Разнообразие митохондриальной ДНК сибирских татар из позднесредневекового могильника Абрамово-10 (Барабинская лесостепь)
")
Автор: Черданцев С.В., Трапезов Р.О., Томилин М.А., Федосеева Е.И., Журавлев А.А., Пилипенко И.В., Рыкун М.П., Молодин В.И., Пилипенко А.С.
Журнал: Вестник Новосибирского государственного университета. Серия: История, филология @historyphilology
Рубрика: Археология Евразии
Статья в выпуске: 7 т.24, 2025 года.
Бесплатный доступ
Представлены первые результаты палеогенетического исследования позднесредневекового населения западносибирской лесостепной зоны. Получены данные о структуре 72 образцов митохондриальной ДНК из могильника сибирских татар Абрамово-10 (XVI–XVIII вв. н. э.), расположенного в Центральной Барабе. Проведен филогенетический и филогеографический анализ исследованной серии. Полученные результаты подтверждают участие компонентов тюркского, угорского и самодийского происхождения в формировании генофонда исследуемой популяции. Выявлено сходство серии из Абрамово-10 с современными группами сибирских татар, особенно тоболо-иртышскими татарами. Высказано предположение о возможности изменения ареалов отдельных групп сибирских татар в позднем Средневековье и Новом времени. Как минимум часть тюркоязычного коренного населения сохранила структуру генофонда митохондриальной ДНК, несмотря на влияние пришлого европеоидного населения.
Сибирские татары, Барабинская лесостепь, позднее Средневековье, палеогенетика, митохондриальная ДНК, этногенетические реконструкции
Короткий адрес: https://sciup.org/147251712
IDR: 147251712 | УДК: 572, 575 | DOI: 10.25205/1818-7919-2025-24-7-132-149
Mitochondrial DNA Diversity in Siberian Tatars from the Late Medieval Burial Ground Abramovo-10 (Baraba Forest-Steppe)
Purpose. Although the medieval population of West Siberian forest-steppe has been studied for a long time by methods of archeology and physical anthropology, paleogenetic data on these groups are still completely absent. We present the first paleogenetic results for late medieval populations of the forest-steppe zone, including the analysis of a mitochondrial DNA series from the Abramovo-10 burial ground (16th – 18th centuries AD, Baraba forest-steppe), and consider these paleogenetic results in the context of the interdisciplinary data on ethnogenetic processes in the region. Results. We analyzed the structure, phylogeny, and phylogeographic features of 72 mitochondrial DNA samples from the Abramovo-10 burial ground. As a result, we obtained a well-representative sample of mtDNA for the late medieval population of Central Baraba. The population shows similarities in mtDNA diversity with modern Siberian Tatars (closer to the Tobolo-Irtysh than to the modern Baraba local group), as well as Ugric and Samoyed indigenous populations of Western Siberia. Conclusion. Our results confirm the participation of genetic components of Turkic (both from the Eurasian steppes and the Altai-Sayan region), Ugric, and Samoyed origin in the formation of the late medieval population gene pool of in West Siberian forest-steppe zone. We assume that sub-ethnic groups of Siberian Tatars could change their settlement territories in the late Middle Ages and Modern times. At least a part of the Turkic-speaking indigenous population in the West Siberian forest-steppe retained the structure of the mitochondrial DNA gene pool, despite the influence of the migrant Caucasian population.
Текст научной статьи Разнообразие митохондриальной ДНК сибирских татар из позднесредневекового могильника Абрамово-10 (Барабинская лесостепь)
В основе этногенетических процессов в лесостепной и южно-таежной зонах Западной Сибири в Средневековье лежало взаимодействие автохтонных для региона протоугорских и протосамодийских групп с пришлыми тюркскими популяциями, что отразилось на материальной культуре, языках и антропологическом составе популяций [Молодин и др., 1990; Соболев, 2008]. Результатом стало формирование основных этнических групп, составляющих современное коренное население региона. В лесостепи происходит сложение тюркоязычных групп, которые, как правило, объединяются специалистами под общим названием «сибирские татары» [Валеев, 1993]. Завершающие этапы этих этногенетических процессов приходятся на XIII–XIV вв. [Соболев, 2008]. Для последующего периода позднего Средневековья археологами открыты и исследованы многочисленные поселенческие и погребальные памятники, для которых существует возможность этнической атрибуции. Одним из наиболее исследованных в данном отношении районов западносибирской лесостепной зоны является Барабинская лесостепь. Центральная, собственно лесостепная, зона Барабы является ареалом коренного тюркоязычного населения – барабинских татар, или барабинцев. Наряду с тоболо-иртышскими и томскими татарами они рассматриваются большинством специалистов в качестве одной из основных локально-территориальных групп сибирских татар [Томилов, 1992; Валеев, 1993; Валеев, Томилов, 1996]. Следует отметить, что некоторые исследователи выводят барабинцев из общности сибирских татар, считая их самостоятельным этносом [Токарев, 1958; Селезнев, 1994].
Среди археологических памятников, связываемых с барабинцами, особое место занимает могильник Абрамово-10 (Куйбышевский район Новосибирской области) – самый большой полностью раскопанный позднесредневековый могильник в Центральной Барабе. На нем исследовано 118 погребений XVI–XVIII вв. [Молодин и др., 1990; Соболев, 2008]. Важнейшее значение для этнокультурных реконструкций имеет краниологическая серия из Абрамово-10, насчитывающая почти 80 индивидов. Большая численность серии и высокий уровень сохранности черепов (и зубов, в частности) обеспечивают перспективность палеогенетического исследования данных материалов. Публикацией первых палеогенетических результатов для могильника Абрамово-10 мы открываем цикл работ о генетическом составе позднесредневековых популяций Западной Сибири. В данной работе мы представляем результаты анализа серии образцов митохондриальной ДНК ( N = 72) из этого памятника и интерпретируем полученные данные в контексте имеющихся представлений об этногенетических процессах в лесостепной зоне Западной Сибири в позднем Средневековье.
Материалы и методы
Палеоантропологические материалы, исследованные в работе. Для проведения работы были получены образцы от 74 индивидов из позднесредневековых погребений могильника Абрамово-10 (табл. 1). Основная часть черепов из этого памятника хранится в составе краниологической коллекции кабинета антропологии ТГУ. Из нее были отобраны образцы для 73 из 74 индивидов, исследованных в нашей работе (инвентарные номера краниологической коллекции кабинета антропологии ТГУ № 3931–4004). Отбор проводился совместно палеогенетиками ИЦиГ СО РАН (А. С. Пилипенко, А. А. Журавлев) и палеоантропологом (М. П. Ры- кун). Образцы для одного индивида из могильника Абрамово-10 получены из палеоантропологической коллекции ИАЭТ СО РАН (шифр индивида 32/18Б).
Таблица 1
Выборка палеоантропологических образцов из позднесредневекового могильника Абрамово-10, исследованная в рамках данной работы
Description of paleoanthropological samples from the Late Medieval burial ground Abramovo-10 analyzed in this work
Table 1
|
№ п/п |
Инвентарный номер |
Лабораторный шифр индивида |
№ п/п |
Инвентарный номер |
Лабораторный шифр индивида |
№ п/п |
Инвентарный номер |
Лабораторный шифр индивида |
|
1 |
3934 |
Abr1 |
26 |
3965 |
Abr26 |
51 |
3931 |
Abr51 |
|
2 |
3982 |
Abr2 |
27 |
3959 |
Abr27 |
52 |
3933 |
Abr52 |
|
3 |
3932 |
Abr3 |
28 |
3953 |
Abr28 |
53 |
3962 |
Abr53 |
|
4 |
3954 |
Abr4 |
29 |
3987 |
Abr29 |
54 |
3996 |
Abr54 |
|
5 |
3956 |
Abr5 |
30 |
3978 |
Abr30 |
55 |
3944 |
Abr55 |
|
6 |
3973 |
Abr6 |
31 |
3977 |
Abr31 |
56 |
3936 |
Abr56 |
|
7 |
3975 |
Abr7 |
32 |
3942 |
Abr32 |
57 |
3945 |
Abr57 |
|
8 |
3946 |
Abr8 |
33 |
3972 |
Abr33 |
58 |
3989 |
Abr58 |
|
9 |
3948 |
Abr9 |
34 |
3955 |
Abr34 |
59 |
3957 |
Abr59 |
|
10 |
3966 |
Abr10 |
35 |
4002 |
Abr35 |
60 |
4004 |
Abr60 |
|
11 |
3943 |
Abr11 |
36 |
3990 |
Abr36 |
61 |
3952 |
Abr61 |
|
12 |
3979 |
Abr12 |
37 |
3983 |
Abr37 |
62 |
4003 |
Abr62 |
|
13 |
4000 |
Abr13 |
38 |
3975 |
Abr38 |
63 |
3967 |
Abr63 |
|
14 |
3974 |
Abr14 |
39 |
3997 |
Abr39 |
64 |
3969 |
Abr64 |
|
15 |
3986 |
Abr15 |
40 |
3940 |
Abr40 |
65 |
3984 |
Abr65 |
|
16 |
3981 |
Abr16 |
41 |
4001 |
Abr41 |
66 |
3995 |
Abr66 |
|
17 |
3993 |
Abr17 |
42 |
3958 |
Abr42 |
67 |
3939 |
Abr67 |
|
18 |
3992 |
Abr18 |
43 |
3970 |
Abr43 |
68 |
3950/1 |
Abr68 |
|
19 |
3961 |
Abr19 |
44 |
3960 |
Abr44 |
69 |
3980 |
Abr69 |
|
20 |
3994 |
Abr20 |
45 |
3968 |
Abr45 |
70 |
3047 |
Abr70 |
|
21 |
3937 |
Abr21 |
46 |
3964 |
Abr46 |
71 |
3941 |
Abr71 |
|
22 |
3950/2 |
Abr22 |
47 |
3999 |
Abr47 |
72 |
3051 |
Abr72 |
|
23 |
3963 |
Abr23 |
48 |
3988 |
Abr48 |
73 |
3971 |
Abr73 |
|
24 |
3935 |
Abr24 |
49 |
3998 |
Abr49 |
74 |
32/18Б |
Abr74 |
|
25 |
3949 |
Abr25 |
50 |
3991 |
Abr50 |
Во всех случаях в качестве образцов были отобраны зубы высокой степени макроскопической сохранности.
Предварительную обработку палеоантропологического материала и получение образцов суммарной ДНК осуществляли с помощью методов, детально описанных в наших предшествующих работах [Pilipenko et al., 2015; 2018]. Поверхность зуба механически очищали от загрязнений, выдерживали зуб в растворе гипохлорита натрия, облучали ультрафиолетом и размалывали с помощью шаровой мельницы Retsch Mixer Mill MM200 (Германия).
Материал зубов декальцинировали с помощью 0,5М ЭДТА с последующим лизисом протеиназой K (Сибэнзим, Россия). ДНК выделяли методом фенол-хлороформной экстракции.
Анализ структуры мтДНК включал определение последовательности первого гипервариабельного сегмента контрольного района мтДНК (ГВСI мтДНК). Амплификацию ГВСI мтДНК проводили параллельно двумя методами: 1) в виде четырех коротких перекрывающихся фрагментов [Haak et al., 2005]; 2) в виде одного длинного фрагмента с помощью «вложенной» ПЦР [Пилипенко и др., 2008]. Для части образцов с помощью ПЦР-систем, описанных в работе [Wilde et al., 2014], определяли статус филогенетически информативных позиций в кодирующей части мтДНК.
Последовательности нуклеотидов определяли с использованием набора реактивов ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit v.3.1 (Thermo Scientific, СШA). Продукты секвенирующей реакции анализировали на автоматическом капиллярном секвенаторе ABI Prism 3130XL Genetic Analyser (Applied Biosistems, США) в центре коллективного пользования «Геномика» СО РАН. Филогенетическую и филогеографическую интерпретацию результатов осуществляли методами, описанными нами ранее [Pilipenko et al., 2018].
Все работы с древним материалом выполнены на базе специализированной палеогенети-ческой инфраструктуры межинститутской лаборатории молекулярной палеогенетики и палеогеномики ИЦИГ СО РАН (Новосибирск, Россия), которая отвечает всем современным требованиям, предъявляемым к палеогенетическим лабораториям. Меры против контаминации и процедуры верификации результатов описаны нами ранее [Ibid.]. Условия проведения исследования, используемые протоколы работы и особенности экспериментальных результатов однозначно свидетельствуют о высокой достоверности полученных нами палеогенетиче-ских данных.
Результаты исследования
Для каждого из 74 индивидов было получено 2–3 экстракта суммарной ДНК. Серия из могильника Абрамово-10 продемонстрировала очень высокий уровень сохранности ДНК (в 72 из 74 образцов, т. е. более 97 %). Это может объясняться как поздней датировкой могильника, так и благоприятными условиями для сохранности ДНК в месте расположения памятника. Для всех 72 образцов была определена последовательность ГВСI мтДНК. На основании полученных данных о структуре гаплотипа ГВСI мтДНК был выполнен первичный филогенетический анализ, позволивший установить филогенетическое положение всех исследованных вариантов (табл. 2). Принадлежность к гаплогруппам для части индивидов была подтверждена с помощью анализа гаплогрупп-специфичных позиций в кодирующей части мтДНК.
При численности выборки N = 72 можно ожидать, что в ее составе зафиксированы все основные компоненты генофонда мтДНК группы населения, сформировавшей могильник Аб-рамово-10: с вероятностью 95 % в нашей выборке присутствуют все гаплогруппы мтДНК, частота которых в генофонде популяции составляла 4 % и выше [Ibid.].
В составе серии выявлено 24 структурных варианта мтДНК, которые относятся как к западно-евразийским (H, J, T*, T1a2a, U2e, U4, U5a), так и к восточно-евразийским (A10, C*, C5b1, D, F1b, M7, N9a, Z) гаплогруппам мтДНК. Западно-евразийский компонент доминирует по числу носителей (45 из 72 образцов, 62,5 %). Но на уровне разнообразия гаплотипов можно говорить о сопоставимой представленности компонентов западно-евразийского и восточно-евразийского происхождения в исследуемой серии: 13 восточно-евразийских, 11 западно-евразийских из 24 вариантов всего.
Наибольшую представленность и разнообразие вариантов в западно-евразийском кластере демонстрирует гаплогруппа H, а в восточно-евразийском – C и D. Каждая из них представлена 4 структурными вариантами. В этом отношении с ними контрастирует гаплогруппа U4,
Таблица 2
Структура и филогенетическая принадлежность вариантов мтДНК из могильника Абрамово-10
Structure and phylogenetic position of mtDNA lineages from the Abramovo-10 burial ground
Table 2
|
Номер структурного варианта мтДНК |
Носители варианта из могильника Абрамово-10 |
Структура гаплотипа ГВСI мтДНК |
Число носителей в серии |
Гаплогруппа мтДНК |
|
1 |
Abr12 |
16223T-16227C-16290T-16311C-16319A |
1 |
A10 |
|
2 |
Abr50, Abr68 |
16093C-16223T-16255A-16298C-16327T |
2 |
C* |
|
3 |
Abr18 |
16129A-16150T-16223T-16298C-16327T |
1 |
C4a1a4a |
|
4 |
Abr53 |
16223T-16291T-16298C-16327T |
1 |
C4b3 |
|
5 |
Abr6, Abr45, Abr46, Abr54, Abr55, Abr60, Abr70, Abr73, Abr74 |
16148T-16223T-16288C-16298C-16327T |
9 |
C5b1 |
|
6 |
Abr59, Abr64 |
16218T-16223T-16362C |
2 |
D |
|
7 |
Abr27, Abr38 |
16223T-16362C |
2 |
D |
|
8 |
Abr20, Abr36 |
16223T-16291T-16362C |
2 |
D |
|
9 |
Abr1, Abr39 |
16223T-16291T-16311C-16362C |
2 |
D |
|
10 |
Abr3 |
16172C-16179T-16183C-16189C- 16232A-16249-16304C-16311C |
1 |
F1b |
|
11 |
Abr2, Abr5, Abr8, Abr9, Abr21, Abr24, Abr42, Abr43, Abr58, Abr65, Abr66, Abr71 |
CRS |
12 |
H |
|
12 |
Abr63 |
16209C-16299G |
1 |
H |
Окончание табл. 2
|
Номер структурного варианта мтДНК |
Носители варианта из могильника Абрамово-10 |
Структура гаплотипа ГВСI мтДНК |
Число носителей в серии |
Гаплогруппа мтДНК |
|
13 |
Abr16, Abr33, Abr41, Abr61 |
16311C |
4 |
H |
|
14 |
Abr19 |
16362C |
1 |
H |
|
15 |
Abr26 |
16069T-16126C-16235G |
1 |
J |
|
16 |
Abr11, Abr32 |
16093C-16129A-16192T-223T-16297C- 16309G |
2 |
M7 |
|
17 |
Abr51 |
16172C-16189C-16209C-16223T- 16257A-16261T |
1 |
N9a |
|
18 |
Abr40, Abr57, Abr72 |
16126C-16294T |
3 |
T* |
|
19 |
Abr15, Abr47, Abr56, Abr67, Abr69 |
16126C-16163G-16186T-16189C- 16261T-16294T |
5 |
T1a2a |
|
20 |
Abr52 |
16051G-16129C-16189C-16256T-16362C |
1 |
U2e |
|
21 |
Abr4, Abr7, Abr10, Abr14, Abr17, Abr28, Abr29, Abr30, Abr31, Abr35, Abr44, Abr48, Abr49 |
16134T-16356C |
13 |
U4 |
|
22 |
Abr13 |
16256T-16270T-16294T-16360T |
1 |
U5a |
|
23 |
Abr23, Abr25, Abr37 |
16086С-16192T-16239T-16256T-16270T |
3 |
U5a |
|
24 |
Abr22 |
16185T-16223T-16260T-16298C |
1 |
Z |
Всего образцов 72
большое число носителей которой (13), представлены единственным гаплотипом 16134T-16356C (№ 21, здесь и далее номер гаплотипа указан по табл. 2). Большим числом носителей в исследованной серии представлены также варианты западно-евразийских гаплогрупп H (№ 11) и T1a2 (№ 19), среди восточно-евразийских – вариант гаплогруппы C5b1 (№ 5). При исследовании серии образцов из одного могильника существует вероятность попадания в выборку близких родственников. Присутствие большого числа родственников может искажать картину соотношения компонентов в исследуемой серии по сравнению с популяцией в целом. Для выявления индивидов, состоящих в близком родстве, в палеогенетике требуется проведение дополнительной экспериментальной работы (которая не входила в наши текущие задачи). Однако варианты, обнаруженные нами с высокой частотой в серии из Абрамово-10 (№ 21, 11, 5, 19), присутствуют также и в генофондах современных сибирских татар – бара-бинских [Gubina et al., 2018] и тоболо-иртышских [Naumova et al., 2008], при анализе которых все близкие родственники по материнской линии были исключены из состава выборок. В связи с этим мы полагаем, что высокая частота отдельных вариантов мтДНК, выявленная нами в серии из Абрамово-10, в значительной степени являлась характерной чертой генофонда позднесредневековых сибирских татар, а не следствием присутствия в серии родственников. Наблюдаемая нами повышенная представленность в исследуемой серии отдельных вариантов мтДНК может быть связана с высокой частотой эндогамных браков, которая была установлена этнографами для нескольких локальных групп барабинских татар [Томилов, 1992].
Вклад различных филогенетических кластеров мтДНК в состав исследованной серии из Абрамово-10 показан на рис. 1.
На основе полученных данных мы реконструировали филогенетическое дерево, отражающее эволюционные взаимоотношения всех выявленных вариантов мтДНК и наглядно демонстрирующее картину разнообразия мтДНК в серии из могильника Абрамово-10 (рис. 2).
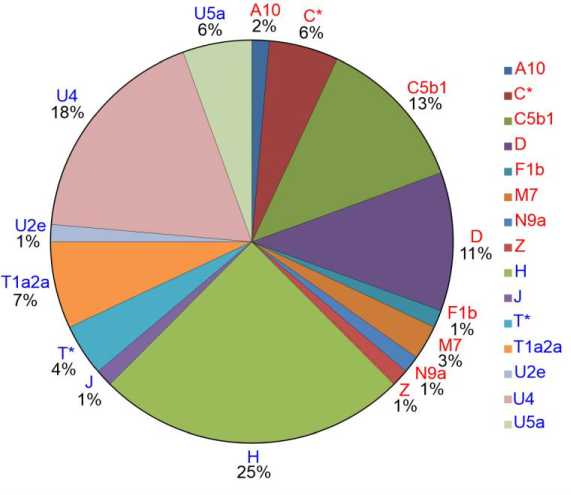
Рис. 1. Соотношение филогенетических кластеров мтДНК в исследованной серии из могильника Абрамово-10 ( N = 72). Названия гаплогрупп западно-евразийского кластера выделены синим цветом, названия восточно-евразийских гаплогрупп – красным
Fig. 1. Correlation of mtDNA phylogenetic clusters in the studied series from the Abramovo-10 burial ground ( N = 72). The names of Western Eurasian haplogroups shown in blue, the names of the Eastern Eurasian haplogroups shown in red
Т1а2а
F1b
/ 16£88 16093/
16232А
16*129
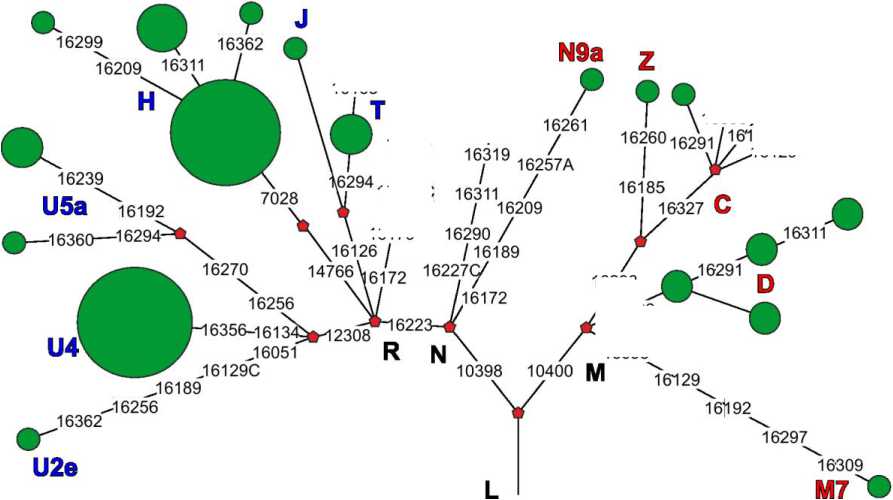
Рис. 2. Филогенетическое дерево структурных вариантов мтДНК, выявленных в серии из позднесредневекового могильника Абрамово-10. Названия гаплогрупп западно-евразийского кластера выделены синим цветом, названия восточно-евразийских гаплогрупп – красным
Fig. 2. Phylogenetic tree of mtDNA lineages identified in the series from the Late Medieval burial ground Abramovo-10. The names of Western Eurasian haplogroups shown in blue, the names of the Eastern Eurasian haplogroups shown in red
16183С

Важным инструментом интерпретации полученных нами данных о разнообразии мтДНК в серии из Абрамово-10 является ее сравнительный анализ с другими сериями, опубликованными в научной литературе. В отношении позднесредневековых популяций, которые имеют в той или иной степени уверенную этническую атрибуцию, сравнение с современными популяциями является корректным подходом для анализа палеогенетических данных. Исходя из особенностей этногенетических процессов в регионе, в качестве сравнительного материала для интерпретации полученных нами результатов мы выбрали литературные данные по популяциям, предки которых могли иметь отношение к интересующим нас позднесредневековым процессам: две группы современных сибирских татар – барабинские [Gubina et al., 2018] и тоболо-иртышские [Naumova et al., 2008], серию хантов [Губина и др., 2005] и селькупов [Tamm et al., 2007]. Эти серии позволяют нам охватить три основных этногенетических компонента (тюркский, угорский и самодийский), потенциально влиявших на процессы формирования генетического состава тюркоязычного позднесредневекового населения Барабин-ской лесостепи, в том числе и оставившей могильник Абрамово-10 группы населения. Необходимо отметить, что использованные нами в анализе серии современного угорского (ханты) и самодийского (селькупы) населения Западной Сибири происходят из их северных локальных популяций, проживающих на большом удалении от рассматриваемой нами лесостепной и южно-таежной зон. Эти серии могут не в полной мере отражать состав генофонда южных угров (например, хантов Васюганья) и южных самодийцев (нарымских селькупов), непосредственно контактировавших с позднесредневековыми тюркоязычными популяциями Барабы. Более корректным сравнительным материалом станут серии из могильников позднего Средневековья и Нового времени, относящихся к хантам и нарымским селькупам, а также серия носителей кыштовской культуры из памятников Северной Барабы, таких как позднесредневековый могильник Кыштовка-2, для населения которых предполагается южно-угорская этническая атрибуция [Молодин, 1979]. Анализ репрезентативных серий мтДНК от этих групп в настоящее время уже выполняется авторами.
Учитывая существующую дискуссию о потенциальной вовлеченности в процессы тюрки-зации западносибирского населения двух древнетюркских компонентов, связываемых со степными кимако-кыпчакскими группами (через распространение культур сросткинского круга) [Савинов, 1984; Молодин и др., 1988; 1990] и кругом тюркоязычных народов Алтае-Саянского региона [Селезнев, 1994], мы включили в предварительный сравнительный анализ еще ряд популяций: серии казахов с территории современного Казахстана и Алтая, хакасов, шорцев, ряд групп северных и южных алтайцев. Также в анализ включено несколько альтернативных выборок мтДНК русского населения. Современное русское население из различных регионов России может не отражать особенностей генетической структуры популяций, непосредственно контактировавших с коренными тюркскими популяциями Барабы позднего Средневековья и Нового времени. Корректнее было бы проводить сравнение с данными по русским из памятников финального Средневековья и Нового времени с территории Западной Сибири (их исследование осуществляется авторами). Но для предварительной стадии анализа достаточно использовать имеющиеся литературные данные о разнообразии мтДНК в генофондах современных локальных групп русских.
Сравнительный анализ с перечисленными современными популяциями проводили на нескольких уровнях: с помощью оценки межпопуляционного генетического расстояния по М. Слаткину [Slatkin, 1995], анализа общих гаплотипов мтДНК между популяциями, анализа филогеографических особенностей вариантов мтДНК из Абрамово-10. Исследованная нами позднесредневековая серия мтДНК демонстрирует наибольшее сходство с выборками из групп сибирских татар (тоболо-иртышских и барабинских) (табл. 3). Это коррелирует с данными археологии о формировании могильника Абрамово-10 позднесредневековыми сибирскими татарами [Молодин и др., 1990; Соболев, 2008]. Интересно, что могильник Абрамо-во-10 рассматривается археологами как сформированный барабинскими татарами, но исследованная серия с точки зрения структуры генофонда мтДНК оказалась ближе к современным тоболо-иртышским татарам, населяющим более западные и северо-западные территории. Это может свидетельствовать об изменении ареалов отдельных групп сибирских татар за последние века. Этот вывод нуждается в дальнейшем подтверждении с помощью анализа дополнительных серий мтДНК из локальных позднесредневековых популяций сибирских татар, а также на уровне анализа мужского генофонда (полиморфизма Y-хромосомы). В этой связи, в частности, представляют интерес материалы позднесредневекового могильника Крючное-6 на средней Таре, по данным материальной культуры и погребальной практике близкого к Абрамово-10 [Молодин и др., 2012]. Их палеогенетическое исследование уже начато авторами.
Низкое значение межпопуляционного генетического расстояния также зафиксировано между Абрамово-10 и современными выборками селькупов и хантов, что подтверждает участие самодийских и угорских генетических компонентов в формировании рассматриваемого нами позднесредневекового населения. Это подтверждается и результатами филогеографи-ческого анализа вариантов Абрамово-10, и анализом общих гаплотипов. Часть компонентов из Абрамово-10, помимо современных популяций сибирских татар, также характерна для западносибирских угров (хантов и манси) [Губина и др., 2005; Derbeneva et al., 2002] и самодийцев (селькупов) [Tamm et al., 2007]. Отметим, что источником как минимум части этих угро-самодийских компонентов могли быть автохтонные группы населения лесостепной зоны Западной Сибири предшествующих эпох. Прежде всего это касается таких кластеров, как
A10, Z, U2e, U5a, длительно присутствующих в генофонде популяций региона [Молодин и др., 2013; Pilipenko et al., 2015].
Таблица 3
Оценка генетического расстояния по Слаткину между исследованной в данной работе серией из Абрамово-10 и рядом современных популяций региона
Estimation of the Slatkin’s genetic distance between the Abramovo-10 series and some of the modern populations of the region
Table 3
|
Популяция |
Fst по Слаткину |
Источник данных о популяции |
|
Тоболо-иртышские татары |
0.00998 |
Naumova et al., 2008 |
|
Селькупы |
0, 01193 |
Tamm et al., 2007 |
|
Барабинские татары |
0.01958 |
Gubina et al., 2018 |
|
Ханты |
0, 02174 |
Губина и др., 2005 |
|
Хакасы |
0.03314 |
Derenko et al., 2007 |
Следует отдельно отметить, что сходство серий Абрамово-10 и селькупов (см. табл. 3) отчасти может объясняться спецификой современной селькупской серии: в ней присутствует необычно много носителей двух гаплотипов, которые также представлены и в Абрамово-10 (№ 11 и 21). Вероятно, это может быть связано с особенностями формирования выборки современных селькупов и попаданием в нее больших групп близких родственников или с особенностями локальной группы северо-селькупского населения [Tamm et al., 2007]. Эти предположения подтверждаются полученными нами предварительными результатами исследования большой серии образцов древней мтДНК от нарымских селькупов (данные готовятся к печати), в которой оба эти варианта слабо представлены или отсутствуют. Мы полагаем, что при использовании корректной сравнительной серии древних селькупских образцов генетическое сходство с Абрамово-10 с высокой степенью вероятности окажется не таким выраженным.
Филогеографический анализ вариантов мтДНК из могильника Абрамово-10 позволил выявить редкие варианты мтДНК, включая и специфичные для региона субкластеры. Например, вариант гаплогруппы T1a2a, выявленный у пяти индивидов из Абрамово-10 (№ 19), является специфичным для современных популяций лесостепной и таежной зон Западной Сибири: встречается только в генофонде сибирских татар, угорских и самодийских популяций региона. Очевидно, он может иметь автохтонное региональное происхождение. Некоторые варианты мтДНК из Абрамово-10 не были выявлены нами в подборке современных популяций Евразии, использованных для анализа (№ 2, 12, 17), а вариант № 9 был выявлен только у кетов [Derbeneva et al., 2002].
Филогеографический анализ также дает аргументы в пользу существования и других векторов внешних генетических связей для позднесредневекового тюркского населения западносибирской лесостепи. Ряд вариантов и субкластеров маркирует возможное генетическое влияние популяций Алтае-Саянского региона. Они ожидаемо относятся к восточно-евразийскому кластеру гаплогрупп мтДНК, а именно к гаплогруппам C и D. Субкластер C5b1, представленный в серии из Абрамово-10 одним вариантом (но девятью носителями, № 5), является общим для современного коренного населения Западной Сибири (тюркского, угорского и самодийского) и современных тюрков Алтае-Саянского региона, а также монголоязычных народов Восточной Сибири и Центральной Азии. Разнообразие вариантов свидетельствует, что именно Южная Сибирь является местом формирования этого кластера. Другими вариантами, связанными с южно-сибирскими популяциями Алтая, а также более вос- точных регионов, являются C4a1a4a (№ 3) и линия 16218T-16223T-16362C гаплогруппы D (№ 6). В отличие от C5b1 они слабо представлены в Западной Сибири: либо полностью отсутствуют в генофонде современных западносибирских популяций (№ 6), либо встречаются здесь только у тюркоязычных групп, включая барабинских татар (C4a1a4, № 3). По-ви-димому, эти варианты играли меньшее значение для дальнейших этапов формирования генофонда коренного населения Западной Сибири. Однако их присутствие в серии из Абра-мово-10 свидетельствует в пользу генетических контактов с популяциями Алтае-Саянского региона в период функционирования рассматриваемого нами могильника, чему имеются археологические, этнографические [Селезнев, 1994] и антропологические [Ким, 1998] свидетельства.
Таким образом, наши результаты подтверждают, что в основе механизма формирования генетического состава сибирских татар лежит взаимодействие групп тюркского, угорского и самодийского происхождения. В результате в регионе формируются популяции, имеющие смешанный генетический состав при варьирующем на разных памятниках вкладе компонентов различного происхождения. Полученные нами первые данные о генофонде позднесредневекового населения Центральной Барабы хорошо коррелируют с данными археологии и антропологии и сложной картиной взаимодействия на территории региона контрастных в этнокультурном отношении групп населения. Существует дискуссия относительно времени завершения этногенеза барабинских татар (и других групп сибирских татар): происходит ли консолидация этносов в XIII–XIV вв. [Молодин и др., 1990; Соболев, 2008] или позже [Селезнев, 1994]. К периоду функционирования могильника Абрамово-10 (XVI–XVIII вв.) эти процессы, очевидно, уже были завершены. В этот период одним из важных факторов динамики генетического состава населения становится влияние на коренные популяции пришлого европеоидного населения. Сравнение серии из Абрамово-10 и современных барабинских и тоболо-иртышских татар не выявляет существенных изменений в структуре генофонда мтДНК, которые можно было бы считать признаками влияния пришлого европеоидного населения. Так как из выборок современных сибирских татар были исключены индивиды, в родословной которых наблюдалась недавняя метисация, этот вывод нельзя распространить на все современное коренное население. Однако как минимум часть коренного тюркоязычного населения продолжает сохранять прежнюю структуру генофонда мтДНК, в том числе за счет высокого уровня эндогамных браков внутри локальных групп сибирских татар.
Заключение
Основной особенностью средневековых этногенетических процессов на территории лесостепной и южно-таежной зон Западной Сибири, которые рассматриваются в рамках данной работы, является взаимодействие нескольких контрастных в культурном, лингвистическом и генетическом отношении групп населения. На протяжении нескольких этапов Средневековья продолжался процесс тюркизации населения лесостепной зоны. Представленные в статье первые данные по структуре генофонда позднесредневекового населения, полученные нами на материалах могильника Абрамово-10, подтверждают участие компонентов тюркского, угорского и самодийского происхождения в формировании генофонда коренного населения Центральной Барабы. Выявленное сходство позднесредневековой серии мтДНК с выборками от современных групп сибирских татар подтверждает, что к началу периода функционирования могильника, т. е. к началу второй половины II тыс. н. э., уже сложилась структура генофонда тюркоязычного населения лесостепной зоны Западной Сибири. При этом ареалы локальных групп сибирских татар, по-видимому, могли меняться в позднем Средневековье и Новом времени, о чем свидетельствует большее сходство серии мтДНК из Абрамово-10 с современными тоболо-иртышскими татарами по сравнению с барабинскими татарами, проживающими непосредственно в Центральной Барабе. Помимо влияния со стороны населения степного пояса Евразии и лесостепных / южно-таежных популяций, определенную роль в формировании состава населения, оставившего могильник Абрамово-10, могли играть популяции Южной Сибири, прежде всего Алтае-Саянского региона. Несмотря на нарастающую роль культурного и генетического влияния мигрирующего в регион с запада европеоидного (преимущественно русского) населения, как минимум часть коренного тюркоязычного населения продолжает сохранять основные черты структуры генофонда мтДНК.
В данной работе вводится в научный оборот первая крупная серия ( N = 72) образцов мтДНК от позднесредневекового населения лесостепной зоны Западной Сибири. Сложность этногенетических процессов в регионе требует исследования многочисленных репрезентативных серий от населения региона, как позднесредневекового (и Нового времени), так и относящегося к предшествующим периодам эпохи Средневековья. Важнейшим направлением исследования является анализ мужского генофонда, наряду с генофондом мтДНК, что позволит получить более целостную и объективную картину популяционно-генетических процессов. Наличие большого объема палеоантропологического материала, доступного для исследования, и возможности анализа репрезентативного серийного материала современными палеогенетическими методами позволяют авторам данной работы выполнить масштабное систематическое исследование структуры генофонда средневекового населения региона во временной динамике с помощью сочетания синхронного и диахронного популяционного анализа и осуществить объективную реконструкцию этногенетических процессов в Западной Сибири, завершившихся формированием современного населения. Эти результаты будут изложены в серии работ, которую и открывает данная статья.
