Разнообразие аллельного состояния генов Glu-1 в коллекции образцов твердой пшеницы (Triticum durum Desf.)
")
Автор: Коробкова В.А., Крупина А.Ю., Архипов А.В., Яновский А.С., Воропаева А.Д., Беспалова Л.А., Мудрова А.А., Назарова Л.А., Магомедов М.М., Крупин П.Ю., Самарина М.А., Ульянов Д.С., Карлов Г.И., Дивашук М.Г.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы
Статья в выпуске: 5 т.58, 2023 года.
Бесплатный доступ
Зерно твердой пшеницы, которое используется в макаронной и крупяной промышленности, должно обладать высокими показателями качества клейковины, которое зависит от многих факторов, в том числе от аллельного состояния генов высокомолекулярных глютенинов, кодирующих субъединицы запасных белков пшеницы (HMW-GS). В настоящей работе впервые показана структура обширной коллекции сортов и селекционных линий-двуручек твердой пшеницы по сочетанию аллельных вариантов генов локуса Glu-1 . Цель работы состояла в идентификации аллельного состояния генов высокомолекулярных глютенинов Glu-A1 и Glu-B1 с применением методов SDS-PAGE и KASP-анализа в коллекции, включающей сорта и перспективные селекционные линии двуручек твердой пшеницы, и оценке их влияния на индекс глютена. Материалом для исследований служила коллекция из 198 селекционных линий и сортов-двуручек твердой пшеницы, собранная в НЦЗ им. П.П. Лукьяненко. Аллельное состояние HMW-GS оценивали методом SDS-PAGE. Величину индекса глютена измеряли с помощью системы Perten Glutomatic® 2100 System («PerkinElmer», США). Для выявления аллельных вариантов генов HMW-GS также применяли KASP-маркеры: Glu-Ax1/x2*_SNP для Glu-A1 и BX7OE_866_SNP для идентификации не отличимых с помощью SDS-PAGE аллелей Glu-B1b (Bx7 + By8) и Glu-B1al (Bx7OE + By8). Маркер BX7OE_866_SNP основан на SNP в промоторной области: вариант С ассоциирован с аллелем Glu-B1al , несущим двойную копию Bx7 в результате дупликации локуса, вариант G - с отсутствием двойной копии Bx7. Вывод об аллельном состоянии генов HMW-GS Glu-A1 и Glu-B1 делали на основании сравнения данных, полученных с помощью KASP-анализа и SDS-PAGE. По результатам исследований было выявлено три аллеля в локусе Glu-A1 и восемь - в локусе Glu-B1 . В подавляющем большинстве изученных образцов присутствовал аллель Glu-A1c (98 %), а на долю образцов с Glu-A1a и Glu-A1b приходилось соответственно 0,5 и 1,5 %. По локусу Glu-B1 наблюдалось численное превосходство образцов с аллелем Glu-B1al (60 %) над образцами, несущими Glu-B1d (17 %) и Glu-B1e (12 %). На долю образцов с субъединицами Bx7 + By8 ( Glu-B1b ) пришлось 3 % исследованных образцов. Мы также выявили редкие аллели Glu-B1h (1 %), Glu-B1i (1 %), а также Glu-B1z (1 %) и Glu-B1z* (5 %). Два последних различались однонуклеотидным полиморфизмом в промоторной области гена, кодирующего Bx7 субъединицу. Прослеживалась тенденция к распространению и закреплению в сортах твердой пшеницы аллеля Glu-B1z* . Также было отмечено положительное влияние Glu-B1d на индекс глютена и негативный эффект Glu-B1e на его величину.
Triticum durum, твердая пшеница, высокомолекулярные глютенины, hmw-gs, glu-a1, glu-b1, kasp-маркер, sds-page, индекс глютена
Короткий адрес: https://sciup.org/142239854
IDR: 142239854 | УДК: 633.11:631.51:577.2 | DOI: 10.15389/agrobiology.2023.5.840rus
Study of the allelic diversity of the Glu-1 genes in the durum wheat (Triticum durum Desf.) collection
Durum wheat grain, which is used in the production of pasta and groats, must meet requirements for specific parameters of gluten quality. Its high quality depends on various factors, including the allelic state of the high-molecular-weight glutenin genes that encode subunits of wheat storage proteins (HMW-GS). This work presents, for the first time, the allelic structure of the Glu-1 loci in a wide collection of facultative durum wheat varieties and breeding lines. The objective of this study was to identify the allelic state of the Glu-A1 and Glu-B1 genes which encode high-molecular-weight glutenins in this collection. Additionally, the study aimed to evaluate the impact of these genes on the gluten index. The material for the research consisted of a collection of 198 breeding lines and varieties of facultative durum wheat obtained from the Lukyanenko National Grain Center. The allelic state of HMW-GS was assessed using SDS-PAGE. The gluten index value was measured using the Perten Glutomatic® 2100 System (PerkinElmer, USA). To identify allelic variants of HMW-GS genes, KASP markers were also used. The Glu-Ax1/x2*_SNP marker was utilized to identify the alleles of Glu-A1 . The BX7OE_866_SNP marker was employed to differentiate between the Glu-B1b (Bx7 + By8) and Glu-B1al (Bx7OE + By8) alleles, which cannot be distinguished through SDS-PAGE. The BX7OE_866_SNP marker is based on a single nucleotide polymorphism (SNP) in the promoter region: the C variant is associated with the Glu-B1al allele, which carries a duplicated copy of the Bx7 locus, while the G variant is associated with the absence of the Bx7 duplication. The allelic state of the Glu-A1 and Glu-B1 was identified by comparing the results obtained using the KASP and SDS-PAGE assays. According to the research results, three alleles were identified for Glu-A1 , and eight alleles were identified for Glu-B1 . The vast majority of the studied accessions contained the Glu-A1c allele (98 %), while the proportion of accessions with Glu-A1a and Glu-A1b accounted for 0.5 % and 1.5 %, respectively. For Glu-B1 , there was a higher number of accessions with the Glu-B1al allele (60 %) compared to accessions carrying Glu-B1d (17 %) and Glu-B1e (12 %). The accessions containing subunits Bx7 + By8 ( Glu-B1b ) accounted for 3 % of the collection. In addition, rare alleles Glu-B1h (1 %), Glu-B1i (1 %), as well as Glu-B1z (1 %) and Glu-B1z* (5 %) were identified. The last two differed in a single nucleotide polymorphism in the promoter region of the gene that encodes the Bx7 subunit. There was a tendency towards the distribution and fixation of the Glu-B1z* allele in durum wheat varieties. A positive effect of Glu-B1d on the gluten index and a negative effect of Glu-B1e on its value were also observed.
Текст научной статьи Разнообразие аллельного состояния генов Glu-1 в коллекции образцов твердой пшеницы (Triticum durum Desf.)
Твердая пшеница (Triticum durum Desf.) — одна из основных культур, возделываемых для производства продуктов питания. Ее зерно широко используется в макаронной и крупяной промышленности, а также в хлебопечении (1). Для получения высококачественной макаронной продукции зерно твердой пшеницы должно обладать соответствующими показателями по таким признакам, как стекловидность, содержание белка, клейковины и каротиноидных пигментов (2-4). В связи с тем, что потребители проявляют интерес к продуктам с улучшенными свойствами, возрастают и требования производителей к качеству сырья (5). Особое внимание уделяется клейковине, состояние которой оценивают с помощью индекса глютена, характеризующего силу внутри- и межмолекулярных связей. Индекс глютена также используют в селекционной работе для отбора наиболее перспективных линий твердой пшеницы наряду с такими классическими методами, как анализ фаринограмм и миксограмм или определение SDS-седиментации (6, 7). Индекс глютена крайне важен для современной технологии высокотемпературной сушки макаронных изделий, которая широко применяется в производстве пасты в Северной Америке, Австралии и странах Европы. Эта технология, увеличивающая как объем, так и эффективность производства макаронной продукции, используется и в России, однако ее широкому распространению в нашей стране и странах СНГ препятствует малое число сортов твердой пшеницы с требуемой генетической основой, обеспечивающей клейковину определенного качества (8, 9).
Качество клейковины твердой пшеницы, определяемое ее реологическими свойствами, зависит от компонентов глиадиновой и глютениновой фракций белка, соотношения глиадинов и глютенинов, аллельных вариантов глиадиновых и глютениновых локусов, а также от взаимодействия генотип—среда (10-12). Глютенины — запасные белки пшеницы, состоящие из связанных между собой многочисленных полипептидных цепей (субъединиц) (13, 14). Глютениновая фракция белка образуется высокомолекулярными (high molecular weight, HMW) и низкомолекулярными (low molecular weight, LMW) глютенинами. Гены, кодирующие высокомолекулярные глютенины, находятся в локусе Glu-1 , гены низкомолекулярных глютенинов локализованы в Glu-3 (15).
Высоко- и низкомолекулярные глютенины неодинаковы по своему влиянию на качество клейковины твердой пшеницы и макаронных изделий. На качество зерна и конечной продукции наибольшее влияние оказывают LMW-глютенины (10, 16, 17). Вместе с тем влияние HMW-глютенинов также немаловажно (11, 18, 19). Высокомолекулярные глютенины у твердой пшеницы кодируются локусами Glu-A1 и Glu-B1 , каждый из них кодирует x- и y-субъединицы, возникшие в результате древней дупликации, при этом y-субъединица у Glu-A1 не экспрессируется (13). О влиянии аллельного состояния Glu-A1 на качество изделий из твердой пшеницы нет однозначного мнения из-за фактической фиксации нуль-аллеля Glu-A1c в большинстве мировых коллекций современных сортов (19, 20). К аллелям Glu-B1 с положительным влиянием на качество клейковины твердой пшеницы и макаронных изделий относят, например, Glu-B1b (Bx7 + By8), Glu-B1al (Bx7OE + By8), Glu-B1d (Bx6 + By8), Glu-B1h (Bx14 + By15), Glu-B1z (Bx7 + By15), к аллелям с негативным эффектом — Glu-B1e (Bx20 + By20) (19, 21, 22).
Поскольку гены высокомолекулярных глютенинов характеризуются значительным разнообразием аллельных вариантов субъединиц белков, различающихся по степени влияния на качество зерна твердой пшеницы, вызывает интерес изучение распространения аллельных вариантов генов HMW-GS, а также сочетания их аллелей в разных коллекциях твердой пшеницы (16, 19, 23). Несмотря на точность таких методов детекции субъединиц запасных белков зерна, как lab-on-a-chip, жидкостная хроматография и масс-спектрометрия, электрофорез белков в полиакриламидном геле с додецилсульфатом натрия (sodium dodecyl sulfate-polyacrylamide gel electrophoresis, SDS-PAGE) остается наиболее доступным и широко используемым (11, 21, 22).
Аллельное состояние генов Glu определяют с помощью классической полимеразной цепной реакции (ПЦР) с разделением продуктов амплификации в агарозном или полиакриламидном геле, а также конкурентной аллель-специфичной ПЦР, или KASP (kompetitive allele specific PCR) (24). KASP-маркеры позволяют различать однонуклеотидные полиморфизмы между аллелями (single nucleotide polymorphisms, SNPs) за счет конкуренции между двумя прямыми праймерами — олигонуклеотидными последовательностями, которые комплементарны целевому участку ДНК, но различаются на 3′-конце целевыми SNP, а на 5′-конце имеют разные «хвосты» с уникальными последовательностями. В набор для KASP помимо двух альтернативных прямых и одного общего обратного праймера входят две FRET-кассеты (fluorescence resonance energy transfer, перенос энергии резонанса флуоресценции), каждая из которых представляет собой двуцепочечный олигонуклеотид. Одна цепочка FRET-кассеты с 3′-конца комплементарна уникальному «хвосту» того или иного прямого праймера, а на 5′-конце несет ковалентно связанный флуорофор (как правило, одна кассета несет FAM, другая — HEX); вторая цепочка FRET-кассеты (комплементарная первой) на 3′-конце несет гаситель флуорофора.
В первом раунде ПЦР происходит отжиг одного из альтернативных прямых праймеров: в конкуренции между прямыми альтернативными праймерами преимуществом обладает праймер, соответствующий SNP матричной ДНК, с которой проходит ПЦР. Во втором раунде ПЦР происходит отжиг обратного (общего) праймера. В третьем раунде ПЦР на «хвосте» прямого праймера происходит отжиг той из двух FRET-кассет, которая комплементарна «хвосту» прямого праймера, имевшего преимущество. В результате происходит пространственная изоляция флуорофора и гасителя и наблюдается испускание сигнала (FAM или HEX). В случае если анализируется гетерозигота, детектируются одновременно два сигнала (24). Благодаря тому, что KASP-метод основан на детекции сигнала в ПЦР в реальном времени, в нем отсутствует этап разделения фрагментов в геле, что значительно ускоряет процедуру генотипирования.
Использование маркеров системы KASP позволяет эффективно выявлять как однонуклеотидные полиморфизмы, так и инсерции/делеции специфического участка ДНК (25). Основой KASP-маркеров могут быть как ранее созданные классические ПЦР-маркеры, разработанные на полиморфизмы между аллелями генов, так и однонуклеотидные полиморфизмы, выявленные методами ассоциативной генетики с помощью высокопроизводительного SNP-генотипирования (26). KASP-маркеры широко применяются в селекции сельскохозяйственных растений, в том числе пшеницы, благодаря высокой точности, эффективности и производительности, что позволяет выполнять масштабный скрининг коллекций сортов, расщепляющихся популяций и селекционных линий (27). KASP-маркеры используются для анализа аллельного состояния генов хозяйственно ценных признаков пшеницы, в том числе генов запасных белков (28-30).
В предыдущих исследованиях в коллекциях сортов и селекционных образцов озимой твердой пшеницы (23) и сортообразцов из питомника конкурсного сортоиспытания яровой твердой пшеницы (21), собранных в Национальном центре зерна (НЦЗ) им. П.П. Лукьяненко, мы проанализировали эффекты аллельных вариантов Glu-A1 и Glu-B1 в отношении качества зерна и макаронных изделий.
В настоящей работе впервые определена структура обширной коллекции сортов и селекционных линий-двуручек твердой пшеницы по сочетанию аллельных вариантов генов локуса Glu-1 .
Цель работы состояла в идентификации аллельного состояния генов высокомолекулярных глютенинов Glu-A1 и Glu-B1 с применением методов SDS-PAGE и KASP-анализа в коллекции, включающей сорта и перспективные селекционные линии двуручек твердой пшеницы, и оценке их влияния на индекс глютена.
Методика. Материалом для исследований служила коллекция из 198 селекционных линий и сортов-двуручек твердой пшеницы, собранная в НЦЗ им. П.П. Лукьяненко.
Аллельное состояние HMW-GS оценивали методом SDS-PAGE согласно ранее описанной методике (23, 31). Величину индекса глютена измеряли с помощью системы Perten Glutomatic® 2100 System («PerkinElmer», США) согласно протоколу (23, 32).
Геномную ДНК выделяли из 4-суточных проростков СТАВ-методом с некоторыми изменениями с использованием титер-трубок для массового анализа (33).
Для выявления аллельных вариантов гена Glu-A1 применяли маркер Glu-Ax1/x2*_SNP, включающий FAM-праймер 5´-AAGTGTAACTTCTCC-GCAACG-3´ (соответствует нуль-аллелю Glu-A1c , Ax-null), HEX-праймер 5´-ACCTAAGTGTAACTTCTCCGCAACA-3´ (соответствует наличию аллеля Glu-A1a , кодирующего субъединицу Ax1, или аллеля Glu-A1b , кодирующего субъединицу Ax2*), общий праймер 5´-CGAAGAAGCTTGGCCTGGATAG-TAT-3´ (28).
Поскольку образцы с аллелем Glu-B1b (Bx7 + By8), несущие одну копию Bx7, и образцы с аллелем Glu-B1al (Bx7OE + By8), характеризующиеся чрезмерной экспрессией Bx7 в результате дупликации фрагмента локуса (34), неотличимы при анализе методом SDS-PAGE, был использован KASP-маркер BX7OE_866_SNP, включающий HEX-праймер 5´-GTGGAATATTA-GTGATGGCGTGAC-3´ (соответствует наличию аллеля Glu-B1al , кодирующего субъединицы Bx7ОЕ + By8), FAM-праймер 5´-GTGGAATATTAGTG-ATGGCGTGAG-3´ (соответствует отсутствию аллеля Glu-B1al ) и общий праймер 5´-TTCTTCTCTCGTTGGCCTTATCGC-3´ (33). Маркер основан на SNP в промоторной области: вариант С ассоциирован с аллелем Glu-B1al , несущим двойную копию Bx7, вариант G — с отсутствием двойной копии Bx7 (28, 29).
ПЦР проводили в 96-луночном ПЦР-планшете. ПЦР-смесь объемом 10 мкл содержала 5 мкл Master mix KBS-1050-102 с присутствием красителей FAM, HEX, ROX («LCG Biosearch Techhnologies», Великобритания), 1,5 пМ аллель-специфических праймеров, 3,75 пМ общего праймера и 5 мкл матричной ДНК (50 нг на лунку). Использовали протокол touchdown: 15 мин при 94 ° С (активация фермента); 20 с при 94 ° С, 60 с при 61-55 ° С (снижение температуры с каждым циклом на 0,6 ° С), 30 с при 94 ° С (10 циклов); 20 с при 94 ° С, 60 с при 55 ° С (35 циклов); считывание сигнала 60 с при 37 ° С. ПЦР проводили в BIO-RAD CFX96 («Bio-Rad Laboratories, Inc.», США) с использованием программного обеспечения Bio-Rad CFX Manager 3.1.
Вывод об аллельном состоянии генов HMW-GS Glu-A1 и Glu-B1 делали на основании сравнения результатов, полученных с помощью KASP-анализа и SDS-PAGE.
При статистической обработке данных по индексу глютена, выраженного в процентах, проводили трансформация logit (35), после чего все расчеты велись с преобразованными значениями. Для различных состояний входных категорий значимость различий среднего индекса глютена оценивали по критерию Тьюки для множественных парных сравнений при 95 % доверительном интервале для однофакторных моделей ANOVA .
Результаты. По результатам KASP-анализа и SDS-PAGE были получены данные о структуре коллекции селекционных линий и сортов-двуручек твердой пшеницы (рис. 1, приложение 1; полностью данные приложения по 198 образцам см. на .
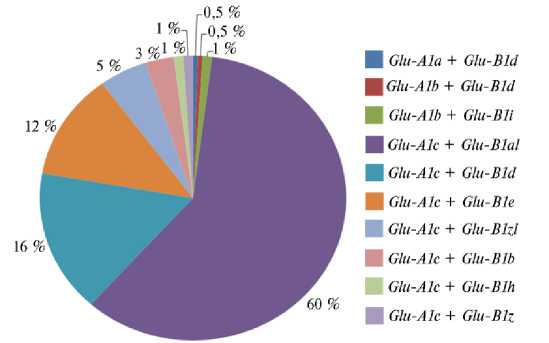
Рис. 1. Распределение сочетаний аллелей генов HMW-GS у образцов из коллекции селекционных линий и сортов-двуручек твердой пшеницы ( Triticum durum Desf.) (Национальный центр зерна имени П.П. Лукьяненко, 2021 год) .
Приложение 1. .Аллельные варианты генов HMW-GS в коллекции селекционных линий и сортов-двуручек твердой пшеницы I kiricum durum Dexf.). собранно!
|
в НПЗ имени П.П. Лукьяненко (Национальный центр зерна имени П.П. Лукьяненко. 2021 гол) |
|||
|
№ н/п |
Образен |
Аллель Glu-Al |
Аллель Glu-Bl |
|
1 |
480111142 |
Qu-Ak |
Glu-Blal |
|
2 |
48221119 |
Qu-A к |
Glu-Blal |
|
3 |
3902113-18-3 |
Qu-A lb |
Glu-BH |
|
4 |
46461)22 |
Qu-Ak |
Glu-Blal |
|
5 |
45831147 |
Qu-А к |
Glu-Blal |
|
6 |
47841126 |
Qu-Ak |
Glu-Blal |
|
7 |
429004 |
Qu-А к' |
Glu-Blal |
|
X |
47541176 |
Qu-Ak |
Glu-Blal |
|
Ч |
45071114 |
Qu-Ak |
Glu-Blal |
|
10 |
47991138 |
Qu-Ak |
Glu-BIb |
|
11 |
4387118 |
Qu-Ak |
Glu-Blal |
|
12 |
4782hb7 |
Qu-Ak |
Glu-BIb |
|
13 |
45031123 |
Qu-A к |
Glu-BIb |
|
14 |
42931125 |
Qu-Ak |
Glu-BIb |
|
15 |
4810054 |
Qu-Ak |
Glu-Blal |
|
10 |
4814020 |
Qu-A lb |
Glu-BH |
|
17 |
4291083 |
Qu-A к |
Glu-Blal |
|
18 |
4297012 |
Qu-Ak |
Glu-Blal |
|
19 |
4316090 |
Qu-Ak |
Glu-Blal |
|
20 |
42490103 |
Qu-A к |
Qu-Bid |
Установлено, что 194 образца из коллекции (98 %) имели аллель Glu-A1c , что свидетельствовало об отсутствии у них субъединицы Ax. Также были обнаружены образцы с аллелями Glu-A1a (1 образец) и Glu-A1b (3 образца), несущие соответственно субъединицы Ax1 и Ax2*.
По локусу Glu-B1 мы выявили 8 вариантов аллелей. Аллель Glu-B1b , соответствующий наличию субъединиц Bx7 + By8, был обнаружен у 6 образцов. Сочетание субъединиц Bx6 + By8, присущее Glu-B1d , выявлено у 34 образцов, что составляет 17 % изучаемой коллекции. Аллель Glu-B1e несли 24 образца с субъединицами Bx20 + By20. Редкими аллелями были Glu-B1h (Bx14 + By15) и Glu-B1i (Bx17 + By18), каждый из которых встречался только у двух образцов (соответственно у 3552h59-18-7 и COLOSSEO и у 4814h20 и 3902h3-18-3). Наибольшая часть коллекции (118 образцов, или 60 %) была представлена аллелем Glu-B1al (Bx7OE + Bx8), для которого характерна чрезмерная экспрессия субъединицы Bx7, обусловленная дупликацией фрагмента локуса (34). В образцах с субъединицами Bx7 + By15 наблюдался полиморфизм по KASP-маркеру: два образца (4636h22 и 4593h36) показали вариант G, 10 образцов — вариант С в промоторной области, в связи с чем первый вариант мы назвали Glu-B1z (Bx7 + By15), второй — Glu-B1z* , субъединицы условно обозначены Bx7(С) + By15 (рис. 2). Glu-A1a и Glu-A1b в изученной коллекции твердой пшеницы встречались только в комбинации с Glu-B1d и Glu-B1i . Glu-B1d также встречался в сочетании с Glu-A1c .
На рисунке 2 представлены паттерны белковых субъединиц высокомолекулярных глютенинов, идентифицированные в процессе анализа методом SDS-PAGE и дополненные результатами KASP-анализа.
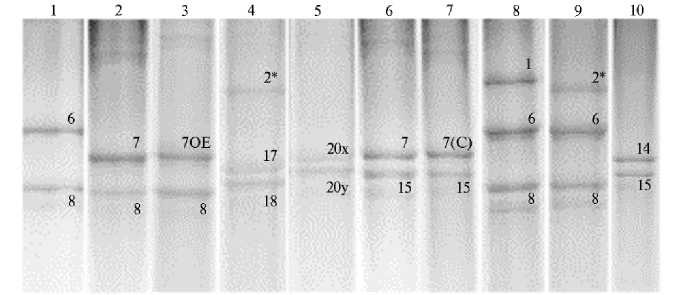
Рис. 2. Паттерны сочетаний субъединиц HMW-GS, детектированные у образцов из коллекции селекционных линий и сортов-двуручек твердой пшеницы ( Triticum durum Desf.) методом SDS-PAGE и подтвержденные с помощью KASP-анализа: 1 — 4598h48 (Bx6 + By8), 2 — ALLEMANNO (Bx7 + By8), 3 — 4583h47 (Bx7OE + By8), 4 — 4814h20 (Ax2*; Bx17 + By18), 5 —SAN MARCO (Bx20 + By20), 6 — 4593h36 (Bx7 + By15), 7 — 4808h8 (Bx7(C) + By15), 8 — 4626h42 (Ax1; Bx6 + By8), 9 — 4391h47 (Ax2*; Bx6 + By8), 10 — 3552h59-18-7 (Bx14 + By15) (Национальный центр зерна имени П.П. Лукьяненко, 2021 год) .
Влияние аллелей высокомолекулярных глютенинов и составляющих их субъединиц на индекс глютена отражено в таблице. Ввиду отсутствия выборки у образцов с сочетаниями аллелей Glu-A1a + Glu-B1d и Glu-A1b + Glu-B1d статистические данные не представлены.
Величина индекса глютена у образцов из коллекции селекционных линий и сортов-двуручек твердой пшеницы ( Triticum durum Desf.) в зависимости от аллелей и субъединиц HMW-GS (Национальный центр зерна имени П.П. Лукьяненко, 2021 год)
|
Фактор |
Индекс глютена, % |
Число образцов |
||
|
By |
By15 |
94,37ab |
14 |
|
|
By18 |
99,11ab |
2 |
||
|
By20 |
94,09b |
24 |
||
|
By8 |
97,63a |
158 |
||
|
Glu-A1 |
Glu-A1b |
98,91a |
3 |
|
|
Glu-A1c |
97,15a |
194 |
||
|
Glu-A1+Glu-B1 |
Glu-A1b + |
Glu-B1i |
99,11ab |
2 |
|
Glu-A1c + |
Glu-B1al |
97,46ab |
118 |
|
|
Glu-A1c + |
Glu-B1b |
98,29ab |
6 |
|
|
Glu-A1c + |
Glu-B1d |
97,98a |
32 |
|
|
Glu-A1c + |
Glu-B1e |
94,09b |
24 |
|
|
Glu-A1c + |
Glu-B1h |
89,45ab |
2 |
|
|
Glu-A1c + |
Glu-B1z |
97,81ab |
2 |
|
|
Glu-A1c + |
Glu-B1z* |
94,05ab |
10 |
|
|
Glu-B1 |
Glu-B1al |
97,46ab |
118 |
|
|
Glu-B1b |
98,29ab |
6 |
||
|
Glu-B1d |
98,03a |
34 |
||
|
Glu-B1e |
94,09b |
24 |
||
|
Glu-B1h |
89,45ab |
2 |
||
|
Glu-B1i |
99,11ab |
2 |
||
|
Glu-B1z |
97,81ab |
2 |
||
|
Glu-B1z* |
94,05ab |
10 |
П р им еч ани е. Статистически незначимые средние значения на основании критерия Тьюки обозначены одинаковыми буквами.
Максимальное значение индекса глютена в 2021 году наблюдалось у образцов с аллелем Glu-B1i. Ввиду низкой встречаемости этого аллеля среди изученных образцов нам не удалось выявить для него статистически значимых отличий от других, однако он вызывает несомненный интерес для дальнейшего изучения. Если рассматривать вклад каждой из субъединиц в повышение индекса глютена, то можно заметить, что присутствие By8 значительно увеличивало этот показатель. Статистически достоверное снижение индекса глютена связано с присутствием субъединицы By20, характерной для Glu-B1e.
Локус Glu-A1 в наших исследованиях был представлен тремя вариантами аллелей с преобладанием Glu-A1c (98 %). Преобладание аллеля Glu-A1c прослеживается и во многих других коллекциях твердой пшеницы вне зависимости от региона происхождения сортов (36-38). И хотя аллель Glu-A1c повсеместно распространен и зафиксирован даже у сортов современной селекции, среди образцов твердой пшеницы встречаются генотипы с аллелями Glu-A1a и Glu-A1b (19, 23, 38). В нашей коллекции на их долю пришлось соответственно 0,5 и 1,5 %. По сравнению с современными сортами повышенной частотой встречаемости аллеля Glu-A1a (до 38 %) характеризуются ландрасы и стародавние сорта Средиземноморья (37, 39). Аллели Glu-A1a и Glu-A1b представляют интерес для дальнейшего изучения, поскольку у образцов, в которых они комбинировались с аллелями локуса Glu-B1 , наблюдались высокие показатели индекса глютена. Например, наибольшее значение индекса глютена было отмечено в образце с сочетанием Glu-A1b + Glu-B1i , однако ввиду малой выборки нам не удалось выявить статистически значимых различий между этим сочетанием аллелей и другими сочетаниями (см. табл.).
Локус Glu-B1 был представлен 8 аллельными вариантами. Доминирующее положение среди всех обнаруженных вариантов занимал аллель Glu-B1al (60 %) с субъединицами Bx7OE + By8. В связи с тем, что SDS-PAGE не позволяет отличить Bx7OE + By8 ( Glu-B1al ) от Bx7 + By8 ( Glu-B1b ), мы провели анализ с помощью KASP-маркера, выявляющего полиморфизм С/G в промоторной области гена, кодирующего субъединицу Bx7 (28). В большинстве других исследований KASP-анализ не проводился, а состав субъединиц определялся с помощью SDS-PAGE. Если объединить полученные данные по Glu-B1b и Glu-B1al , то мы получим долю в 63 % (124 образца), близкую к показателям в европейских (37) и австралийских (38) коллекциях твердых пшениц. По нашим данным, образцы с аллелями Glu-B1al и Glu-B1b обладали высокими значениями индекса глютена (см. табл.).
Сочетание субъединиц Bx6 + By8 ( Glu-B1d ) в сортах мягкой пшеницы приводит к ухудшению хлебопекарных качеств, а в сортах твердой пшеницы, напротив, ассоциировано с высокими показателями качества клейковины (37, 38, 40). По нашим данным, образцы, несущие Glu-B1d , также характеризовались высокими показателями индекса глютена (см. табл.). Этот аллель умеренно распространен в средиземноморских сортах (37), где на его долю приходится около 20 % образцов, как и в наших исследованиях. Значительное распространение Glu-B1d наблюдается в сортах США и Канады (около 70 %) (38, 41).
Аллель Glu-B1e (Bx20 + By20) ухудшает показатели качества клейковины (22, 40, 42). В современных сортах он встречается реже, чем в ланд-расах и сортах старой селекции (16, 37). Хотя он и снижает значение индекса глютена, этот аллель присутствует (15-20 %) в современных сортах твердой пшеницы (19, 21, 39). Это может объясняться тем, что индекс глютена не единственный показатель, определяющий качество макаронных изделий. Поскольку Bx20+By20 по-прежнему встречается у современных сортов коммерческого использования, такие сорта интересны для поиска и изучения других генетических систем, определяющих высокое качество 846
макаронных изделий (LMW-GS, содержание крахмала и белка, глиадины, индекс цвета).
Интерес представляют выявленные редкие аллели Glu-B1h , Glu-B1z , Glu-B1z* , Glu-B1i , поскольку новые аллели позволяют разнообразить генофонд твердой пшеницы и расширить спектр технологических качеств продуктов питания, произведенных из сортов твердой пшеницы. Небольшая численность образцов с аллелями Glu-B1i (Bx17 + By18) и Glu-B1h (Bx14 + By15) не позволила в полной мере оценить их влияние на индекс глютена, однако они характеризовались соответственно самым высоким и самым низким средним значением индекса глютена (см. табл.). Другие исследователи отмечают сниженное значение показателя SDS-седиментации при оценке клейковины у образцов с субъединицами Bx14+By15 по сравнению с образцами, имеющими Bx17+By18 (20).
С помощью KASP-анализа мы выявили два варианта Glu-B1z (Bx7 + By15) c двумя вариантами SNP в промоторной части гена, кодирующего субъединицу Bx7. Образцы с полиморфизмом G — Glu-B1z (2 образца), образцы с полиморфизмом С были обозначены нами Glu-B1z* (10 образцов). Поскольку промотор влияет на экспрессию белка, можно ожидать, что выявленные SNP приводят к фенотипическим различиям между сортами, обладающими аллелями Glu-B1z и Glu-B1z* . Косвенно это подтверждают различия средних значений индекса глютена между образцами, несущими два указанных аллеля (см. табл.). Из-за малого объема выборки разница была статистически недостоверной, поэтому влияние этого SNP требует дальнейшего изучения. Кроме того, интересно, что число образцов с аллелем Glu-B1z* (5 %) преобладало над числом образцов с широко используемым в селекции твердой пшеницы аллелем Glu-B1b (3 %), то есть аллель Glu-B1z* постепенно распространяется и закрепляется в современных сортах твердой пшеницы.
Таким образом, в результате анализа 198 линий и сортов-двуручек твердой пшеницы из коллекции НЦЗ имени П.П. Лукьяненко мы установили, что в подавляющем большинстве изученных образцов присутствовал аллель Glu-A1c (98 %), а на долю образцов с Glu-A1a и Glu-A1b приходилось соответственно 0,5 и 1,5 %. По локусу Glu-B1 наблюдалось численное превосходство образцов с аллелем Glu-B1al (60 %) над образцами, несущими Glu-B1d (17 %) и Glu-B1e (12 %). На долю образцов с субъединицами Bx7 + By8 ( Glu-B1b ) пришлось 3 % от числа исследованных. Кроме того, мы выявили редкие аллели Glu-B1h (1 %), Glu-B1i (1 %), а также Glu-B1z (1 %) и Glu-B1z* (5 %). Два последних различались однонуклеотидным полиморфизмом в промоторной области гена, кодирующего субъединицу Bx7. Прослеживалась тенденция к распространению и закреплению в сортах твердой пшеницы аллеля Glu-B1z* . Также было отмечено положительное влияние Glu-B1d на индекс глютен и оказываемый на его величину негативный эффект Glu-B1e . Результаты анализа образцов двуручек, описанные в настоящей статье, вместе с ранее опубликованными данными, полученными при анализе коллекции озимой твердой пшеницы и перспективных сортообразцов яровой твердой пшеницы, расширяют представления о влиянии высокомолекулярных глютенинов на качество зерна, клейковины и макаронных изделий и могут быть полезны в селекционной работе по улучшению твердой пшеницы и созданию востребованных в производстве сортов.
Список литературы Разнообразие аллельного состояния генов Glu-1 в коллекции образцов твердой пшеницы (Triticum durum Desf.)
- Mastrangelo A.M., Cattivelli L. What Makes Bread and Durum Wheat Different? Trends in Plant Science, 2021, 26(7): 677-684 (doi: 10.1016/j.tplants.2021.01.004).
- Fu B.X., Wang K., Dupuis B., Taylor D., Nam S. Kernel vitreousness and protein content: relationship, interaction and synergistic effects on durum wheat quality. Journal of Cereal Science, 2018, 79: 210-217 (doi: 10.1016/j.jcs.2017.09.003).
- Мясникова М.Г., Мальчиков П.Н., Шаболкина Е.Н., Сидоренко В.С., Тугарева Ф.В., Ро-зова М.А., Чахеева Т.В., Цыганков В.И. Изменение содержания белка в зерне пшеницы твердой яровой в процессе селекции высокоурожайных сортов. Зернобобовые и крупяные культуры, 2019, 4(32): 112-119 (doi: 10.24411/2309-348X-2019-11141).
- Kaplan Evlice A. The effect of durum wheat genotypes on cooking quality of pasta. European Food Research and Technology, 2022, 248(3): 815-824 (doi: 10.1007/s00217-021-03899-4).
- Padalino L., Mastromatteo M., Lecce L., Spinelli S., Contò F., Del Nobile M.A. Effect of durum wheat cultivars on physicochemical and sensory properties of spaghetti. Journal of the Science of Food and Agriculture, 2014, 94(11): 2196-2204 (doi: 10.1002/jsfa.6537).
- Clarke F.R., Clarke J.M., Ames N.A., Knox R.E., Ross R.J. Gluten index compared with SDS-sedimentation volume for early generation selection for gluten strength in durum wheat. Canadian Journal of Plant Science, 2010, 90(1): 1-11 (doi: 10.4141/CJPS09035).
- Cecchini C., Bresciani A., Menesatti P., Pagani M.A., Marti A. Assessing the rheological prop-erties of durum wheat semolina: a review. Foods, 2021, 10(12): 2947 (doi: 10.3390/foods10122947).
- Шевченко С.Н., Мальчиков П.Н., Мясникова М.Г., Винченцо Н., Паскуале Д.В., Джулияни М. Генетические методы улучшения качества пшеницы твердых сортов, адаптированных к климатическим условиям России с особым акцентом на коммерческие характеристики зерна. Известия Самарского научного центра Российской академии наук, 2018, 20(2-2), 220-230.
- Шевченко С.Н., Мальчиков П.Н., Мясникова М.Г., Натоли В., Чахеева Т.В. Ускоренное создание с помощью методов геномной селекции высококачественных аналогов генотипов твердой пшеницы, адаптированных к условиям Среднего Поволжья. Достижения науки и техники АПК, 2019, 33(12): 38-42 (doi: 10.24411/0235-2451-2019-11207).
- Edwards N.M., Mulvaney S.J., Scanlon M.G., Dexter J.E. Role of gluten and its components in determining durum semolina dough viscoelastic properties. Cereal Chemistry, 2003, 80(6): 755-763 (doi: 10.1094/CCHEM.2003.80.6.755).
- Sissons M.J., Soh H.N., Turner M.A. Role of gluten and its components in influencing durum wheat dough properties and spaghetti cooking quality. Journal of the Science of Food and Agricul-ture, 2007, 87(10): 1874-1885 (doi: 10.1002/jsfa.2915).
- Fois S., Schlichting L., Marchylo B., Dexter J., Motzo R., Giunta F. Environmental conditions affect semolina quality in durum wheat (Triticum turgidum ssp. durum L.) cultivars with different gluten strength and gluten protein composition. Journal of the Science of Food and Agriculture, 2011, 91(14): 2664-2673 (doi: 10.1002/jsfa.4509).
- Gianibelli M.C., Larroque O., MacRitchie F., Wrigley C.W. Biochemical, genetic, and molecular characterization of wheat glutenin and its component subunits. Cereal Chemistry, 2001, 78(6): 635-646 (doi: 10.1094/CCHEM.2001.78.6.635).
- Воротынцева М.В. Степень влияния глютенинов на качество зерна как одного из сложных полигенных признаков рода Triticum (обзор). Труды по прикладной ботанике, генетике и селекции, 2021, 182(1): 168-185 (doi: 10.30901/2227-8834-2021-1-168-185).
- Pogna N.E., Autran J.C., Mellini F., Lafiandra D., Feillet P. Chromosome 1B-encoded gliadins and glutenin subunits in durum wheat: genetics and relationship to gluten strength. Journal of Cereal Science, 1990, 11(1): 15-34 (doi: 10.1016/S0733-5210(09)80178-1).
- Subira J., Peña R.J., Álvaro F., Ammar K., Ramdani, A., Royo C. Breeding progress in the pasta-making quality of durum wheat cultivars released in Italy and Spain during the 20th Century. Crop and Pasture Science, 2014, 65(1): 16-26 (doi: 10.1071/CP13238).
- Ruiz M., Bernal G., Giraldo P. An update of low molecular weight glutenin subunits in durum wheat relevant to breeding for quality. Journal of Cereal Science, 2018, 83: 236-244 (doi: 10.1016/j.jcs.2018.09.005).
- Raciti C.N., Doust M.A., Lombardo G.M., Boggini G., Pecetti L. Characterization of durum wheat Mediterranean germplasm for high and low molecular weight glutenin subunits in relation with quality. European Journal of Agronomy, 2003, 19(3): 373-382 (doi: 10.1016/S1161-0301(02)00091-6).
- Roncallo P.F., Guzmán C., Larsen A.O., Achilli A.L., Dreisigacker S., Molfese E., Astiz V., Echenique V. Allelic variation at glutenin loci (Glu-1, Glu-2 and Glu-3) in a worldwide durum wheat collection and its effect on quality attributes. Foods, 2021, 10(11): 2845 (doi: 10.3390/foods10112845).
- Al-Khayri J.M., Alshegaihi R.M., Mahgoub E.I., Mansour E., Atallah O.O., Sattar M.N., Al-Mssallem M.Q., Alessa F.M., Aldaej M.I., Hassanin A.A. Association of high and low molecular weight glutenin subunits with gluten strength in tetraploid durum wheat (Triticum turgidum spp. durum L.). Plants, 2023, 12(6): 1416 (doi: 10.3390/plants12061416).
- Kroupin P.Y., Bespalova L.A., Kroupina A.Y., Yanovsky A.S., Korobkova V.A., Ulyanov D.S., Karlov G.I., Divashuk M.G. Association of high-molecular-weight glutenin subunits with grain and pasta quality in spring durum wheat (Triticum turgidum spp. durum L.). Agronomy, 2023, 13(6): 1510 (doi: 10.3390/agronomy13061510).
- Giraldo P., Ruiz M., Ibba M.I., Morris C.F., Labuschagne M.T., Igrejas G. Durum wheat storage protein composition and the role of LMW-GS in quality. In: Wheat quality for improving processing and human health /G. Igrejas, T.M. Ikeda, C. Guzmán (eds.) Springer International Publishing, Cham, Switzerland, 2020: 73-108 (doi: 10.1007/978-3-030-34163-3_5).
- Kroupina A.Y., Yanovsky A.S., Korobkova V.A., Bespalova L.A., Arkhipov A.V., Bukreeva G.I., Voropaeva A.D., Kroupin P.Y., Litvinov D.Y., Mudrova A.A., Ulyanov D.S., Karlov G.I., Di-vashuk M.G. Allelic variation of Glu-A1 and Glu-B1 genes in winter durum wheat and its effect on quality parameters. Foods, 2023, 12(7), 1436 (doi: 10.3390/foods12071436).
- He C., Holme J., Anthony J. SNP genotyping: the KASP assay. In: Crop breeding: methods and protocols /D. Fleury, R. Whitford (eds.). Humana New York, NY, 2014: 75-86 (doi: 10.1007/978-1-4939-0446-4_7).
- Никитина Е.А., Архипов А.В., Минькова Я.В., Яновский А.С., Коробкова В.А., Самарина М.А., Черноок А.Г., Крупин П.Ю., Карлов Г.И., Дивашук М.Г. Конкурентная аллель-специфичная ПЦР (KASP): особенности, интерпретация результатов. Известия Тимиря-зевской сельскохозяйственной академии, 2022, 1(6): 79-93 (doi: 10.26897/0021-342X-2022-6-79-93).
- Semagn K., Babu R., Hearne S., Olsen M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): overview of the technology and its application in crop improvement. Molecular Breeding, 2014, 33: 1-14 (doi: 10.1007/s11032-013-9917-x).
- Allen A.M., Barker G.L.A., Wilkinson P., Burridge A., Winfield M., Coghill J., Uauy C., Grif-fiths S., Jack P., Berry S., Werner P., Melichar J.P.E., McDougall J., Gwilliam R., Robinson P., Edwards K.J. Discovery and development of exome-based, co-dominant single nucleotide poly-morphism markers in hexaploid wheat (Triticum aestivum L.). Plant Biotechnology Journal, 2012, 11(3): 279-295 (doi: 10.1111/pbi.12009).
- Rasheed A., Wen W., Gao F., Zhai S., Jin H., Liu J., Guo Q., Zhang Y., Dreisigacker S., Xia X., He Z. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theoretical and Applied Genetics, 2016, 129: 1843-1860 (doi: 10.1007/s00122-016-2743-x).
- Ravel C., Faye A., Ben-Sadoun S., Ranoux M., Dardevet M., Dupuits C., Exbrayat F., Pon-cet C., Sourdille P., Branlard G. SNP markers for early identification of high molecular weight glutenin subunits (HMW-GSs) in bread wheat. Theoretical and Applied Genetics, 2020, 133: 751-770 (doi: 10.1007/s00122-019-03505-y).
- Коробкова В.А., Беспалова Л.А., Яновский А.С., Воропаева А.Д., Архипов А.В., Шир-нин С.Ю., Черноок А.Г., Никитина Е.А., Ульянов Д.С., Мудрова А.А., Букреева Г.И., Карлов Г.И., Дивашук М.Г. Скрининг коллекции яровой и озимой твёрдой пшеницы с помощью KASP-маркера на аллельное состояние гена Zds. Кормопроизводство, 2023, 4: 25-31 (doi: 10.25685/KRM.2023.4.2023.004).
- Singh N., Shepherd K., Cornish G. A simplified SDS-PAGE procedure for separating LMW subunits of glutenin. Journal of Cereal Science, 1991, 14(3): 203-208 (doi: 10.1016/S0733-5210(09)80039-8).
- Perten Glutomatic-PERGM. PerkinElmer. Режим доступа: https://www.matt.nz/Prod-ucts/Grain/Grain-Applications/Flour-Milling/Glutomatic/. Дата обращения: 19.05.2023.
- Murray M.G., Thompson W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research, 1980, 8(19): 4321-4326 (doi: 10.1093/nar/8.19.4321).
- Ragupathy R., Naeem H. A., Reimer E., Lukow O.M., Sapirstein H.D., Cloutier S. Evolutionary origin of the segmental duplication encompassing the wheat GLU-B1 locus encoding the overex-pressed Bx7 (Bx7OE) high molecular weight glutenin subunit. Theoretical and Applied Genetics, 2008, 116: 283-296 (doi: 10.1007/s00122-007-0666-2).
- Warton D.I., Hui F.K. The arcsine is asinine: the analysis of proportions in ecology. Ecology, 2011, 92(1): 3-10 (doi: 10.1890/10-0340.1).
- Gregová E., Medvecká E., Jómová K., Šliková S. Characterization of durum wheat (Triticum durum Desf.) quality from gliadin and glutenin protein composition. Journal of Microbiology, Biotechnol-ogy and Food Sciences, 2021: 610-615.
- Nazco R., Peña R.J., Ammar K., Villegas D., Crossa J., Moragues M., Royo C. Variability in glutenin subunit composition of Mediterranean durum wheat germplasm and its relationship with gluten strength. The Journal of Agricultural Science, 2014, 152(3): 379-393 (doi: 10.1017/S0021859613000117).
- Sissons M.J., Ames N.P., Hare R.A., Clarke J.M. Relationship between glutenin subunit compo-sition and gluten strength measurements in durum wheat. Journal of the Science of Food and Agriculture, 2005, 85(14): 2445-2452 (doi: 10.1002/jsfa.2272).
- Moragues M., Zarco-Hernández J., Moralejo M.A., Royo C. Genetic diversity of glutenin protein subunits composition in durum wheat landraces [Triticum turgidum ssp. turgidum Convar. du-rum (Desf.) MacKey] from the Mediterranean Basin. Genetic Resources and Crop Evolution, 2006, 53: 993-1002 (doi: 10.1007/s10722-004-7367-3).
- Ammar K., Kronstad W.E., Morris C.F. Breadmaking quality of selected durum wheat geno-types and its relationship with high molecular weight glutenin subunits allelic variation and gluten protein polymeric composition. Cereal Chemistry, 2000, 77(2): 230-236 (doi: 10.1094/CCHEM.2000.77.2.230).
- Chegdali Y., Ouabbou H., Essamadi A., Cervantes F., Ibba M.I., Guzmán C. Assessment of the glutenin subunits diversity in a durum wheat (T. turgidum ssp. durum) collection from Morocco. Agronomy, 2020, 10(7): 957 (doi: 10.3390/agronomy10070957).
- Brites C., Carrillo J.M. Influence of high molecular weight (HMW) and low molecular weight (LMW) glutenin subunits controlled by Glu‐1 and Glu‐3 loci on durum wheat quality. Cereal Chemistry, 2001, 78(1): 59-63 (doi: 10.1094/CCHEM.2001.78.1.59).
