Разнообразие вариантов митохондриальной ДНК у носителей окуневской культуры из могильника Сыда V (Минусинская котловина)
")
Автор: Пилипенко И. В., Пристяжнюк М. С., Трапезов Р. О., Черданцев С. В., Молодин В. И., Пилипенко А. С.
Журнал: Вестник Новосибирского государственного университета. Серия: История, филология @historyphilology
Рубрика: Археология Евразии
Статья в выпуске: 7 т.21, 2022 года.
Бесплатный доступ
Статья посвящена результатам анализа серии образцов митохондриальной ДНК (N = 25) из могильников окуневской культуры Минусинской котловины. Проведенное исследование позволило почти вдвое увеличить суммарную численность успешно исследованных окуневских образцов мтДНК. Исследованная нами серия образцов из могильника Сыда V (N = 23) является первой репрезентативной локально-территориальной серией образцов мтДНК для популяции окуневской культуры региона. Полученные нами новые результаты и опубликованные данные обсуждаются в контексте проблемы формирования генетического состава носителей культуры и их роли в дальнейшей генетической истории населения южных районов Сибири. Согласно имеющимся данным, ключевую роль в формировании окуневской популяции играли генетические компоненты, связанные с автохтонным населением региона, предшествовавшим появлению на юге Сибири афанасьевских популяций. Для более детальных реконструкций требуется накопление дополнительных репрезентативных палеогенетических данных.
Окуневская культура, минусинская котловина, эпоха развитой бронзы, палеогенетика, митохондриальная днк, y-хромосома, этногенетические реконструкции
Короткий адрес: https://sciup.org/147238469
IDR: 147238469 | УДК: 572, | DOI: 10.25205/1818-7919-2022-21-7-53-71
Diversity of mitochondrial DNA variants within Okunevo culture carriers from the Syda V burial ground (Minusinsk basin)
Purpose. Despite a long study, the problem of the Okunevo population origin remains unresolved, including the level of participation of autochthonous and possible migrant groups in the formation of its genetic composition. The purpose of this article is to obtain new data on the mitochondrial DNA diversity in the Okunevo population, in particular in a series of samples from the Syda V burial ground, and also to interpret these new paleogenetic results in the light of possible mechanisms for the formation of specific features of the Okunevo gene pool.Results. We successfully analyzed structure, phylogeny and phylogeography of 25 new mitochondrial DNA samples from the burial grounds of the Okunevo culture of the Minusinsk basin. The study made it possible to almost double the total number of successfully studied Okunevo mtDNA samples. The series of samples we studied from the Syda V burial ground (N = 23) is the first high representative local-territorial series of mtDNA samples for the Okunev population of the region. The new results and literature data are discussed in the context of the problem of the formation of the Okunevo population`s genetic composition and its role in the further genetic history of the southern regions of Siberia.Conclusion. The main role in the formation of the Okunev population were played by genetic elements of autochthonous origin associated with the aboriginal pre-Afanasievo groups. Possible role of migrants remains unclear and requires further research using paleogenetic methods.
Текст научной статьи Разнообразие вариантов митохондриальной ДНК у носителей окуневской культуры из могильника Сыда V (Минусинская котловина)
Минусинская котловина представляет собой один из наиболее хорошо исследованных с археологической точки зрения районов юга Сибири. За многие десятилетия проведения интенсивных археологических работ для данного региона накоплены и систематизированы богатейшие археологические и палеоантропологические коллекции, отражающие основные этапы формирования и развития культуры и популяционной структуры населения. Наиболее обширные материалы накоплены для различных периодов эпохи бронзы и раннего железного века, что позволило не только разработать обоснованную классификацию и периодизацию основных этнокультурных образований, существовавших в Минусинской котловине на протяжении III тыс. до н. э. – I тыс. н. э., но и приступить к разработке более подробной периодизации этапов развития каждой из выявленных основных археологических культур. Среди носителей археологических культур Минусинской котловины эпохи бронзы (афанасьевская, окуневская, андроновская (федоровская), карасукская, лугавская культуры) одной из ключевых популяций для объективной реконструкции истории населения региона является окуневская культура, датируемая второй половиной III тыс. до н. э. – II тыс. до н. э. (доанд-роновский период развитой бронзы), которая была выделена в качестве отдельной археологической культуры около 50 лет назад Г. А. Максименковым [1965; 1975], хотя первые раскопки памятников, впоследствии отнесенных к данной культуре, состоялись намного раньше [Теплоухов, 1929]. Несмотря на обширные объемы накопленных источников по материальной культуре окуневского населения и репрезентативные палеоантропологические коллекции, для этого важнейшего этнокультурного образования Южной Сибири всё еще не находят однозначного ответа вопросы об источниках и механизмах ее формирования, а также о роли окуневцев в более поздней этнокультурной истории региона. Эти дискуссии, первоначально проводившиеся с привлечением данных археологии и физической антропологии [Максименков, 1975; Вадецкая и др., 1980; Лазаретов, 1997], в последнее время вновь интенсифицировались с привлечением в качестве дополнительных аргументов данных палеогенетики [Поляков, 2019; 2022; Козинцев, 2020]. При этом с точки зрения популяционно-генетической истории основной темой для дискуссии является степень участия автохтонных южносибирских и, напротив, мигрантных западно-евразийских компонентов (а также происхождение последних) в процессах формирования генетического состава окуневского населения. Важно отметить, что, в отличие от археологического и антропологического, палеогенетическое исследование носителей окуневской культуры находится на одной из начальных стадий и далеко от формирования исчерпывающего генетического портрета этой группы древнего населения Южной Сибири. Вплоть до настоящего времени в палеогенетической литературе отсутствуют работы, сфокусированные на генетических характеристиках окуневской популяции. Окуневцы присутствуют лишь в составе географически и хронологически широких выборок, исследование которых направлено на реконструкцию масштабных популяционногенетических процессов в Евразии [Allentoft et al., 2015; Damgaard et al., 2018], или в составе небольших выборок разновременного населения Южной Сибири [Hollard et al., 2018]. Коллегами археологами и антропологами предпринимались попытки (весьма информативные) обзора имеющихся палеогенетических данных и их использования для обсуждения проблем происхождения и дальнейшей истории окуневской популяции [Поляков, 2019; 2022; Козинцев, 2020]. Результаты этих работ, и в частности имеющиеся противоречия между ними, очевидно свидетельствуют, что для полноценной реконструкции генетической истории окуневского населения необходимо накопление значительного дополнительного объема генетических данных, включая как анализ отдельных филогенетически информативных маркеров, прежде всего митохондриальной ДНК (мтДНК) и Y-хромосомы, так и методы полногеномного анализа с использованием высокопроизводительных методов палеогеномики. Примечательно, что к настоящему моменту ни по одному из этих направлений не достигнут высокий уровень репрезентативности исследованных выборок, а следовательно, все эти направления остаются актуальными. Одним из направлений развития палеогенетики в последние годы является переход от анализа географически и хронологически широких моделей (в которых каждая из групп древнего населения представлена небольшой выборкой или вообще единичными образцами) к анализу репрезентативных серий для отдельных групп древнего населения. Не является исключением и окуневская популяция. Такие серии помимо суммарной численной репрезентативности должны включать образцы из разных археологических памятников, зачастую отражающих локально-территориальные особенности генетической структуры населения и / или особенности отдельных этапов становления или развития ис- следуемого древнего этнокультурного образования. В связи с этим для детализации имеющихся представлений о генофонде окуневского населения необходимо получение палеогене-тических результатов из могильников, для которых эти данные не были получены ранее на уровне репрезентативных серий. В данной работе мы представляем результаты исследования разнообразия мтДНК в серии (N = 25), представленной преимущественно образцами из окуневского могильника Сыда V, а также единичными образцами мтДНК из двух других окуневских могильников (подробности см. в разделе «Материалы и методы»). Полученные результаты рассмотрены на фоне ранее накопленных данных о генофонде окуневского населения (и других популяций региона), а также обсуждены в контексте проблем происхождения окуневской популяции и ее роли в генетической истории населения региона (хотя на данном этапе окончательное решение этих проблем с применением методов палеогенетики требует проведения значительных дальнейших исследований).
Материалы и методы
Палеоантропологические материалы, исследованные в работе. Все образцы, исследованные в данной работе, получены из состава палеоантропологической коллекции ИАЭТ СО РАН (Новосибирск). Отбор образцов выполнен палеогенетиком из ИЦиГ СО РАН (А. С. Пилипенко) совместно с археологами и антропологами из отдела археологии палеометалла ИАЭТ СО РАН (рук. В. И. Молодин). В состав выборки были включены материалы трех могильников окуневской культуры. Основу исследованной серии образцов составили палеоантропологические материалы из могильника окуневской культуры Сыда V, расположенного на первой надпойменной террасе правого берега р. Сыда в 300 м от впадающей в нее р. Уза, в самом центре с. Сыда в Сыда-Ербинской котловине, которая является частью Минусинской котловины (рис. 1). Раскопки могильника проводились с 1965 г. Красноярской археологической экспедицией ЛОИА АН СССР, под руководством М. П. Грязнова [Грязнов, Комарова, 2006]. Палеоантропологические материалы, обнаруженные при раскопках памятника Сы-да V, были переданы авторами раскопок в состав антропологической коллекции ИАЭТ СО РАН, где и находятся в настоящее время.
Для выполнения палеогенетического исследования были отобраны образцы от 31 индивида из погребений могильника Сыда V. Кроме того, один образец был получен от носителя окуневской культуры из могильника Барсучиха II, и два образца – от индивидов из могильника Уйбат. Таким образом, общая численность исследуемой в нашей работе выборки носителей окуневской культуры составила 34 индивида (табл. 1). Образцы представлены костями посткраниального скелета и зубами. Для проведения исследования от каждого скелета отбирали части с признаками наибольшей макроскопической сохранности.
Предварительная обработка материала и получение образцов суммарной ДНК. Использовались методы, детально описанные в наших предшествующих работах [Pilipenko et al., 2015; 2018]. Поверхность посткраниальных костей очищали от видимых загрязнений, обрабатывали раствором гипохлорита натрия, облучали ультрафиолетом для разрушения современной ДНК. Затем механически удаляли поверхностный слой на глубину ~ 1–2 мм и повторно облучали образец ультрафиолетом. Из компактного костного вещества высверливали порошок. Поверхность зубов механически очищали от загрязнений. Далее зубы целиком обрабатывали гипохлоритом натрия, облучали ультрафиолетом и размалывали с помощью шаровой мельницы Retsch MM200 (Германия).
Для выделения ДНК из посткраниального материала костный порошок инкубировали в 5М гуанидинизотиоционатном буфере при температуре 65 ºС и постоянном перемешивании. Материал зубов декальцинировали 0,5-молярным раствором ЭДТА с последующим лизисом с помощью протеиназы K. ДНК выделяли методом фенол-хлороформной экстракции с последующим осаждением изопропанолом.
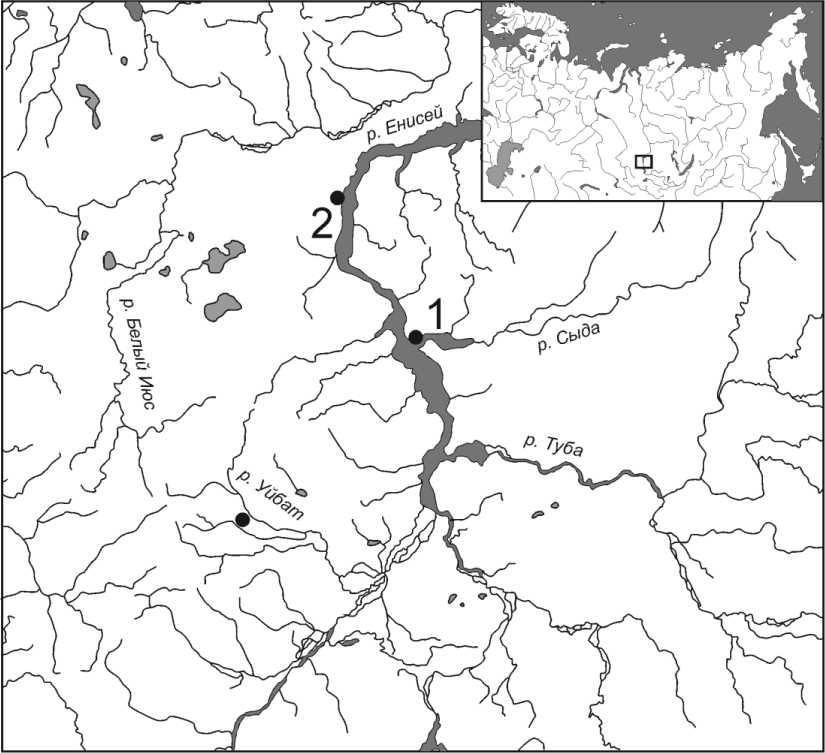
Рис. 1. Расположение могильников окуневской культуры в Минусинской котловине, из которых был получен материал для проведения палеогенетического исследования: 1 – Сыда V; 2 – Барсучиха II; 3 – комплекс памятников Уйбат
Fig. 1. The location of the burial grounds of the Okunevo culture in the Minusinsk basin, from which the material for paleogenetic research was obtained: 1 – Syda V; 2 – Badger II; 3 – a complex of Uybat monuments
Анализ структуры мтДНК включал определение последовательности первого гипервариабельного сегмента контрольного района мтДНК (ГВСI мтДНК). Амплификацию ГВС I мтДНК проводили параллельно двумя методами: 1) в виде четырех коротких перекрывающихся фрагментов (четыре независимых однораундовых ПЦР [Haak et al., 2005]); 2) в виде одного длинного фрагмента с помощью «вложенной» ПЦР (включала два последовательных раунда реакции) [Пилипенко и др., 2008]. Статус филогенетически информативных позиций в кодирующей части мтДНК определяли с помощью ПЦР-систем, описанных в работе [Wilde et al., 2014].
Последовательности нуклеотидов определяли с использованием набора реактивов ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (v.1.1 – для коротких фрагментов, v.3.1 – для длинного фрагмента) (Applied Biosystems, СШA). Продукты секвенирующей реакции анализировали на автоматическом капиллярном секвенаторе ABI Prism 3130XL Genetic Analyser (Applied Biosistems, США) в центре коллективного пользования «Геномика» СО РАН ( http://sequest.niboch.nsc.ru ). Филогенетическую и филогеографическую интерпретацию результатов осуществляли методами, описанными нами ранее [Pilipenko et al., 2018].
Таблица 1
Описание серии палеоантропологических образцов от носителей окуневской культуры, использованных для проведения данного исследования
Table 1
The description of the series paleoanthropological samples from the Okunevo culture population which were used to maintain this research
|
Лабораторный шифр индивида |
Археологический памятник |
Описание * |
Материал для получения образца ДНК |
|
OK1 |
Сыда V |
К. 4, М. 6, Ск.1 |
Бедренная кость |
|
OK2 |
Сыда V |
К. 4, М. 6, Ск. 2 |
Бедренная кость |
|
OK3 |
Сыда V |
К. 4, М. 6, Ск. 3 |
Бедренная кость |
|
OK4 |
Сыда V |
К. 4, М. 6, Ск. 4 |
Бедренная кость |
|
OK5 |
Сыда V |
К. 4, М. 6, Ск. 5 |
Бедренная кость |
|
OK6 |
Сыда V |
К. 4, М. 6, Ск. 6 |
Бедренная кость |
|
OK7 |
Сыда V |
К. 4, М. 6, Ск. 7 |
Бедренная кость |
|
OK8 |
Сыда V |
К. 4, М. 6, Ск. 8 |
Бедренная кость |
|
OK9 |
Сыда V |
К. 3, М. 1, Ск. 1 |
Бедренная кость |
|
OK10 |
Сыда V |
К. 2, М. 2, Ск. 1 |
Бедренная кость |
|
OK11 |
Сыда V |
К. 4, М. 1 |
Большая берцовая кость |
|
OK12 |
Сыда V |
К. 3, М. 1, Ск.1 |
Бедренная кость |
|
OK13 |
Сыда V |
К. 3, М. 12, Ск. 2 |
Бедренная кость |
|
OK14 |
Сыда V |
К. 1, М. 2 |
Большая берцовая кость |
|
OK15 |
Сыда V |
К. 4, М. 10 |
Бедренная кость |
|
OK16 |
Сыда V |
К. 3, М. 1, Ск. 2 |
Бедренная кость |
|
OK17 |
Сыда V |
К. 1, М. 4 |
Большая берцовая кость |
|
OK18 |
Сыда V |
К. 3, М. 22 |
Бедренная кость |
|
OK19 |
Сыда V |
К. 1, М. 3 |
Плечевая кость |
|
OK20 |
Сыда V |
К. 4, М. 5 |
Большая берцовая кость |
|
OK21 |
Сыда V |
К. 3, М. 1, Ск. 2 |
Большая берцовая кость |
|
OK22 |
Сыда V |
К. 3, М. 1, Ск. 11 |
Большая берцовая кость |
|
OK23 |
Барсучиха-2 |
К. 22, М. 2 |
Бедренная кость |
|
OK24 |
Сыда V |
К. 3, М. 9 |
Плечевая кость |
|
OK25 |
Сыда V |
К. 3, М. 2, Ск. 2 |
Зубы |
|
OK26 |
Сыда V |
К. 3, М. 10, Ск. 1 |
Зубы |
|
OK27 |
Сыда V |
К. 3, М. 10, Ск. 2 |
Зубы |
|
OK28 |
Сыда V |
К. 3, М. 12, Ск. 1 |
Зубы |
|
OK29 |
Сыда V |
К. 3, М. 10, Ск. 3 |
Зубы |
|
OK30 |
Сыда V |
К. 3, М. 5, Ск. 1 |
Зубы |
|
OK31 |
Сыда V |
К. 3, М. 5, Ск. 2 |
Зубы |
|
OK32 |
Уйбат |
К. 1, М. 2А |
Зубы |
|
OK33 |
Уйбат |
К. 1, М. 2Б |
Зубы |
|
OK34 |
Сыда V |
К. 2, М. 2, Ск. 2 |
Зубы |
*
Сокращения: К. – курган, М. – могила, Ск. – скелет.
Меры против контаминации и верификация результатов. Все работы с древним материалом выполнены в специально оборудованных для палеогенетических исследований чистых помещениях межинститутской лаборатории молекулярной палеогенетики и палеогеномики ИЦИГ СО РАН (Новосибирск, Россия). Использованная инфраструктура и приборный парк отвечают всем современным требованиям, предъявляемым к палеогенетическим лабораториям. Меры против контаминации и процедуры верификации результатов описаны в нашей статье [Pilipenko et al., 2018] и включали: использование средств индивидуальной защиты и логистики исследования, исключающих загрязнение образцов в процессе работы, проведение всех стадий работы в изолированных друг от друга помещениях лаборатории, многократное воспроизведение экспериментальных процедур для каждого образца, контроль загрязнения реактивов и спорадического загрязнения проб на всех этапах работы и др. Дополнительно для ряда образцов (12 индивидов из исследованной серии) были установлены воспроизводимые профили (полные или частичные) 15 аутосомных STR-локусов и определен генетический пол останков (по полиморфизму гена амелогенина) с использованием коммерческого набора реактивов AmpFlSTR® Identifiler® Plus PCR Amplification Kit (Applied Biosystems, США), который позволяет эффективно контролировать наличие / отсутствие контаминации современной ДНК, а также устанавливать индивидуальный генетический профиль, отличающий каждого из исследуемых индивидов (традиционно используется для анализа степени родства индивидов). Условия проведения исследования, используемые протоколы работы и особенности полученных нами результатов однозначно свидетельствуют о высокой достоверности полученных нами палеогенетических данных.
Результаты исследования
Были получены 3–5 независимых экстрактов суммарной ДНК из останков всех 34 носителей окуневской культуры, включенных в выборку для исследования. Успешно определена последовательность ГВС I мтДНК для 25 из 34 индивидов, что составляет ~ 73,5 % первоначальной выборки (табл. 2). Таким образом продемонстрирована высокая степень сохранности ДНК в останках большей части исследуемых представителей окуневской культуры, что коррелирует с хорошей макроскопической сохранностью окуневского палеоантропологического материала. Для оставшихся 9 индивидов, первоначально включенных в серию, получить полноценные и воспроизводимые данные о структуре мтДНК не удалось. Отчасти это связано с низкой сохранностью ДНК в образцах, не позволившей амплифицировать все участки ГВС I мтДНК (для 7 индивидов). Для 2 индивидов выявлены признаки тотальной контаминации материала современной ДНК, поэтому они также были исключены из анализа.
В исследованной серии из 25 образцов выявлено 10 структурных вариантов (гаплотипов) ГВС I мтДНК. Шесть структурных вариантов выявлены лишь у одного индивида из серии, 4 структурных варианта выявлены у нескольких индивидов (от 2 до 8), при этом варианты 1 и 3 (табл. 2) представлены с высокой частотой (8 и 6 индивидов соответственно). На основе данных о последовательности ГВС I мтДНК и статусе информативных гаплогрупп-специ-фичных позиций в кодирующей части мтДНК было определено филогенетическое положение и установлена принадлежность к гаплогруппам всех 25 успешно исследованных образцов мтДНК, и построено дерево, отражающее филогенетические отношения исследованных структурных вариантов мтДНК (рис. 2).
Учитывая тот факт, что в трех ранее опубликованных работах зарубежных коллег, в которых проводился анализ образцов из окуневской популяции, присутствуют данные о структуре и филогенетической принадлежности 32 окуневских образцов мтДНК [Allentoft et al., 2015; Damgaard et al., 2018; Hollard et al., 2018], исследованная нами серия увеличивает суммарную выборку мтДНК для этой популяции почти на 80 %, а ее суммарная численность теперь превышает 50 образцов (индивидов). При этом выборка из могильника Сыда V, исследованная нами (N = 22), является наиболее репрезентативной серией из одного могильника среди всех опубликованных к настоящему моменту (для других могильников исследованы серии не более 6–7 образцов).
Таблица 2
Разнообразие структурных вариантов мтДНК в исследованной серии носителей окуневской культуры Минусинской котловины
The variety of structural variants of mtDNA in the researched series of Okunevo culture population of the Minusinsk basin
Table 2
|
№ варианта |
Гаплотип ГВС I мтДНК |
Статус ОНП в кодирующей части мтДНК |
Гаплогруппа мтДНК |
Число носителей варианта в исследованной серии |
Носители данного варианта |
|
1 |
16223-16242 16278-16290 16319 |
663G |
A8 |
6 |
OK1, OK7, OK17, OK18, OK19, OK32 |
|
2 |
16242-16290- 16293C-16319 |
663G |
A11 |
1 |
OK23 |
|
3 |
16093-16223 16288-16298 16327 |
13263G |
C5 |
8 |
OK2, OK3, OK8, OK9, OK11, OK13, OK22, OK29 |
|
4 |
16239-16298 16327 |
13263G |
C |
1 |
OK25 |
|
5 |
CRS |
7028C |
H/U |
1 |
OK16 |
|
6 |
16362 |
7028C |
H |
1 |
OK14 |
|
7 |
16051-16129C- 16183C-16189 |
12308G |
U2e |
1 |
OK24 |
|
8 |
16356 |
12308G |
U4 |
2 |
OK6, OK33 |
|
9 |
16192-16256 16270 |
12308G |
U5a1 |
3 |
OK10, OK12, OK21 |
|
10 |
16129-16192 16256-16270 16318 |
12308G |
U5a1 |
1 |
OK15 |
В исследованной нами серии обнаружены варианты гаплогрупп мтДНК, относящихся к западно-евразийскому (Н, U2e, U4, U5a1) и восточно-евразийскому (А8, А11, С*, С5) кластерам (см. рис. 2). Варианты западно-евразийского кластера мтДНК доминируют в отношении числа гаплотипов (6 из 10 выявленных гаплотипов), но уступают восточно-евразийским по численности их носителей (всего 9 носителей западно-евразийских вариантов из 25 исследованных индивидов). Такое несоответствие между разнообразием вариантов и общей долей их носителей объясняется высокой частотой встречаемости двух из четырех восточноевразийских вариантов – № 1 и 3 в табл. 2, относящихся в гаплогруппам A8 и С5 соответственно, в то время как другие два восточно-евразийские варианта (№ 2 и 4 в табл. 2) встречены у одного индивида каждый.
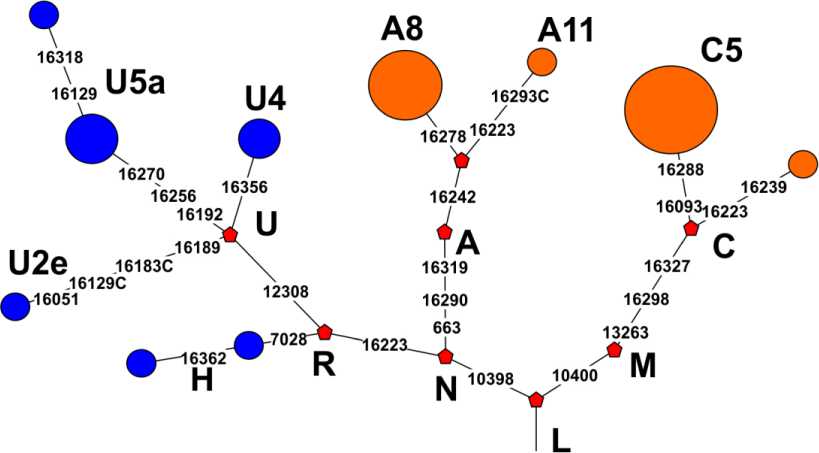
Рис. 2. Филогенетическое дерево структурных вариантов мтДНК, выявленных в генофонде окуневской популяции Минусинской котловины. Синим цветом обозначены варианты, относящиеся к западно-евразийскому кластеру гаплогрупп мтДНК, оранжевым – к восточно-евразийскому кластеру гаплогрупп мтДНК
Fig. 2. Phylogenetic tree of mtDNA structural variants identified in the gene pool of the Okunevo population of the Minusinsk basin. Blue indicates variants related to the Western Eurasian cluster of mtDNA haplogroups, orange – to the Eastern Eurasian cluster of mtDNA haplogroups
Следует отметить, что наличие выраженных компонентов, сформированных вариантами западно- и восточно-евразийского происхождения, было характерно и для ранее исследованных серий мтДНК окуневцев [Allentoft et al., 2015; Damgaard et al., 2018; Hollard et al., 2018], среди которых восточно-евразийские варианты также преобладали, составляя две трети выборки, что сопоставимо с нашей серией. Другим важным сходством нашей серии с ранее опубликованными является высокая частота вариантов гаплогрупп A8 и С5, зафиксированная не только на могильнике Сыда V, но и ранее на других памятниках. Таким образом, высокую частоту вариантов восточно-евразийских гаплогрупп A8 и С5, отмеченную в нашей серии, следует рассматривать скорее не как специфичную черту группы, сформировавшей могильник Сыда V, а как распространенную черту генофонда окуневской популяции в целом, т. е. общую для локально-территориальных групп населения. Наряду с этим мы считаем несколько преждевременными выводы коллег, о том, что локально-территориальные группы окуневского населения, представленные материалами отдельных могильников, демонстрируют признаки «хаотичного смешения» в результате «частых перекрестных браков с представителями других обществ» [Поляков, 2019, с. 100]. Прежде всего, до получения нами данных о разнообразии мтДНК в окуневских комплексах могильника Сыда V в литературе попросту отсутствовали репрезентативные данные для какого-либо окуневского памятника: серии 2–7 образцов мтДНК нельзя считать выборками, репрезентативно отражающими состав генофонда мтДНК населения, сформировавшего тот или иной крупный могильник. Кроме того, исследованная нами серия из могильника Сыда V демонстрирует некоторые предварительные отличия от ранее исследованных серий. Например, для нее характерна бо́льшая доля вариантов различных подгрупп гаплогруппы U, U2e, U4, U5a; при этом гапло-группа U2e впервые выявлена нами в окуневском генофонде, в составе западно-евразийской части серии, и наоборот, меньшая доля вариантов гаплогруппы H, поэтому говорить о полном сходстве локальный серий на данном этапе нельзя. С другой стороны, эти отличия также могут объясняться недостаточной репрезентативностью большинства локальных серий образцов мтДНК (помимо серии из могильника Сыда V), поэтому мы предлагаем вернуться к обсуждению данного аспекта после накопления соответствующих репрезентативных генетических результатов.
Хотя основной целью нашей статьи является введение в научный оборот новых данных по структуре генофонда мтДНК окуневского населения Минусинской котловины, мы не можем остаться в стороне от дискуссии о вопросах происхождения окуневцев, которая в последнее время развернулась с новой силой, с активным привлечением имеющихся данных палеогенетики [Поляков, 2019; 2022; Козинцев, 2020]. В связи с этим дальнейшее обсуждение результатов анализа исследованной нами серии методами филогенетики и филогеографии мы будем проводить в контексте проблемы происхождения и механизмов формирования окуневской популяции. Важно подчеркнуть, что данные палеогенетики, как и данные физической антропологии, имеют непосредственное отношение лишь к популяционно-генетическим аспектам этногенетических процессов, т. е. отражают именно биологическую составляющую процессов формирования населения исследуемой территории, в то время как данные археологии отражают в первую очередь культурогенез. Эти аспекты этнокультурных процессов часто, но далеко не всегда коррелируют друг с другом. Ведь, например, изменение духовной и материальной культуры может сопровождаться миграционными процессами и межпопуляционным взаимодействием, а может быть обусловлено только культурными контактами (через торговлю и обучение), без миграционной составляющей и соответствующих заметных генетических контактов между популяциями. Более того, оценка степени корреляции культурогенетических и популяционно-биологических процессов сама по себе часто является интересной и сложной задачей междисциплинарного исследования. В этой связи нельзя однозначно согласиться с утверждением о второстепенной роли антропологии и палеогенетики в этнокультурных реконструкциях, по сравнению с археологическими подходами [Поляков, 2022, с. 132]. Каждое из этих научных направлений позволяет анализировать прежде всего отдельные аспекты этнокультурных процессов. И только их полноценное сочетание позволяет получить комплексную картину изучаемых событий прошлого. В данной работе мы анализируем генетические данные, поскольку они в первую очередь связаны с популяционно-генетическими аспектами формирования окуневского населения.
Основным дискуссионным моментом в вопросах происхождения окуневского населения является степень участия в его формировании автохтонных южносибирских компонентов, с одной стороны, и пришлых компонентов из более западных регионов Евразии, потенциально связанных с миграционными потоками с запада, с другой. При этом под автохтонными компонентами в данном случае подразумеваются генетические компоненты, свойственные доафанасьевскому населению региона (в частности неолитической и более ранним популяциям). А пришлые компоненты связывают либо с носителями афанасьевской культуры, имеющими пришлое происхождение [Хохлов и др., 2016], либо с отдельным эпизодом миграции из Восточной Европы, не относящимся напрямую к ямно-афанасьевскому феномену [Поляков, 2019; 2022].
Обратимся к имеющимся в нашем распоряжении данным по генофонду мтДНК окуневского населения, а также опубликованным ранее данным о генофонде Y-хромосомы этой популяции (данные по разнообразию Y-хромосомы окуневцев из могильника Сыда V будут опубликованы в другой статье, посвященной диахронному анализу мужского генофонда населения Минусинской котловины).
Мы уже отметили, что характерной чертой окуневского генофонда является «смешанный» состав из вариантов гаплогрупп западно- и восточно-евразийского происхождения. Отметим, что наличие в составе генофонда одновременно западно- и восточно-евразийских гаплогрупп является довольно распространенной чертой многих древних и современных популяций Южной и Западной Сибири, а также Центральной Азии и само по себе не может рассматриваться как доказательство смешанного происхождения той или иной популяции, так как пер- воначальное проникновение и смешение этих компонентов могло происходить в регионе задолго до возникновения исследуемой древней популяции и являться уже «реликтовым» признаком, характерным для предшествующих популяций региона. Например, на соседней территории Барабинской лесостепи такая «смешанная» структура зафиксирована для всех групп, доступных для исследования, включая и наиболее ранние популяции (неолит и эпоха раннего металла) [Молодин и др., 2013; Molodin et al., 2012], и сформировалась, по-види-мому, в еще более ранние эпохи. В частности варианты различных подгрупп западно-евразийской гаплогруппы U присутствовали в Барабе уже в неолите, наряду с рядом восточноевразийских гаплогрупп. Наиболее вероятно, такая смешанная структура могла быть характерна и для других районов юга Сибири задолго до начала процесса формирования окуневского населения и до первой масштабной миграционной волны на юг Сибири, связанной с ямно-афанасьевским феноменом. Иначе говоря, сама по себе структура генофонда мтДНК, состоящего из западно- и восточно-евразийского компонентов, свидетельствует в пользу участия автохтонных групп в формировании генофонда окуневцев. Более того, филогеогра-фический анализ с привлечением имеющихся в литературе данных о древних и современных популяциях Евразии четко свидетельствует, что целый ряд гаплогрупп, входящих в состав окуневского генофонда мтДНК, имеет выраженное сибирское (южносибирское) и / или центральноазиатское происхождение. Это в частности касается гаплогрупп A8, A11 и С5, выявленных как в нашей, так и в ранее опубликованных окуневских сериях. Кроме того, к числу несомненно автохтонных для южных районов Сибири кластеров мтДНК относится гапло-группа А10, выявленная у одного из носителей окуневской культуры [Hollard et al., 2018]. Ранее мы показали, что варианты гаплогруппы A10 являлись важным автохтонным компонентом генофонда мтДНК населения сопредельных районов западносибирской лесостепи, как минимум с эпохи неолита и на протяжении всей эпохи бронзы [Pilipenko et al., 2015]. Таким образом, автохтонное южносибирское происхождение основных гаплогрупп восточноевразийского кластера, выявленных в окуневском генофонде мтДНК, не вызывает сомнений с точки зрения данных филогеографии.
В составе западно-евразийской части генофонда мтДНК присутствуют варианты, которые могут с равной вероятностью происходить из автохтонных неолитических популяций региона или попасть в регион в связи с ямно-афанасьевской волной миграции из западных районов Евразии, это варианты гаплогрупп U2e, U4, U5a. Для остальных западно-евразийских гап-логрупп, выявленных в окуневском генофонде (прежде всего варианты гаплогруппы H, а также более редкие J, T), которые не были выявлены в ранних популяциях юга Сибири (неолит и ранний металл), вероятным является мигрантное происхождение с запада. При этом важно отметить, что практически все эти варианты (или очень близкие к ним) также выявлены в составе генофонда мтДНК афанасьевского или ямного населения (см. обзор данных в [Поляков, 2019; 2022] или первоисточники [Allentoft et al., 2015; Damgaard et al., 2018; Hollard et al., 2018; Narasimhan et al., 2019] и др.). Никаких явных признаков появления другого пришлого населения с запада, не связанного с афанасьевской культурой, в генофонде мтДНК окуневской популяции к настоящему моменту не выявлено (хотя полностью опровергнуть наличие такой миграции в небольшом масштабе также пока нельзя). В любом случае именно автохтонные для юга Сибири компоненты, составили основу генофонда мтДНК окуневского населения. А компоненты, потенциально связанные с возможной миграцией с запада, составляют относительно незначительную часть (порядка 20 % или менее) и могут быть объяснены влиянием афанасьевского населения.
С этим согласуются и имеющиеся данные по генофонду Y-хромосомы (как литературные, так и наши собственные, пока не опубликованные). Основной компонент в генофонде окуневского населения – вариант Q1a (в меньшей степени Q1b) гаплогруппы Y-хромосомы. Линии этой гаплогруппы довольно широко распространены в популяциях восточной части Евразии [Petkovski et al., 2006], в частности в Центральной Азии и Сибири, также ее можно встретить среди коренных народов Америки [Rasmussen et al., 2014]. При этом именно юж- ные районы Сибири, предположительно, являются местом ее первоначального происхождения и первичной диверсификации [Zegura et al., 2004]. Интересно, что ранее наличие гапло-группы Q было выявлено также у представителей населения соседнего Горного Алтая, также датируемых эпохой развитой бронзы (II тыс. до н. э.) [Пилипенко и др., 2016]. В целом сибирское или центрально-азиатское происхождение данного компонента мужского генофонда окуневской популяции не вызывает сомнений, что позволяет отнести его к числу потенциально автохтонных кластеров. Выявленные в меньшем количестве в составе окуневского мужского генофонда варианты кластера NO (NO1) [Hollard et al., 2018] сложнее в филогео-графической интерпретации, однако и они являются характерными для восточной и северовосточной части Евразии, т. е. не могут быть связаны с миграцией с запада. Наконец последний, также редкий (минорный) компонент генофонда мужской части окуневской популяции относится к специфической подгруппе гаплогруппы R1b (R1b1a2a) [Hollard et al., 2018; Damgaard et al., 2018], которая абсолютно доминирует среди вариантов Y-хромосомы в генофонде носителей афанасьевской и ямной культур и, как правило, маркирует генетическое влияние групп, связанных с ямно-афанасьевским феноменом. Отметим, что влияние это на окуневцев если и было, то выражено относительно слабо на уровне генофонда как мтДНК, так и Y-хромосомы.
Таким образом, компоненты потенциально автохтонного южносибирского происхождения доминируют как в женском, так и в мужском генофонде окуневского населения. По-ви-димому, именно автохтонные компоненты играли главную роль при формировании генетического состава окуневцев. Основной проблемой для окончательного (прямого) обоснования методами антропологии и палеогенетики главенствующей роли автохтонных для Южной Сибири генетических компонентов в формировании окуневского населения, как вполне справедливо было отмечено [Поляков, 2022, с. 132–134], является недостаток (в случае антропологии) или полное отсутствие (в случае палеогенетики) репрезентативных данных о доафанасьевском населении Минусинской котловины. И обращение к верхнепалеолитическим материалам Восточной Сибири [Козинцев, 2020] является полезным, но не может полностью компенсировать пробел в данных по неолитическим популяциям региона. Однако современные возможности филогенетического и филогеографического анализа данных по мтДНК и Y-хромосоме позволяют с высокой достоверностью фиксировать автохтонные компоненты генофонда, поэтому длительное присутствие ряда кластеров мтДНК из окуневской серии в южных районах Сибири (в данном случае автохтонность) не вызывает сомнений вне зависимости от наличия / отсутствия палеогенетических результатов по неолитическому населению Минусинской котловины. В этой связи резкая смена генетического состава населения, фиксируемая диахронным анализом при переходе от афанасьевской к окуневской культуре, с генетической точки зрения может быть обусловлена не притоком совершенно нового населения в регион (как это было в случае самого афанасьевского населения и позже, в период андроновской миграции на юг Сибири), а резким увеличением роли автохтонных (доафанасьевских) генетических компонентов. С точки зрения популяционной биологии такой сценарий мог быть реализован в случае первоначального оттеснения автохтонных популяций с территории Минусинской котловины с приходом в регион афанасьевского населения с последующей репопуляцией этих территорий преимущественно автохтонными по происхождению группами после исчезновения или ослабления здесь (по тем или иным причинам) афанасьевцев с формированием окуневского населения. С этим хорошо согласуется и установленный в последние годы сравнительно короткий период бытования в Минусинской котловине памятников афанасьевской культуры [Poliakov et al., 2019]. Этот сценарий коррелирует с моделью, предложенной ранее Г. А. Максименковым [1975]. Однако необходимо подчеркнуть, что в данном случае мы говорим лишь о популяционно-генетических аспектах этнокультурных процессов, что не отменяет ряд сложностей, возникающих с пониманием культурогенетических аспектов рассматриваемых процессов, справедливо отмеченных А. В. Поляковым [2022, c. 132–134]. При этом важно отметить, что в полученных к настоя- щему моменту палеогенетических данных пока отсутствуют компоненты западного происхождения, которые можно было бы потенциально связать с отдельным от ямно-афанасьев-ского миграционным потоком с запада преимущественно мужского населения, который мог бы внести определяющий вклад в формирование окуневской популяции. Речь может идти о носителях сейминско-турбинского транскультурного феномена, проникавших на территорию Минусинской котловины с запада и ослабивших специфические следы, вероятно, в комплексах окуневской культуры [Молодин, 2022] В любом случае для дальнейшей реконструкции популяционно-генетических аспектов формирования окуневского населения необходимо накопление большого объема дополнительных данных по различным генетическим маркерам как для окуневских групп, так и для популяций Южной Сибири предшествующих и более поздних периодов.
Важно также отметить, что некоторые из автохтонных для юга Сибири генетических компонентов, формирующих основу окуневского генофонда, такие как гаплогруппы A8 и C5 мтДНК, сохраняют свое значения для популяций юга Сибири и в последующие эпохи [Pilipenko et al., 2018], в отличие от компонентов, связанных с афанасьевским населением, которые, по-видимому, полностью исчезают из регионального генофонда.
Заключение
Проведенное нами исследование серии из 25 образцов мтДНК из останков носителей окуневской культуры Минусинской котловины позволило существенно (почти вдвое) увеличить численность их суммарной серии, опубликованной к настоящему времени. Поскольку 22 из 25 исследованных образцов были взяты из одного могильника – Сыда V, мы получили первые репрезентативные данные о разнообразии вариантов мтДНК в одном могильнике, так как для других могильников к настоящему времени было исследовано не более 6–7 образцов мтДНК. Это отвечает современным тенденциям развития палеогенетики, когда наблюдается переход от анализа географически и хронологически разрозненных образцов к работе с репрезентативными локально-территориальными сериями, позволяющими получить более полное представление о деталях генетического состава отдельных групп древнего населения.
Полученные нами данные вместе с накопленными ранее в научной литературе были рассмотрены в контексте проблем происхождения и генетической истории окуневской популяции. Хотя мы еще не можем выполнить исчерпывающую этногенетическую реконструкцию, уже сейчас очевидно, что основу окуневского генофонда мтДНК и Y-хромосомы составили компоненты, автохтонные для южных районов Западной и Восточной Сибири, т. е. связанные с доафанасьевскими популяциями региона. В то же время участие компонентов миграционного происхождения из западной части Евразии существенно менее выражено, а признаков участия в формировании окуневского генофонда миграционного потока с запада, отличного от миграции ямно-афанасьевских групп, на данный момент в генофонде не выявлено. Дальнейший прогресс в реконструкции популяционно-генетических аспектов становления и развития окуневского генофонда будет связан с накоплением репрезентативных данных о локально-территориальных и хронологических группах носителей окуневской культуры, что является актуальной задачей как на уровне отдельных филогенетически и филогеографически информативных маркеров (мтДНК, Y-хромосома, отдельные аутосомные маркеры), так и на уровне полногеномного анализа высокопроизводительными методами.
Список литературы Разнообразие вариантов митохондриальной ДНК у носителей окуневской культуры из могильника Сыда V (Минусинская котловина)
- Вадецкая Э. Б., Леонтьев Н. В., Максименков Г. А. Памятники окуневской культуры. Л.: Наука, 1980. 148 с.
- Грязнов М. П., Комарова М. Н. Сыда V - могильник окуневской культуры // Окуневский сборник 2. СПб., 2006. С. 53-59.
- Козинцев А. Г. Происхождение окуневского населения Южной Сибири по данным физической антропологии и генетики // Археология, этнография и антропология Евразии. 2020. Т. 48, № 4. С. 135-145. DOI 10.17746/1563-0102.2020.4S.4.135-145
- Лазаретов И. П. Окуневские могильники в долине р. Уйбат // Окуневский сборник. СПб.: Петро-РИФ, 1997. С. 19-64.
- Максименков Г. А. Окуневская культура в Южной Сибири // МИА. 1965. № 130. С. 168174.
- Максименков Г. А. Окуневская культура: Автореф. дис. ... д-ра ист. наук. Новосибирск, 1975. 39 с.
- Молодин В. И. Окуневская культура и сейминско-турбинский феномен (историографические заметки) // Археологические вести. 2022. № 34. С. 123-129.
- Молодин В. И., Пилипенко А. С., Чикишева Т. А., Ромащенко А. Г., Журавлев А. А., Поздняков Д. В., Трапезов Р. О. Мультидисциплинарные исследования населения Барабинской лесостепи V-I тыс. до н. э.: археологический, палеогенетический и антропологический аспекты. Новосибирск: Изд-во СО РАН, 2013. 220 с.
- Пилипенко А. С., Ромащенко А. Г., Молодин В. И., Куликов И. В., Кобзев В. Ф., Поздняков Д. В., Новикова О. И. Особенности захоронения младенцев в жилищах городища Чича-I Барабинской лесостепи по данным анализа структуры ДНК // Археология, этнография и антропология Евразии. 2008. №. 2. С. 57-67.
- Пилипенко А. С., Молодин В. И., Трапезов Р. О., Черданцев С. В., Журавлев А. А. Молекулярно-генетический анализ останков людей из погребального комплекса эпохи бронзы Бертек-56 (II тысячелетие до н. э., Республика Алтай, Россия) // Археология, этнография и антропология Евразии. 2016. № 4. С. 141-149.
- Поляков А. В. Обзор результатов начального этапа палеогенетических исследований населения эпохи бронзы Минусинских котловин // Теория и практика археологических ис-следований.2019.№ 2. С. 91-10S.
- Поляков А. В. Хронология и культурогенез памятников эпохи палеометалла Минусинских котловин. СПб.: ИИМК РАН, 2022. 364 с.
- Теплоухов С. А. Опыт классификации древних металлических культур Минусинского края // Материалы по этнографии. 1929. Т. 4, вып. 2. C. 41-62.
- Хохлов А. А., Солодовников К. Н., Рыкун М. П., Кравченко Г. Г., Китов Е. П. Краниологические данные к проблеме связи популяций ямной и афанасьевской культур Евразии начального этапа бронзового века // Вестник археологии, антропологии и этнографии. 2016. № 3. С. 86-106.
- Allentoft M. E., Sikora M., Sjögren K. G., Rasmussen S., Rasmussen M., Stenderup J., Damgaard P. B., Schroeder H., Ahlström T., Vinner L., Malaspinas A.-S., Marga-ryan A., Higham T., Chivall D., Lynnerup N., Harvig L., Baron J., Casa P. D., D^brow-ski P., Duffy P. R., Ebel A. V., Epimakhov A., Frei K., Furmanek M., Gralak T., Gro-mov A., Gronkiewicz S., Grupe G., Hajdu T., Jarysz R., Khartanovich V., Khokhlov A., Kiss V., Kolár J., Kriiska A., Lasak I., Longhi C., McGlynn G., Merkevicius A., Merky-te I., Metspalu M., Mkrtchyan R., Moiseyev V., Paja L., Pálfi G., Pokutta D., Pospiesz-ny L., Price T. D., Saag L., Sablin M., Shishlina N., Smrcka V., Soenov V. I., Szeve-rényi V., Tóth G., Trifanova S. V., Varul L., Vicze M., Yepiskoposyan L., Zhitenev V., Orlando L., Sicheritz Pontén T., Brunak S., Nielsen R., Kristiansen K., Willerslev E. Population genomics of Bronze Age Eurasia. Nature, 2015, vol. 522, pp. 167-172.
- Damgaard P., Martiniano R., Kamm J., Moreno-Mayar J. V., Kroonen G., Peyrot M., Barjamovic G., Rasmussen S., Zacho C., Baimukhanov N., Zaibert V., Merz V., Bid-danda A., Merz I., Loman V., Evdokimov V., Usmanova E., Hemphill B., Seguin Orlando A., Yediay F. E., Ullah I., Sjögren K.-G., Iversen K. H., Choin J., Fuente C. de la, Ilardo M., Schroeder H., Moiseyev V., Gromov A., Polyakov A., Omura S., Senyurt S. Y., Ahmad H., McKenzie C., Margaryan A., Hameed A., Samad A., Gul N., Khokhar M. H., Goriunova O. I., Bazaliiskii V. I., Novembre J., Weber A. W., Orlando L., Allen-toft M. E., Nielsen R., Kristiansen K., Sikora M., Outram A. K., Durbin R., Willerslev E. The First Horse Herders and the Impact of Early Bronze Age Steppe Expansions into Asia. Science, 2018, vol. 360, iss. 6396: eaar7711.
- Haak W., Forster P., Bramanti B., Matsumura S., Brandt G., Tanzer M., Villems R., Renfrew C., Gronenborn D., Werner A. K., Burger J. Ancient DNA from the first European farmers in 7500-Year-Old Neolithic sites. Science, 2005, vol. 305, pp. 1016-1018.
- Hollard C., Zvenigorosky V., Kovalev A., Kiryushin Y., Tishkin A., Lazaretov I., Crubezy E., Ludes B., Keyser C. New genetic evidence of affinities and discontinuities between bronze age Siberian populations. American Journal of Physical Anthropology, 2018, no. 167, pp. 97107.
- Molodin V. I., Pilipenko A. S., Romaschenko A. G., Zhuravlev A. A., Trapezov R. O., Chi-kisheva T. A., Pozdnyakov D. V. Human migrations in the southern region of the West Siberian Plain during the Bronze Age: Archaeological, palaeogenetic and anthropological data. In: Population Dynamics in Pre- and Early History: New Approaches Using Stable Isotopes and Genetics. Berlin, 2012, pp. 95-113.
- Narasimhan V., Patterson N., Moorjani P., Lazaridis I., Mark L., Mallick S., Rohland N., Bernardos R., Kim A., Nakatsuka N., Olalde I., Coppa A., Mallory J., Moiseyev V., Monge J., Olivieri L., Adamski N., Broomandkhoshbacht N., Candilio F., Cheronet O., Culle-ton B., Ferry M., Fernandes D., Gamarra B., Gaudio D., Hajdinjak M., Harney E., Harper T., Keating D., Lawson A.-M., Michel M., Novak M., Oppenheimer J., Rai N., Sirak K., Slon V., Stewardson K., Zhang Z., Akhatov G., Bagashev A., Baitanayev B., Bonora G., Chikisheva T., Derevianko A., Enshin D., Douka K., Dubova N., Epima-khov A., Freilich S., Fuller D., Goryachev A., Gromov A., Hanks B., Judd M., Kazizov E., Khokhlov A., Kitov E., Kupriyanova E., Kuznetsov P., Luiselli D., Maksudov F., Meikle-john C., Merrett D., Micheli R., Mochalov O., Muhammed Z., Mustafakulov S., Na-yak A., Rykun M., Pettner D., Potts R., Razhev D., Sarno S., Sikhymbaevae K., Slepchen-ko S., Stepanova N., Svyatko S., Vasilyev S., Vidale M., Voyakin D., Yermolayeva A., Zubova A., Shinde V., Lalueza-Fox C., Meyer M., Anthony D., Boivin N., Thangaraj K., Kennett D., Frachetti M., Pinhasi R., Reich D. The Formation of human populations in South and Central Asia. Science, 2019, vol. 365, iss. 6457 (eaat7487). DOI 10.1126/ science.aat7487
- Petkovski E., Keyser-Tracqui C., Crubezy C. MALDI-TOF MS analysis of Y-SNPs in ancient samples. Int. Congr Ser., 2006, vol. 1288, pp. 25-27.
- Pilipenko A. S., Trapezov R. O., Cherdantsev S. V., Babenko V. N., Nesterova M. S., Pozd-nyakov D. V., Molodin V. I., Polosmak N. V. Maternal genetic features of the Iron Age Tagar population from Southern Siberia (1st millennium BC). PLoS ONE, 2018. DOI 10.1371/ journal.pone.0204062.
- Pilipenko A. S., Trapezov R. O., Zhuravlev A. A., Molodin V. I., Romaschenko A. G. MtDNA Haplogroup A10 Lineages in Bronze Age Samples Suggest That Ancient Autochthonous Human Groups Contributed to the Specificity of the Indigenous West Siberian Population. PLoS ONE, 2015, vol. 10 (5), p. e0127182.
- Poliakov A., Svyatko S., Stepanova N. A review of the radiocarbon dates for the Afanasyevo Culture (Central Asia): Shifting towards the "shorter" chronology. Radiocarbon, 2019, vol. 61, iss. 1, pp. 243-263.
- Rasmussen M., Anzick S. L., Waters M. R., Skoglund P., DeGiorgio M., Stafford T. W. Jr., Rasmussen S., Moltke I., Albrechtsen A, Doyle S. M., Poznik G. D., Gudmundsdottir V., Yadav R., Malaspinas A. S., White S. S. 5th, Allentoft M. E., Cornejo O. E., Tambets K., Eriksson A., Heintzman P. D., Karmin M., Korneliussen T. S., Meltzer D. J., Pierre T. L.,
- Stenderup J., Saag L., Warmuth V. M., Lopes M. C., Malhi R. S., Brunak S., Sicheritz-Ponten T., Barnes I., Collins M., Orlando L., Balloux F., Manica A., Gupta R., Metspa-lu M., Bustamante C. D., Jakobsson M., Nielsen R., Willerslev E. The genome of a late Pleistocene human from a Clovis. Nature, 2014, vol. 506, no. 7487, pp. 225-229.
- Wilde S., Timpson A., Kirsanow K., Kaiser E., Kayser M., Unterländer M., Hollfelder N., Po-tekhina I. D., Schier W., Thomas M. G., Burger J. Direct evidence for positive selection of skin, hair, and eye pigmentation in Europeans during the last 5,000 y. Proc. Natl. Acad. Sci., 2014, vol. 111, no. 13, pp. 4832-4837.
- Zegura S. L., Karafet T. M., Zhivotovsky L. A., Hammer M. F. High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of Native American Y chromosomes into the Americas. Mol. Biol. Evol, 2004, vol. 21 (1), pp. 164-175. DOI 10.1093/molbev/msh009
