появляться и беспокоиться около гнезда при появлении опасности.
По нашим данным, насиживание продолжается от 13 до 20 сут. В среднем по 22 гнёздам период насиживания составил 15.6 сут (табл. 10). По данным других исследователей, длительность периода насиживания у крапивника может изменяться от 12 до 21 сут, а средняя продолжительность его в разных частях ареала составляет 15.1-16.0 сут (табл. 11).
При этом продолжительность периода насиживания уменьшается в ходе репродуктивного сезона (табл. 10). Корреляционный анализ показал наличие значимой отрицательной связи между сроками начала кладки и длительностью её насиживания: г = -0.65± 0.17; п = 20; Р < 0.001. Влияние фактора “время сезона” оказалось значимым и сильным — 82.8%
Таблица 10. Продолжительность периода насиживания у крапивника на стационаре “Коваши”
|
Год |
lim, сут |
Средняя продолжительность насиживания, сут* |
|||
|
Май |
Июнь |
Июль |
Весь репродуктивный сезон |
||
|
1995 |
15 |
— |
15.0 (3) |
— |
15.0 (3) |
|
1996 |
14-16 |
— |
15.0 (5) |
15.0 (1) |
15.0 (6) |
|
1997 |
20 |
20.0 (1) |
— |
— |
20.0 (1) |
|
1998 |
14-17 |
— |
14.7 (3) |
17.0(1) |
15.3 (4) |
|
1999 |
13-20 |
18.3 (3) |
14.0 (4) |
15.0 (1) |
15.8 (8) |
|
В среднем |
13-20 |
18.8 (4) |
14.7 (15) |
15.7 (3) |
15.6 (22) |
* В скобках — число кладок под наблюдением.
Таблица 11. Продолжительность периода насиживания у крапивника в разных точках ареала
|
Регион |
lim, сут |
Средняя продолжительность насиживания, сут* |
||||
|
Весь сезон |
Апрель |
Май |
Июнь |
Июль |
||
|
Нидерланды1) 14-20 16.0(43) 17.5(8) 16.3(17) 15.3(14) 14.5(4) Западная Германия2) 12-21 15.2(286) - 16.3(171) 15.1(98) 14.4(69) Восточная Германия3) 13-20 15.1 (53) — — — — Ленинградская обл.4) 13-20 15.6(22) — 18.8(4) 14.7(15) 15.7(3) Тянь-Шань5) 15-17 16.0(3) — — — — Тянь-Шань6* 15-16 15.8(5) — — — - * В скобках — число кладок под наблюдением. 1) Kluijver etal. 1940; 2) Dallmann 1987; 3) Creutz 1956; 4) Наши данные; 5) Родионов 1970; 6) Ковшарь 1979. |
||||||
(показатель силы влияния фактора Снедекора). Данные других исследователей также показывают уменьшение продолжительность периода насиживания в ходе репродуктивного сезона (табл. 11). М.Даллманн (Dall-mann 1987) видит причину этого в более плотном насиживании поздних кладок, что становится возможным благодаря большему обилию корма и меньшими затратами времени на его поиск в летние месяцы по равнению с весенними. Отчасти разделяя эту точку зрения, мы, кроме того, полагаем, что определённый вклад в уменьшение продолжительности периода насиживания в ходе сезона вносит и ускорение становления ритма инкубации при более поздних кладках.
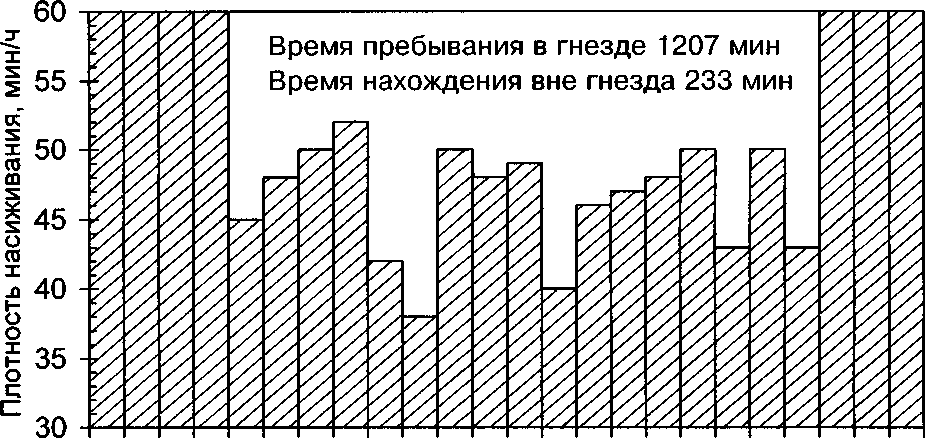
16-17 июня 1995 мы провели суточные наблюдения за одним гнездом в середине периода насиживания (рис. 1). Самка находилась в гнезде 83.8% времени суток. Вне гнезда она провела 233 мин (3.9 ч). Утром (59 ч) и вечером (19-22 ч) самка вылетала из гнезда 1 раз в час на 2-18 мин. С 9 до 18 ч, она вылетала кормиться 2 раза в час. Сходный ритм активности насиживающей самки крапивника описывает А.Ф.Ковшарь (1979).
При подозрительном шуме насиживающая самка выглядывает из гнезда, а затем затаивается в нём. При опасности она лишь в крайнем случае покидает гнездо и молча прячется в ближайшем укрытии.
О 2 4 6 8 10 12 14 16 18 20 22
Время суток, ч
Рис. 1. Изменение времени, проводимого насиживающей самкой крапивника в гнезде, на протяжении суток. 16-17 июня 1995.
Выкармливание птенцов
После вылупления птенцов самка всегда выносит скорлупу яиц. Яйца-“болтуны” и “задохлики” обычно остаются в гнезде.
В районе исследований у крапивника кормит птенцов, как правило, одна самка. Самцы участвовали в выкармливании выводка лишь в 15% случаев. При этом мы ни разу не наблюдали, чтобы самцы кормили птенцов до достижения ими возраста 7 сут. Если птенцов кормили оба партнёра, самец приносил корм реже, чем самка, затрачивал на его сбор больше времени (в среднем 10 мин) и улетал за кормом дальше, за 100 м и более. Самка обычно собирала корм в радиусе до 50 м вокруг гнезда. На сбор одной порции корма она затрачивала 1-15, чаще 4-5 мин.
Суточные наблюдения за частотой кормления птенцов проведены в июне-июле 1995 на гнёздах с птенцами в возрасте 3-7 сут (рис. 2). Во всех этих случаях птенцов кормила одна самка. Частота прилётов с кормом варьировала от 1 до 17 раз/ч, медиана 8 раз/ч (58 ч наблюдений). В течение дня неявно выделяются три пика активности по кормлению птенцов: утренний (5-8 ч), дневной (около полудня) и вечерний (20-23 ч).
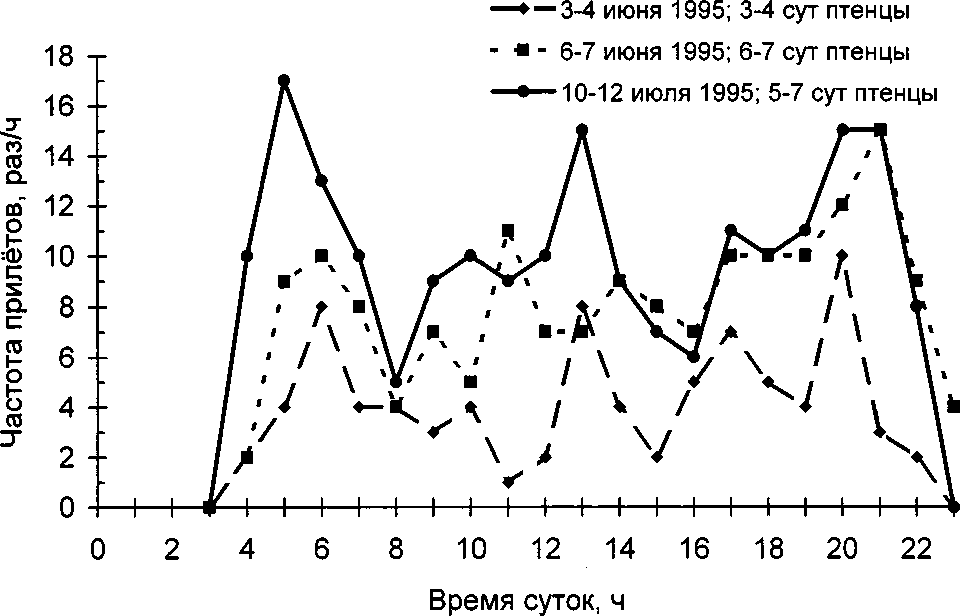
Рис. 2. Изменение частоты кормления птенцов самкой крапивника на протяжении суток.
Вылет птенцов из гнёзд. Послегнездовая жизнь выводка
Уже в возрасте 10 сут птенцы крапивника могут покинуть гнездо в случае опасности. При отсутствии беспокойства вылет обычно происходит на 15-19-е сут. Чаще всего птенцы вылетают в возрасте 16 сут. В среднем по 30 гнёздам птенцы вылетали в возрасте 16.8±0.22 сут.
Первые 2-3 нед. выводок держится на гнездовом участке. На ночь птенцы могут возвращаться в гнездо, но чаще ночуют под корягами, в корнях и ветвях поваленных деревьев. В 3 случаях слётки ночевали в не-использовавшихся для размножения самцовых гнёздах. Они до 8-9 ч не покидали гнёзда, а самка не только приносила туда им корм, но и выносила фекальные капсулы.
Согласно другим авторам, вылет птенцов крапивника происходит на 14-19-е сут, чаще на 15-16-е. По данным одних орнитологов, выводки распадаются через 2-3 нед. после оставления гнезда (Kluijver et al. 1940; Armstrong 1955; Dallmann 1987; Мальчевский, Пукинский 1983), по данным других — уже через 1 нед. (Птушенко, Иноземцев 1968; Родионов 1968).
Самец часто сопровождает перемещающийся по его территории выводок и громко трещит около него. По нашим наблюдениям в 1998-1999, самцы принимали участие в кормлении слётков в 21% случаев, тогда как в гнезде они кормили птенцов в 15% случаев. Птенцов вторых выводков самцы кормят чаще. В 1998 они участвовали в кормлении 9% первых и 13% вторых выводков; в 1999 — соответственно, 14% и 63%. По усреднённым данным за оба года, самцы кормили птенцов в 11% первых выводков и 38% вторых. По наблюдениям М.Даллманна (Dallmann 1987), обычно оба партнёра сопровождают выводок, но самец кормит слётков реже. Е.Армстронг (Armstrong 1955) пишет, что самцы участвуют в кормлении птенцов в 40% случаев и в одиночку докармливают выводок, если самка уже приступила к насиживанию второй кладки. В двух других исследованиях оказалось, что во всех гнёздах самки кормили птенцов до их вылета без помощи самцов (Kluijver et al. 1940; Garson 1978). Г.Клюйвер отмечает также, что существенную помощь в выкармливании птенцов самцы начинают оказывать только в конце сезона размножения. Таким образом, степень участия самцов в выкармливании птенцов сильно зависит от времени репродуктивного сезона и значительно различается в разных частях ареала.
Успешность гнездования
Рассчитанная традиционным способом успешность размножения (отношение числа успешно вылетевших птенцов к общему числу отложенных яиц) в среднем за 1995-1999 составила 60.7% (табл. 12). Всего из 621 отложенного яйца вылупилось 73.8% птенцов (успешность вылупления), из 458 вылупившихся птенцов 82.3% вылетело из гнёзд (успешность
Таблица 12. Успешность размножения крапивника на стационаре “Коваши”
Потери на стадиях откладки яиц и насиживания как правило выше, чем на стадии выкармливания птенцов в гнезде. Значительная часть потерь яиц обусловлена т.н. эмбриональной смертностью, т.е. неоплодотво-рённостью яиц и гибелью эмбрионов. У крапивника среди неразвивших-ся яиц преобладают т.н. “болтуны” — неоплодотворённые яйца и яйца с эмбрионами, погибшими на самых ранних стадиях развития. Доля болтунов варьировала в разные годы от 4.3% (и = 94) до 11.8% (и = 153) и в целом за 5 лет составила 8.5% (и = 504). Сходная доля неоплодотворённых яиц (около 8%) наблюдалась у крапивника в Германии (Dallmann 1987).
В разные годы от 5 до 12, в среднем 9% гнёзд оказывались брошенными по неизвестным нам причинам.
По наблюдениям М.Даллманна (Там же) в Германии значительная часть гнёзд крапивника (30%) разоряется ещё до завершения кладки. На стадии выкармливания птенцов гибнет только 13% выводков. В Беловежской Пуще до 60% гнёзд разоряется во время откладки и насиживания яиц (Wesolowski 1983). Согласно М.Даллманну, гнёзда крапивников в основном разоряют белки Sciurus vulgaris, горностаи Mustela erminea, мыши Apodemus, бурозубки Sorex и сойки Garrulus glandarius. Иногда гнёзда крапивника занимают шмели Bombus, а также слизни Arion rufus, обильно покрывающие слизью содержимое гнезда. Много гнёзд гибнет в Германии из-за лесоустроительных работ и другой хозяйственной деятельности человека.
В районе наших исследований основными разорителями гнёзд крапивника являются белки, куньи (Mustela erminea, М. nivalis, Martes martes), мыши Apodemus и полёвки. Одно гнездо разорила сойка. Шмели и осы Vespula обычно поселялись в не используемых для размножения гнёздах.
