Разнообразие и особенности фоторецепторов в сетчатке камерного глаза брюхоногого моллюска Helicigona lapicida Linnaeus, 1758 (Pulmonata, Stylommatophora)
")
Автор: Шепелева И.П.
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Биохимия и физиология человека и животных
Статья в выпуске: 1, 2010 года.
Бесплатный доступ
Анализируется сетчатка наземного брюхоногого легочного моллюска Helicigona lapicida Linnaeus, 1758. Она содержит два типа микровиллярных фоторецепторов, которые представлены четырьмя видами: одним видом фоторецепторов первого типа и тремя видами фоторецепторов второго типа. Показано, что апикальная поверхность фоторецепторов первого типа имеет особенную выпуклость наподобие шляпки гриба. Фоторецепторы второго типа не выделяются какими-либо структурными особенностями, но различаются между собой диаметром дистальной части тела клетки, степенью ее вогнутости и длиной микровилл. Фоторецепторы первого типа более чувствительны к свету (0,5 мкм2 · ср-1) по сравнению с фоторецепторами второго типа (0,04, 0,008, 0,006 мкм2 · ср-1) за счет более длинных микровилл и большего диаметра апикальной поверхности. Варьирование чувствительности фоторецепторов второго типа обусловлено разным диаметром их дистальной поверхности и разной длиной микровилл. Присутствие в сетчатке нескольких видов фоторецепторов может означать наличие нескольких зрительных пигментов с разной спектральной чувствительностью и являться признаком цветового зрения.
Короткий адрес: https://sciup.org/14719496
IDR: 14719496
Текст научной статьи Разнообразие и особенности фоторецепторов в сетчатке камерного глаза брюхоногого моллюска Helicigona lapicida Linnaeus, 1758 (Pulmonata, Stylommatophora)
Введение. Фоторецепторные образования присутствуют у большинства групп животных и, несмотря на структурное разнообразие, обладают некоторыми общими свойствами, связанными с фундаментальным механизмом фоторецепции [10]. Ультраструк-турные исследования показали, что одним из таких свойств является увеличение площади клеточной мембраны, по которой распределен зрительный пигмент. Такое увеличение может происходить за счет выпячивания дистальной части тела клетки, формирующего многочисленные микровиллы, и в этом .случае называется микровиллярным или рабдомерным. Способ увеличения площади мембраны за счет развития микровилл широко распространен в рецепторных клетках беспозвоночных животных, включая брюхоногих моллюсков [8]. В их сетчатке можно обнаружить два типа рабдо-мерных фоторецепторов — первый и второй. Для выделения фоторецепторов в разные типы используют морфологические признаки: геометрию апикальной поверхности клеток, длину и толщину микровилл. Фоторецепторы первого типа имеют длинные тонкие микровиллы, которые отходят либо от апикального отростка, либо от более или менее выпуклой апикальной поверхности клетки. Дистальная поверхность фоторецепторов второго типа плоская или вогнутая и несет короткие и чаще всего более толстые микровиллы [6; 10; 17]. Сетчатка наземного брюхоногого легочного моллюска Helicigona lapicida Linnaeus, 1758 по своему клеточному составу отличается от сетчаток всех исследованных видов брюхоногих моллюсков: она содержит два типа микровиллярных фоторецепторов, которые представлены четырьмя видами [4]. Цель настоящей работы — изучить особенности фоторецепторного аппарата Я. lapicida, Задачи — оценить особенности клеточного состава фоторецепторного аппарата; описать морфологические особенности фоторецепторов; сравнить оптическую чувствительность разных типов фоторецепторов; определить возможную причину разнообразия фоторецепторов в сетчатке моллюска.
Материал и методы. Материалом для исследования служили взрослые особи наземного брюхоногого легочного моллюска Helicigona lapicida Linnaeus, 1758 (Helicidae, Ariantinae) с высотой раковины 6—8 и диаметром 17 — 19 мм. Моллюсков собирали на острове Эланд (Швеция) в июне 2004 г. Животных содержали в террариумах с почвой при температуре 18—20 ’С и естественном световом режиме. Кормили листьями одуванчиков.
Для световой и электронной микроскопии использовали глазные стебельки моллюсков, адаптированных к темноте, которые фиксировали в 2,5 % растворе глутараль-
дегида на ОД М какодилатном буфере (pH = = 8,0) в течение 2 ч при 4 °C. После отмывания от глутаральдегида в нескольких порциях буфера препараты фиксировали в 1 % растворе OsO4 на 0,1 М какодилатном буфере (pH = 8,0) в течение 1 ч при 4 °C. Обезвоживание проводили в серии этилового спирта возрастающей концентрации. Заливали материал эпоном-812. Полутонкие и ультратон кие срезы нарезали стеклянными ножами при помощи ультрамикротома V LKB 2088. Для световой микроскопии полутонкие срезы (2 мкм) окрашивали 0,5 % раствором толуидинового синего с добавлением 1 % карбоната натрия и просматривали при помощи светового микроскопа Zeiss Axiophot. Фотографии получали при помощи цифровой камеры Olympus DP 50. Для электронной микроскопии ультратон-кие срезы (70 нм) последовательно окрашивали 2 % раствором уранилацетата и 0,1 % раствором цитрата свинца и изучали при помощи трансмиссионного электронного микроскопа JEM-1230. Морфометрию выполняли на фотографиях срезов при помощи компьютерной программы CorelDraw 10.
Для расчета оптической чувствительности фоторецепторов к белому свету (S£, мкм2 • ср"1) использовали формулу [20]:
Sr = (—) х Л х (—) х---------
5 4 f (2,3 +AD где А — диаметр апертуры глаза, составляющий 103 ± 3,0 мкм [4], d —диаметр фоторецептора (табл. 1), f — фокусное расстояние оптической системы глаза, равное 162 мкм [4], к — коэффициент экстинкции фотопигмента у ракообразных in situ, равный 0,0067 мкм"’ [7], И / — длина фоторецептора, соответствующая длине микровил-лярного слоя (см. табл. 1).
Для расчетов использовали средние значения полученных данных. В подписях к формуле и табл. 1 эти значения приведены со стандартным отклонением [2].
Таблица 1
Параметры фоторецепторов центральной области сетчатки глаза Heticigona lapicida
|
Параметр |
Фр I |
Фр П-1 |
Фр П-2 |
Фр II 3 |
|
Диаметр фоторецептора d, мкм |
8,0 ± 0,9 (п = 75) |
5,0 ± 0,2 (п = 29) |
2,5 ± 0,1 (п = 25) |
2,0 ± 0,1 (я = 25) |
|
Длина фоторецептора 1, мкм |
11,0 ± 1,0 (п = 90) |
2,3 ±0,1 (п = 19) |
1,9 ±0,1 (п = 15) |
2,2 ± 0,1 (п = 15) |
|
Толщина микровилл, мкм |
0,07 ± 0,006 (п = 20) |
0,1 ± 0,03 (п = 30) |
||
|
Оптическая чувствительность фоторецепторов 5£| мкм2 ■ ср-1 |
0,5 |
0,5 |
0,008 |
0,006 |
Примечание. Фр I — фоторецептор первого типа, Фр II-1 — фоторецептор второго типа первого вида, Фр П-2 — фоторецептор второго типа второго вида, Фр П-З — фоторецептор второго типа третьего вида. Все параметры взяты из [4].
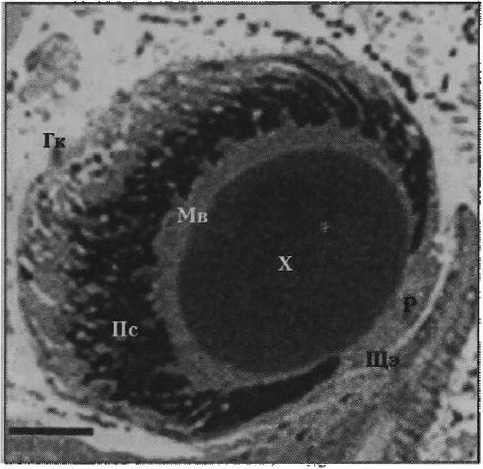
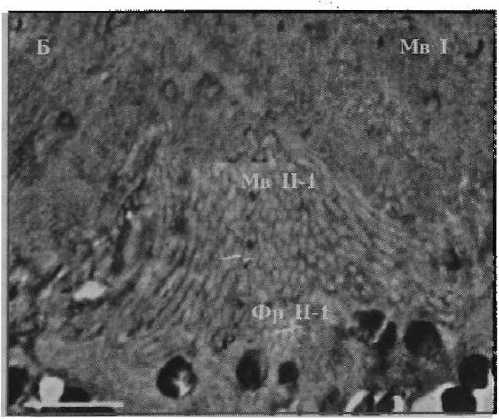
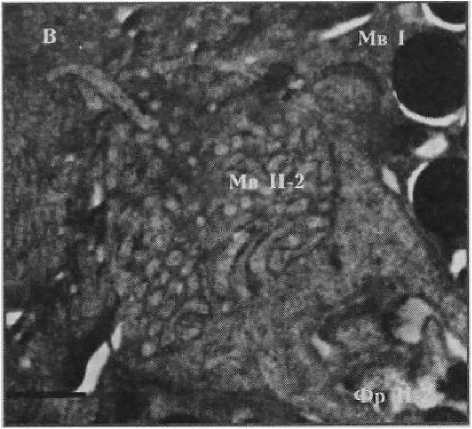
Результаты. Ранее было показано [4], что Н. lapicida имеет камерные глаза с роговицей, хрусталиком, стекловидным телом и неинвертированной сетчаткой, в которой можно выделить микровил лярные фоторецепторы первого и второго типа (рис. 1). Фоторецепторы первого типа — клетки с сильновыпуклой апикальной поверхностью, от которой отходят длинные тонкие микровиллы (рис. 2, а). Центральные микровиллы направляются вверх к хрусталику, латеральные контактируют с микровиллами соседних фоторецепторов первого типа, покрывают соседние фоторецепторы второго типа и пигментные клетки. Фоторецепторы второго типа представлены тремя видами, которые различаются между собой диаметром дистальной части тела клетки, степенью ее вогнутости и длиной микровилл. Первый вид характеризуется самой широкой и лишь слегка вогнутой апикальной поверхностью, а также самыми длинными микровиллами (рис. 2, б). У второго вида отмечается более узкая и более вогнутая верхняя часть тела клетки с микровиллами меньшей длины (рис. 2, в). Для третьего вида характерна еще более узкая и еще более вогнутая апикальная поверхность с микровиллами немного большей длины (рис. 2, г). Толщина микровилл у всех видов фоторецепторов второго типа одинаковая и немного больше толщины микровилл фоторецепторов первого типа (см. табл. 1; табл. 2). Микровиллы фоторецепторных клеток обоих типов образуют микровиллярный, или светочувствительный, слой сетчатки, основным компонен- том которого являются микровиллы фоторецепторов первого типа. Длина микровил-лярного слоя уменьшается в направлении от центральной области сетчатки к периферической — зрачку. В этой части сетчатки фоторецепторные клетки отсутствуют.
Рисунок 1
Фотография продольного полутонкого среза глаза Helicigona lapicida [4]
Масштаб: 45 мкм
Щэ — щупальцевый эпидермис
Р — роговица
X — хрусталик
Мв — микровиллярный слой сетчатки
Пг — пигментный слой сетчатки
Гк — глазная капсула
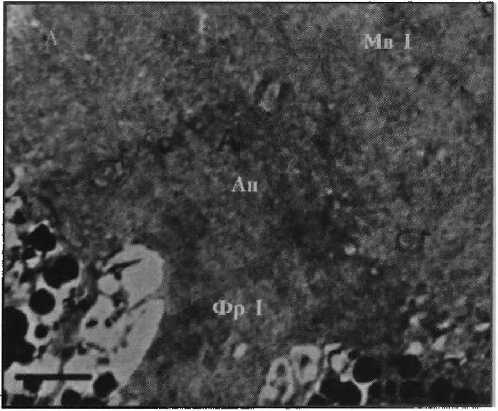
Фоторецептор первого типа
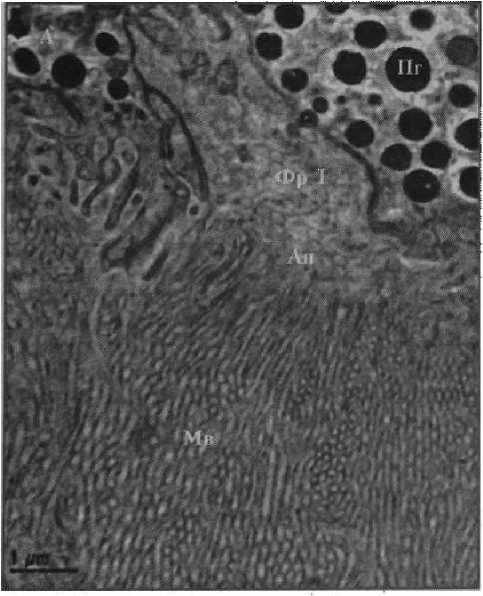
Рисунок 2
Фотографии фоторецепторов центральной сетчатки глаза Helicigona lapicida [4] Масштаб: / лекл (А, Б) и 0,5 мкм (В, Г)
Фр I — фоторецептор первого типа;
Фр П-1 — фоторецептор второго типа первого вида
Фр П-2 — фоторецептор второго типа второго вида
Фр П-3 — фоторецептор второго типа третьего вида
Мв 1 — микровиллы фоторецептора первого типа
Мв П-1 — микровиллы фоторецептора второго типа первого вида
Мв П-2 — микровиллы фоторецептора второго типа второго вида
Мв П-3 — микровиллы фоторецептора второго типа третьего вида
Ап — апикальная поверхность фоторецептора
Фоторецептор второго типа первого вида
Фоторецептор второго типа второго вида
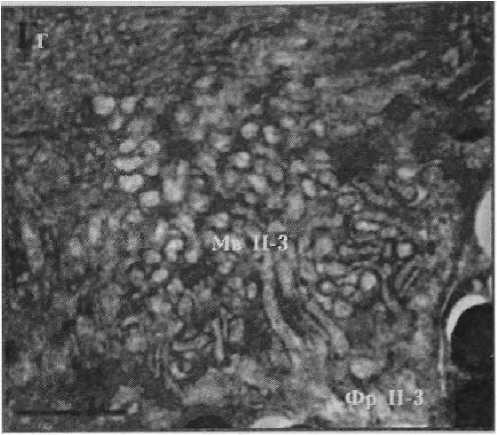
Фоторецемтор второго типа третьего вида
, Рисунок 2 ,
Фотографии фоторецепторов центральной сетчатки глаза Helicigona lapicida [4] Масштаб! 1 мкм (Л, Б) и 0,5 мкм (В, Г)
Фр Г — фоторецептор первого типа
Фр П-1 — фоторецептор второго типа первого вида
Фр 11-2 — фоторецептор второго типа второго вида
Фр П-З — фоторецептор второго типа третьего вида
Mb I — микровиллы фоторецептора первого типа
Мв II-1 — микровиллы фоторецептора второго типа первого вида
Мв П-2 — микровиллы фоторецептора второго типа второго вада
Мв П-З — микровиллы фоторецептора второго типа третьего вида
Ап — апикальная поверхность фоторецептора
Таблица 2
Сравнительная характеристика фоторецепторов сетчатки глаза Helicigona lapicida
|
Фоторецептор |
Апикальная поверхность |
Микровиллы |
||
|
Первого типа |
Широкая |
Сильновыпуклая, наподобие шляпки гриба |
Длинные |
Тонкие |
|
Второго типа |
Узкая |
Вогнутая |
Короткие |
Толстые |
|
Фоторецепторы второго типа |
||||
|
Первого вида |
Самая широкая |
Слабовогнутая |
Самые длинные |
Одинаковой толщины |
|
Второго вида |
Средней ширины |
Средневогнутая |
Самые короткие |
|
|
Третьего вада |
Самая узкая |
С ильновогнутая |
Средней длины |
|
Обсуждение. Электронно-микроскопические исследования камерных глаз брюхоногих моллюсков, принадлежащих к разным филогенетическим группам, позволили сформировать представление об их классическом микровиллярном фоторецепторном аппарате. Оказалось, что он может состоять из фоторецепторов первого типа, представленных одним видом, или из фоторецепторов первого и второго типа, каждый из которых представлен также одним видом. Примечательно, что один тип фоторецепторов встречается преимущественно у первично-водных морских и пресноводных жабер- пых видов, а два типа — у наземных легочных и вторично-водных пресноводных легочных видов [6; 8—10; 13; 17-19]. В сетчатке морского заднежаберного моллюска Bulla gouldiana Pilsbry, 1895 [14] и наземного легочного моллюска, объекта настоящего исследования, Н. lapicida [4; 6] наблюдается исключительное разнообразие микровиллирных фоторецепторов. Сетчатка В. gouldiana содержит три вида микровил-лярных фоторецепторов, два из которых соответствуют фоторецепторам первого типа, а один — фоторецепторам второго типа. Морфологические различия между двумя видами фоторецепторов первого типа выражаются в форме и диаметре апикального отростка, несущего микровиллы, и диаметре микровилл. У одного вида клеток апикальный отросток колбообразный, с толстыми микровиллами, у другого вида — пальцевидный, меньшего диаметра и с менее толстыми микровиллами. Фоторецепторы вто- рого типа — маленькие клетки с немного вогнутой дистальной поверхностью и короткими тонкими микровиллами [14]. Таким образом, фоторецепторный аппарат В, gouldiana, состоящий из трех видов микровил-
лярных фоторецепторов, является уникальным для брюхоногих моллюсков, особенно для заднежаберных, которым свойствен только один вид клеток. Фоторецепторный аппарат Н. lapicida уникален по-своему: он образован четырьмя видами микровилляр-ных фоторецепторов — одним видом фоторецепторов первого типа и тремя видами фоторецепторов второго типа [4; 6]. В отличие от В. gouldiana у Н. lapicida дополнительные виды клеток присутствуют среди фоторецепторов второго типа, а не первого. Морфология фоторецепторов первого и второго типа у Н. lapicida в целом подходит под общепринятое описание этих типов клеток у брюхоногих моллюсков. Однако апикальная поверхность фоторецепторов первого типа особенная, потому что имеет выпуклость наподобие шляпки гриба (см. рис. 2, а). Как правило, апикальная поверхность фоторецепторов первого типа имеет полукруглую или коническую выпуклость разной степени выраженности или отросток разной формы и длины (рис. 3) [6; 19; 8; 10; 13; 17]. Длина апикального отростка в центральной сетчатке обычно варьируется от 9 мкм, как у пресноводного легочного
Наземный слизень Arion rufus [6]
Рисунок 3 _
Фотографии фоторецепторов первого типа центральной области сетчатки глаз брюхоногих моллюсков. Масштаб: 1 мкм
Фр I и Ph I — фоторецептор; Мв и Mv — микровиллы фоторецептора; Ап и Ар — апикальная поверхность фоторецептора; Пг и Pg — пигментные гранулы
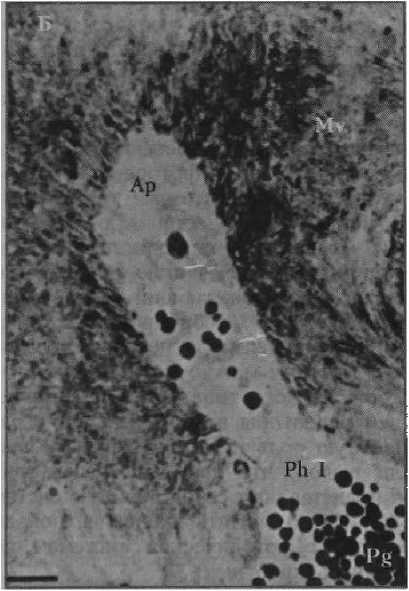
Пресноводный легочный моллюск Lymnaea stagnalis [10]
моллюска Planorbarius corneas Linnaeus, 1758, до 71 мкм, как у пресноводного легочного моллюска Lymnaea stagnates Linnaeus, 1758 [19]. У наземного слизня Athoracophorus bitentacuiatus Quoy, Gaimard, 1832 обнаружены необыкновенно длинные рецепторные отростки (около 100 мкм), несущие тысячи микровилл [11]. Таким образом, среди исследованных видов брюхоногих моллюсков A. bitentacuiatus и Н. lapicida — обладатели фоторецепторов первого типа с необычной организацией. Фоторецепторы второго типа у Н. lapicida не выделяются какими-либо структурными особенностями (см. рис. 2, б—гХ Тем не менее эти клетки различаются теми параметрами, которые определяют их чувствительность к свету (см. табл. 1). Очевидно, что сетчатка наземного брюхоногого легочного моллюска Н. lapicida отличается от сетчаток изученных видов брюхоногих моллюсков как клеточным составом фоторецепторного аппарата, так и морфологией фоторецепторов первого типа.
В 1979 г. Сузуки и соавторы [9] при помощи электрофизиологических опытов у наземного слизня Limax flavus Linnaeus, 1758 выявили функциональные различия между фоторецепторами первого и второго типа: фоторецепторы первого типа оказались более чувствительными к свету, чем фоторецепторы второго типа. В глазу L. flavus пространство между хрусталиком и сетчаткой почти полностью занято плотной массой микровилл фоторецепторов первого типа. Фоторецепторы второго типа имеют меньшее количество коротких микровилл. Поскольку микровиллы являются световоспринимающей частью фоторецепторных клеток, свет, попадающий в глаз, должен более эффективно абсорбироваться микровиллами фоторецепторов первого типа, чем второго. Поэтому можно предположить, что рецепторы первого типа более чувствительные, чем рецепторы второго типа, и играют доминантную роль в фоторецепции. Ученые сделали вывод, что два типа рецепторных клеток представляют собой двойную систему, которую можно считать эквивалентной ско-топической и фотопической системам глаз позвоночных животных.
Позднее для брюхоногих моллюсков из разных мест обитания при помощи вычислительных методов [20] было показано, что оптическая чувствительность фоторецепторов первого типа превосходит чувствительность фоторецепторов второго типа в десятки, а иногда и в сотни раз [3; 5; 6]. У Н. lapicida чувствительность фоторецепто ров первого типа (0,5 мкм2 - ср') выше чувствительности фоторецепторов второго типа (0,04, 0,008, 0,006 мкм2 ■ ср 9 в 12,5, 62,5 и 83,3 раза соответственно. Увеличение чувствительности к свету фоторецепторов первого типа по сравнению с фоторецепторами второго типа достигается за счет удлинения световоспринимающих частей в 4,8, 5,8, 5,0 раза и увеличения их диаметра в 1,6, 3,2, 4,0 раза соответственно. Варьирование чувствительности фоторецепторов второго типа обусловлено разным диаметром их дистальной поверхности и разной длиной микровилл (см. табл. 1). Таким образом, чувствительность фоторецепторов первого типа возрастает в основном за счет длины микровилл, а чувствительность фоторецепторов второго типа меняется чаще всего за счет диаметра поверхности, несущей микровиллы. Так как чувствительность всех видов фоторецепторных клеток ниже единицы, по классификации Лэнда и Нильссона [16] эти клетки предназначены для функционирования при дневном свете. Это заключение соответствует типу активности Н. lapicida, которая обитает на скалах, прячась в расщелинах, и появляется на поверхности днем [15]. Однако существенные различия в чувствительности рецепторных клеток позволяют полагать, что более чувствительные фоторецепторы первого типа могут работать при более низком, а менее чувствительные фоторецепторы второго типа — при более высоком уровне освещенности [4; 6]. Оптическая чувствительность фоторецепторов первого и второго типа у Н. lapicida (0,5/0,04, 0,008, 0,006 мкм2-ср9 сравнима с чувствительностью фоторецепторов дневных брюхоногих моллюсков: морского прибрежного переднежаберного моллюска Littorina littorea Linnaeus, 1758 (0,4 мкм2 ср'1) [20], пресноводного легочного моллюска L. stagnates (0,04 мкм2 ср-9, выбирающего хорошо освещенные прибрежные участки мелких прудов [12], наземных легочных моллюсков Cochlodina laminata Montagu, 1803 (0,3 мкм2 - ср-1), живущего на стволах деревьев в лесах, Рег-foratella incamata Muller, 1774 (0,07 мкм2х х ср"9, предпочитающего участки с редкой невысокой растительностью под деревьями в лесах [3], Arianta arbustorum Linnaeus, 1758 (0,73/0,002 мкм2 ср'9 и Сераса hortensis Muller, 1774 (0,65/0,02 мкм2 - ср*1), обитающих в зарослях крапивы на полях [5].
Почему же у Н. lapicida возникло такое разнообразие микровиллярных фоторецепторных клеток? Известно, что наличие в сетчатке нескольких типов фоторецепторов, чувствительных к световым волнам разной длины, — признак цветового зрения. Оно улучшает различимость объектов и обеспечивает дополнительную информацию о них, расширяет возможности животного добывать пищу и избегать врагов [1]. Спектральная чувствительность глаза Н. lapicida не исследована, что не позволяет сделать вывод о наличии или отсутствии цветового зрения у этого моллюска. Однако из тех немногих видов наземных брюхоногих моллюсков, для которых определены зрительные способности, только Н. lapicida предпочитает открытые участки на скалистом берегу рядом с морем стволам деревьев, зарослям кустарников и травы. Глаз Н. 1а picida приспособлен к зрению в дневное время [4; 6]. Возможно, на открытом пространстве реализация тех или иных форм поведения требует более подробной информации об окружающих объектах. Поэтому наиболее вероятным предположением для объяснения возникновения разнообразия фоторецепторов в сетчатке Я. lapicida следует считать способность моллюска различать длины волн света.
Заключение. Исследования сетчатки камерного глаза наземного брюхоногого легочного моллюска Н. lapicida показали, что среди изученных видов брюхоногих моллюсков его фоторецепторный аппарат является уникальным как по клеточному составу, так и по морфологии фоторецепторов первого типа. Присутствие нескольких видов фоторецепторов, возможно, означает наличие цветового зрения у моллюска.
Благодарность. Автор благодарит профессора Э. Дж. Ворранта за предоставленную возможность проведения исследований в Университете г. Лунда, М. Соренссо-на за доставку моллюсков с острова Эланд и заведующую лабораторией Р. Валлен за помощь в изготовлении ультратонких срезов.
Список литературы Разнообразие и особенности фоторецепторов в сетчатке камерного глаза брюхоногого моллюска Helicigona lapicida Linnaeus, 1758 (Pulmonata, Stylommatophora)
- Гиляров М. С. Биологический энциклопедический словарь/М. С. Гиляров; под. ред. М. С. Гилярова. -2-е изд., испр. -М.: Большая рос. энцикл., 1995. -С. 698.
- Лакин Г. Ф. Биометрия/Г. Ф. Лакин. -М.: Высш. шк., 1990. -351 с.
- Шепелева И. П. Сравнительное изучение морфологии и оптики глаз наземных брюхоногих моллюсков Cochlodina laminata и Perforatella incarnata (Pulmonata: Stylommatophora) с различными световыми предпочтениями/И. П. Шепелева//Сенсор. системы. -2006. -Т. 20, № 1. -С. 40-51.
- Шепелева И. П. Глаз наземного брюхоногого моллюска Helicigona lapicida (Pulmonata: Stylommatophora)/И. П. Шепелева//Сенсор. системы. -2006. -Т. 20, № 1. -С. 52-58.
- Шепелева И. П. Сравнительное изучение морфологии и оптики глаз наземных брюхоногих моллюсков Arianta arbustorum и Cepaea hortensis (Pulmonata: Stylommatophora) со сходными световыми предпочтениями/И. П. Шепелева//Сенсор. системы. -2006. -Т. 20, № 1. -С. 59-67.
- Шепелева И. П. Сравнительная морфология и оптические свойства глаз брюхоногих моллюсков Stylommatophora (Gastropoda: Pulmonata): дис. … канд. биол. наук/И. П. Шепелева. -М., 2007. -147 с.
- Bruno M. S. The visual pigment and visual cycle in the lobster Homarus/M. S. Bruno, S. N. Barnes, T. H. Goldsmith//Journ. of Comparative Physiology. -1977. -№ 8. -P. 123-142.
- Charles J. H. Sense organs (less cephalopods)/J. H. Charles//Physiology of Molluscs. -1966. -V. 2, № 4. -P. 455-521.
- Duplex system in the simple retina of a gastropod mollusc, Limax flavus L./H. Suzuki, M. Watanabe, Y. Tsukahara [et al.]//Jou rn. of Comparative Physiology. -1979. -№ 133. -P. 125-130.
- Eakin R. Evolution of photoreceptors/R. Eakin//Evolutionary Biology. -1968. -№ 2. -P. 194-242.
- Eakin R. M. Fine structure of the eye of the New Zealand slug Athoracophorus bitentaculatus/R. M. Eakin, J. L. Brandenburger, J. M. Barker//Zoomorphology. -1980. -№ 94. -P. 225-239.
- Fixed focal-length optics in pulmonate snails (Mollusca, Gastropoda): squaring phylogenetic background and ecophysiological needs (II)/J. Gal, M. V. Bobkova, V. V. Zhukov [et al.]//Jou rn. of Invertebrate Biology. -2004. -V. 123, № 2. -P. 116-127.
- Hughes H. P. I. A light and electron microscopy study of some opisthobranch eyes/H. P. I. Hughes//Zeitschrift fur Zellforschung und Mikroskopische Anatomie. -1970. -№ 106. -P. 79-98.
- Jacklet J. W. Ultrastructure of photoreceptors and circadian pacemaker neu rons in the eye of a gastropod, Bulla/J. W. Jacklet, W. Colquhoun//Jou rn. of Neurocytology. -1983. -№ 12. -P. 673-696.
- Kerney M. P. A field guide to the land snails of Britain and north-west Europe/M. P. Kerney, R. A. D. Cameron. -L.: Collins St. Jame's Place, 1979. -288 p.
- Land M. F. Animal eyes/M. F. Land, D.-E. Nilsson. -Oxford: Oxford University Press, 2002. -P. 47-54.
- Messenger J. B. Comparative physiology of vision in molluscs/J. B. Messenger. -Berlin-Heidelberg; New York: Springer-Verlag, 1981. -P. 93-200.
- Seyer J.-O. Comparative optics of prosobranch eyes: PhD thesis/J.-O.Seyer. -Lund, 1998. -86 p.
- Variations in the retinal design of pulmonate snails (Mollusca, Gastropoda): squaring phylogenetic background and ecophysiological needs (I)/M. V. Bobkova, J. Gal, V. V. Zhukov [et al.]//Jou rn. of Invertebrate Biology. -2004. -V. 123, № 2. -P. 101-115.
- Warrant E. J. Absorption of white light in photoreceptors/E. J. Warrant, D.-E. Nilsson//Vision Research. -1998. -V. 38, № 2. -P. 195-207.
