Разнообразие Y-хромосомы сибирских татар из позднесредневекового могильника Абрамово-10 (Барабинская лесостепь)
")
Автор: Трапезов Р.О., Пилипенко И.В., Федосеева Е.И., Черданцев С.В., Журавлев А.А., Поздняков Д.В., Томилин М.А., Рыкун М.П., Нестерова М.С., Молодин В.И., Пилипенко А.С.
Журнал: Вестник Новосибирского государственного университета. Серия: История, филология @historyphilology
Рубрика: Археология Евразии
Статья в выпуске: 5 т.25, 2026 года.
Бесплатный доступ
Представлены первые данные о мужском генофонде позднесредневекового населения западносибирской лесостепи. Исследована структура 18 образцов Y-хромосомы из могильника сибирских татар Абрамово-10 (XVI–XVIII вв. н. э.). В составе серии выявлено три филогенетических кластера: N-VL67/N1a2CTS6380>> VL67 (N = 13), Q-L56/Q-M346/Q1a2 (N = 4) и R-M198/R1a-M198 (N = 1). Основу мужского генофонда исследуемой группы составляют варианты, проникшие в регион в процессе его тюркизации на протяжении эпохи Средневековья, и более древние для региона компоненты, общие для этнокультурно контрастных групп населения Западной Сибири. Близкая к современной филогеографическая структура мужского населения сформировалась раньше, чем особенности генофонда мтДНК. В период миграции в регион европеоидного населения основные особенности мужского генофонда сибирских татар Барабы сохранились.
Сибирские татары, Барабинская лесостепь, позднее Средневековье, палеогенетика, Y-хромосома, этногенетические реконструкции
Короткий адрес: https://sciup.org/147253847
IDR: 147253847 | УДК: 572, 575 | DOI: 10.25205/1818-7919-2026-25-5-127-147
Y-Chromosome Diversity within Siberian Tatars from the Late Medieval Burial Ground Abramovo-10 (Baraba Forest-Steppe)
Purpose. The paper continues a series of articles devoted to the paleogenetic study of the Late Medieval populationfrom the West Siberian forest-steppe zone. The aim of the study is to obtain and analyze the first data on the Y-chromosome lineage diversity of the population that formed the Abramovo-10 burial ground in the central part of the Baraba forest-steppe (16th – 18th centuries AD). Results. Data on the structure of 18 Y-chromosome samples (male individuals) from the burial ground of the SiberianTatars Abramovo-10 were obtained. Phylogenetic and phylogeographic analysis of the studied lineages was carried out. Three phylogenetic clusters were identified in the studied series: N-VL67/N1a2CTS6380>>VL67 (N = 13, or ~ 72 % of the sample), Q-L56/Q-M346/Q1a2 (N = 4, or ~ 22 %) и R-M198/R1a-M198 (N = 1, or 5.5 %). Conclusion. The obtained results suggest that the basis of the male gene pool in the studied group consists of Y-chromosome lineages that spread across the region during the Turkification in the Middle Ages, as well as oldercomponents common to ethno-culturally contrasting populations of the Western Siberia. The phylogeographicstructure of the Y-chromosome pool, which is close to the modern one, was formed earlier than the features of the mtDNA gene pool, apparently due to the patrilocality of the studied group. During the period of the intensive Caucasianpopulation migration to the region, the main features of the male gene pool remained almost unchanged for significantpart of the Baraba Siberian Tatars.
Текст научной статьи Разнообразие Y-хромосомы сибирских татар из позднесредневекового могильника Абрамово-10 (Барабинская лесостепь)
В лесостепной зоне Западной Сибири и в прилегающей полосе южной тайги в эпоху Средневековья происходят интенсивные этногенетические процессы при участии автохтонных для данного региона групп угорского и самодийского происхождения, подвергавшихся воздействию мигрировавших на эту территорию тюркских популяций [Молодин и др., 1990]. В результате взаимодействия этих популяций в регионе формируется сложная структура современного коренного населения, относящегося к нескольким этническим образованиям. Очевидно, что формируемые популяции отличаются составом и относительным вкладом различных этнокультурных компонентов, что проявляется в наличии многочисленных локально-территориальных групп населения со своей спецификой на уровне материальной культуры и структуры генофонда.
Непосредственно в лесостепной зоне происходит сложение групп тюркоязычного населения, в совокупности рассматриваемого специалистами под названием «сибирские татары» [Валеев, 1993]. Большинство специалистов считает, что основные этапы формирования этнических групп в регионе завершаются к XIII–XIV вв. [Соболев, 1984]. Для более ранних археологических материалов этническая атрибуция, как правило, затруднительна. Исследование многочисленных погребальных памятников, связанных с различными группами сибирских татар, позволяет проводить их анализ методами физической антропологии, а в последнее время и палеогенетическими инструментами.
Крупные могильники исследованы на территории современного ареала тоболо-иртыш-ских, томских и барабинских сибирских татар [Валеев, 1993; Валеев, Томилов, 1996; Томилов, 1992; История Сибири, 2019]. Традиционно большинство из них ассоциируется с группой, проживающей в настоящее время на территории расположения могильника. Центральная часть Барабинской лесостепи, в которой расположен могильник Абрамово-10, входит в состав ареала современных барабинских татар.
В антропологических исследованиях [Багашёв, 2017] серия из Абрамово-10 традиционно используется в качестве позднесредневековой выборки этой группы сибирских татар (хотя не все исследователи согласны с включением барабинцев в общность сибирских татар [Токарев, 1958; Селезнёв, 1994]). Могильник, являясь самым крупным позднесредневековым погребальным памятником тюрков для рассматриваемого региона (всего раскопано 118 погребений), представляет несомненный интерес и для популяционно-палеогенетических исследований.
Авторами данной статьи ранее был проведен анализ репрезентативной серии из 72 образцов митохондриальной ДНК (мтДНК) из Абрамово-10 [Черданцев и др., 2025]. Данная работа продолжает серию статей, посвященных палеогенетическому анализу материалов могильника и генофонда сибирских татар в целом. Представлены результаты анализа серии ( N = 20) образцов Y-хромосомы из могильника Абрамово-10. Отметим, что это первая работа, посвященная анализу разнообразия Y-хромосомы в генофонде средневекового населения лесостепной полосы Западной Сибири.
Материалы и методы
Палеоантропологические материалы, исследованные в работе. Для ранее опубликованной нами работы по анализу разнообразия мтДНК сибирских татар из могильника Абрамо-во-10 [Черданцев и др., 2025] нами были использованы образцы от 74 индивидов различного пола и возраста (предварительные половозрастные определения были выполнены методами физической антропологии по морфологическим критериям черепов). Материалы были получены из краниологической коллекции кабинета антропологии ТГУ (73 образца) и ИАЭТ СО РАН (1 образец) [Там же]. Во всех случаях в качестве образцов были отобраны зубы высокой степени макроскопической сохранности.
Предварительную обработку палеоантропологического материала и получение образцов суммарной ДНК осуществляли с помощью методов, детально описанных в наших предшествующих работах [Pilipenko et al., 2015; 2018]. Поверхность зуба механически очищали от загрязнений, выдерживали зуб в растворе гипохлорита натрия, облучали ультрафиолетом и размалывали с помощью шаровой мельницы Retsch Mixer Mill MM200 (Германия). Материал зубов декальцинировали с помощью 0,5М ЭДТА с последующим лизисом протеиназой K (Сибэнзим, Россия). ДНК выделяли методом фенол-хлороформной экстракции.
Для анализа маркеров Y-хромосомы использовали только те образцы, для которых: 1) ранее были получены результаты по мтДНК (72 из 74 индивидов) [Черданцев и др., 2025]; 2) методами физической антропологии был предварительно установлен мужской пол. Таких образцов в серии оказалось 24. Для них проводили анализ маркеров половой принадлежности и аллельного профиля аутосомных STR с целью подтверждения корректности палеоантропологических данных о половой принадлежности останков и оценки степени сохранности ядерной ДНК. Для этого использовали набор COrDIS Expert26 (Гордиз, Россия). Процедуру проводили согласно инструкции производителя. Из 24 индивидов мужской пол и относительно высокая сохранность ядерной ДНК были подтверждены для 20 (см. табл. 1).
Таблица 1
Выборка палеоантропологических образцов из позднесредневекового могильника Абрамово-10, использованная для исследования разнообразия Y-хромосомы
Table 1
Description of paleoanthropological samples from the Late Medieval burial ground Abramovo-10, used to study the diversity of the Y chromosome
|
№ п/п |
Инвентарный номер |
Лабораторный шифр индивида |
№ п/п |
Инвентарный номер |
Лабораторный шифр индивида |
|
1 |
3932 |
Abr3 |
11 |
4001 |
Abr41 |
|
2 |
3973 |
Abr6 |
12 |
3958 |
Abr42 |
|
3 |
4000 |
Abr13 |
13 |
3970 |
Abr43 |
|
4 |
3992 |
Abr18 |
14 |
3960 |
Abr44 |
|
5 |
3994 |
Abr20 |
15 |
3936 |
Abr56 |
|
6 |
3937 |
Abr21 |
16 |
4004 |
Abr60 |
|
7 |
3949 |
Abr25 |
17 |
4003 |
Abr62 |
|
8 |
3959 |
Abr27 |
18 |
3939 |
Abr67 |
|
9 |
4002 |
Abr35 |
19 |
3047 |
Abr70 |
|
10 |
3975 |
Abr38 |
20 |
32/18Б |
Abr74 |
Анализ аллельного профиля YSTR проводили с помощью набора COrDIS Ystr (Гордиз, Россия) согласно инструкции производителя. Фрагментный анализ результатов мультиплексной ПЦР проводили на приборах НАНОФОР05 в ЦКП Генетического анализа ИЦиГ СО РАН.
Анализ аллельного профиля осуществляли с помощью программы GeneMarker (Soft-Genetics). Определение филогенетического положения исследованных структурных вариантов Y-хромосомы осуществляли с помощью программы-предиктора на сайте компании NEVGEN (, находящейся в открытом доступе. При филогенетическом / филогео-графическом анализе результатов в качестве базовой использовали классификацию кластеров Y-хромосомы, реализованную в рамках открытого онлайн-ресурса «The Universal Y-SNP Database» (UYSD v. 1.1) [Ralf et al., 2025], в рамках которой предложено унифицированное филогенетическое дерево вариантов Y-хромосомы человека с соответствующей единой номенклатурой кластеров (. При изложении полученных результатов также приводили варианты названий филогенетических кластеров, предлагаемые программой-предиктором, и ранее использовавшиеся варианты их наименования.
Все работы с древним материалом выполнены на базе специализированной палеогенети-ческой инфраструктуры межинститутской лаборатории молекулярной палеогенетики и палеогеномики ИЦИГ СО РАН (Новосибирск, Россия), которая отвечает всем современным требованиям, предъявляемым к палеогенетическим лабораториям. Меры против контаминации и процедуры верификации результатов описаны в наших предшествующих статьях [Pilipenko et al., 2015; 2018]. Условия проведения исследования, используемые протоколы работы и особенности полученных результатов однозначно свидетельствуют об их высокой достоверности.
Результаты исследования
Проведенное нами ранее исследование серии образцов мтДНК из могильника Абрамо-во-10 [Черданцев и др., 2025] продемонстрировало очень высокий уровень сохранности мтДНК в антропологическом материале, что позволило получить достоверные данные для 72 из 74 индивидов, первоначально задействованных в исследовании. Сохранность ядерной ДНК в останках обычно уступает сохранности мтДНК, что в очередной раз подтвердилось в нашем исследовании. Из 24 индивидов потенциально мужского пола (по антропологическим данным) в четырех образцах сохранность ядерной ДНК не позволила выполнить анализ аллельного профиля STR-локусов (аутосомных и Y-хромосомы). Эти образцы были исключены из дальнейшего исследования. Для оставшихся 20 индивидов были получены полные или частичные аллельные профили 18 STR-локусов Y-хромоcомы (табл. 2).
Для всех 20 индивидов программа-предиктор с высокой вероятностью определяет филогенетическое положение вариантов Y-хромосомы. Тем не менее, мы исключили из дальнейшего рассмотрения два образца, характеризующихся наименее полными аллельными профилями: образец Abr3, для которого был определен статус 10 из 18 YSTR, и Abr70 (успешный анализ лишь 7 из 18 YSTR). Реконструированные для данных индивидов варианты Y-хромосомы были отнесены программой-предиктором к определенным филогенетическим кластерам (см. табл. 2), но вероятность корректности предсказания гаплогруппы для них была ниже, чем для остальных образцов.
Для 18 образцов численность успешно генотипированных YSTR варьировала от 14 до 18. Для 8 из 18 индивидов нам удалось определить полный аллельный профиль всего набора из 18 YSTR, анализируемого используемым нами набором реактивов. Необходимо отметить, что для палеогенетического исследования серия из 18 образцов Y-хромосомы, полученных от индивидов из одного археологического памятника, является высоко репрезентативной. На данный момент намного чаще в литературе встречается ситуация, когда из конкретного археологического памятника происходят лишь единичные успешно исследованные образцы Y-хромосомы.
Таблица 2
Результаты генотипирования Y-STR локусов в образцах из могильника Абрамово-10
Results of genotyping of Y-STR loci in samples from the Abramovo-10 burial ground
Table 2
|
о и о R Pi сл |
О', СП ОП ^ Q |
НН О 00 (^ оо ^ Q |
о > Q |
00 Q |
00 Q |
со 00 ^ Q |
м о СП Q |
Q |
чо 1Л ^ Q |
00 СП и Q |
о 04 СП 00 ^ Q |
04 Q |
оо > Q |
40 1П (Л Q |
04 СП 00 Q |
in 00 (Z) > Q |
m СП 40 00 ^ Q |
Гаплогруппа (согласно NEVGEN) |
|
Abr3 |
10 |
13 |
28 |
10 |
23 |
26 |
15 |
10 |
12/13 |
N1a2CTS6380>>VL67 |
||||||||
|
Abr6 |
10 |
13 |
14 |
13 |
28 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
|
Abr13 |
10 |
13 |
14 |
14 |
13 |
14 |
27 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
N1a2CTS6380>>VL67 |
||
|
Abr18 |
11 |
13 |
13 |
13 |
13 |
14 |
25 |
21 |
11 |
23 |
29 |
13 |
15/17 |
Q-M346 |
||||
|
Abr20 |
10 |
13 |
14 |
13 |
14 |
28 |
10 |
23 |
26 |
19 |
15 |
10 |
12/ |
21 |
N1a2CTS6380>>VL67 |
|||
|
Abr21 |
10 |
13 |
13 |
13 |
31 |
13 |
14 |
25 |
11 |
23 |
29 |
13 |
15/17 |
23 |
Q-M346 |
|||
|
Abr25 |
10 |
14 |
13 |
14 |
14 |
14 |
25 |
17 |
11 |
24 |
31 |
17 |
12 |
13/17 |
22 |
Q-M346 |
||
|
Abr27 |
10 |
13 |
14 |
14 |
33 |
13 |
14 |
27 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
|
Abr35 |
10 |
13 |
14 |
14 |
28 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/15 |
24 |
N1a2CTS6380>>VL67 |
|
Abr38 |
10 |
14 |
16 |
14 |
31 |
13 |
11 |
24 |
19 |
11 |
25 |
32 |
20 |
16 |
11 |
11/14 |
23 |
R1a-M198 |
|
Abr41 |
10 |
13 |
14 |
14 |
33 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
|
Abr42 |
10 |
13 |
14 |
14 |
28 |
13 |
14 |
17 |
10 |
23 |
26 |
15 |
10 |
12/13 |
N1a2CTS6380>>VL67 |
|||
|
Abr43 |
10 |
13 |
14 |
14 |
28 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
|
Abr44 |
10 |
13 |
16 |
14 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
20 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
||
|
Abr56 |
10 |
13 |
14 |
14 |
28 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
|
Abr60 |
10 |
15 |
14 |
14 |
14 |
25 |
11 |
24 |
31 |
19 |
17 |
12 |
11/17 |
22 |
Q-M346 |
|||
|
Abr62 |
10 |
13 |
14 |
14 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
|
|
Abr67 |
10 |
13 |
14 |
14 |
28 |
13 |
14 |
28 |
17 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
24 |
N1a2CTS6380>>VL67 |
|
Abr70 |
10 |
14 |
11 |
24 |
32 |
12 |
/17 |
Q-M346 |
||||||||||
|
Abr74 |
10 |
13 |
14 |
28 |
13 |
14 |
28 |
10 |
23 |
26 |
19 |
15 |
10 |
12/13 |
N1a2CTS6380>>VL67 |
Среди 18 образцов выявлено три филогенетических кластера (в порядке убывания частоты встречаемости): N1a2CTS6380 (по NEVGEN) или N-Y3037 (по UYSD) – 13 из 18 индивидов или ~ 72 % выборки; Q-M346 (по NEVGEN и UYSD, ранее – кластер Q1a2(Q1a2)) – 4 носителя, ~ 22 % выборки; R1a-M198 (по NEVGEN) или R-M198 (по UYSD) – 1 носитель, ~ 5,5 % выборки (см. табл. 2, рис. 1). При численности выборки N = 18 мы, безусловно, не можем утверждать, что зафиксированы все компоненты мужского генофонда популяции, сформировавшей могильник Абрамово-10. Однако состав серии уверенно свидетельствует о высокой роли двух кластеров – N-Y3037 и Q-M346, первый из которых, с высокой вероятностью, является доминирующим в генофонде рассматриваемой группы позднесредневекового населения.
При рассмотрении серии образцов, происходящих из одной локальной группы или могильника, всегда стоит вопрос, не являются ли обнаруженные особенности выборки, такие как повышенная частота одного или нескольких компонентов генофонда, следствием попадания в выборку большого числа близких или даже прямых родственников. Палеогенетиче-ское исследование более уязвимо в данном отношении по сравнению с исследованиями современных выборок, из которых близкие родственники исключаются по результатам анкетирования доноров образцов. В нашей выборке были выявлены носители идентичных аллельных профилей YSTR: например, среди носителей наиболее представленного кластера (N-Y3037) идентичные профили имели 3 из 7 индивидов, для которых был установлен статус всех 18 STR-локусов (рис. 2). Этот факт увеличивает вероятность влияния близкого родства индивидов на полученную картину разнообразия вариантов Y-хромосомы, в частности представленность кластера N-Y3037, оцененная на основе нашей выборки из Абрамово-10 на уровне ~ 72 %, может быть несколько завышенной по сравнению с генофондом популяции в целом. Следует учитывать, что присутствие в локальных популяциях большого числа индивидов с идентичными YSTR-профилями, по-видимому, является характерной чертой многих коренных популяций южных районов Сибири, включая современные тюркоязычные популяции региона, например хакасов и шорцев [Харьков и др., 2020].
Также мы можем провести параллели с результатами, полученными при исследовании разнообразия мтДНК в серии из могильника Абрамово-10 [Черданцев и др., 2025]. В исследованной нами ранее выборке мтДНК также с высокой частотой присутствовали отдельные
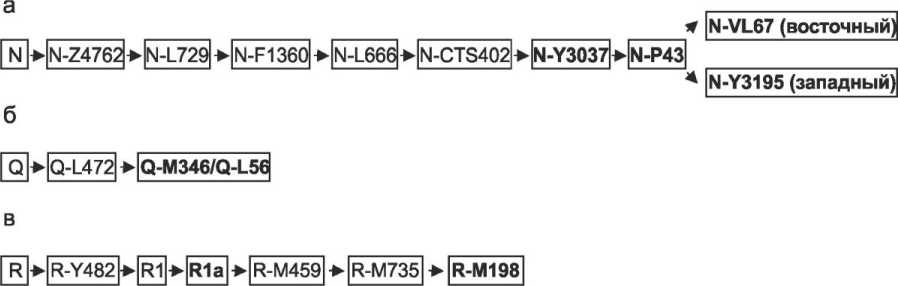
Рис. 1. Филогенетическое положение вариантов Y-хромосомы, выявленных в составе серии образцов из позднесредневекового могильника Абрамово-10 (в соответствии с классификацией кластеров Y-хромосомы, реализованной в рамках «The Universal Y-SNP Database» (UYSD v. 1.1))
Fig. 1. The phylogenetic position of Y-chromosome lineages identified in a series of samples from the Late Medieval Abramovo-10 burial ground (in accordance with the classification of Y-chromosome clusters implemented in the framework of The Universal Y-SNP Database (UYSD v. 1.1))

Археология Евразии
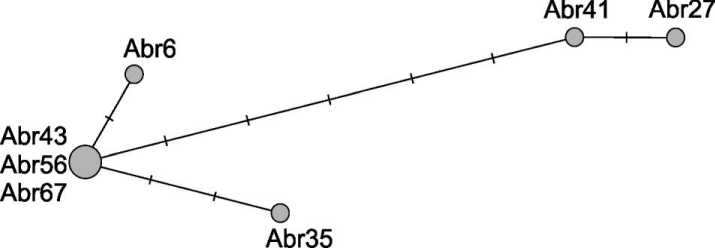
Рис. 2. Медианная сеть вариантов гаплогруппы N-VL67, выявленных в серии образцов Y-хромосомы из позднесредневекового могильника Абрамово-10 (представлены только образцы, для которых получен полный аллельный профиль 18 YSTR-маркеров)
Fig. 2. The median network of haplogroup N-VL67 variants identified in a series of Y chromosome samples from the Late Medieval Abramovo-10 burial ground (only samples with a complete profile of 18 YSTR markers are presented)
варианты гаплогрупп H, U4, C5b1. Однако наличие таких доминирующих вариантов было характерно для средневековых популяций в целом (что устанавливается при анализе сборных серий из ряда могильников, неопубликованные данные авторов), как и для современных популяционных выборок сибирских татар, из которых близкие родственники исключены [Naumova et al., 2008; Gubina et al., 2018]. Мы считаем, что присутствие идентичных вариантов мтДНК и Y-хромосомы является распространенной чертой генофонда мтДНК и Y-хромосомы позднесредневековых популяций региона.
Рассмотрим подробнее филогению и филогеографию кластеров, выявленных в составе исследованной серии из Абрамово-10. Данные о составе мужского генофонда барабинских татар пока не были полноценно опубликованы в научной печати. Однако сибирские татары Ба-рабы были использованы в качестве сравнительной серии в работе, посвященной генофонду Y-хромосомы западносибирских угров (хантов и манси) [Пономарев и др., 2024, рис. 2]. Это позволяет приблизительно оценить параметры мужского генофонда современных барабин-цев и предварительно сравнить их с серией из Абрамово-10. Так, в генофонде современных барабинских татар присутствуют все три кластера, выявленные нами в данной работе: N-Y3037, Q-M346/Q-L56, R-M198. Как и в серии из Абрамово-10 кластеры N-Y3037, Q-M346 составляют основу современного генофонда. Однако их соотношение иное: N-Y3037, составляющая ~ 72 % нашей серии, у современных барабинских татар представлена с частотой всего около 20 %, а общая частота вариантов гаплогруппы Q-M346/Q-L56, напротив, выше у современных барабинцев (~ 45 %), чем в Абрамово-10 (22 %). В современной выборке присутствует и кластер R-M198 (~ 6,5 %), т. е. он представляет собой минорный компонент генофонда, имеющий частоту, сопоставимую со средневековой выборкой (~ 5,5 %). Отсутствие в нашей серии ряда других минорных компонентов, выявленных в современной популяции (R1b, E и др.), отчасти может быть связано с эффектом малой выборки либо объясняется их более поздним проникновением в генофонд населения региона. Таким образом, можно констатировать, что некоторые основные черты мужского генофонда сибирских татар Бара-бинской лесостепи (по крайней мере состав важнейших кластеров) в значительной степени были сформированы уже ко времени функционирования могильника Абрамово-10 (XVI– XVIII вв.). По-видимому, основные изменения впоследствии касались в первую очередь соотношения различных кластеров: в частности, мы фиксируем увеличение роли вариантов гаплогруппы Q-M346 за счет снижения доли вариантов гаплогруппы N-Y3037.
Все выявленные в Абрамово-10 варианты гаплогруппы N-Y3037/N1a2CTS6380 относятся к субкластеру N-P43, а точнее – к одной из двух основных ветвей N-P43 – гаплогруппе N-VL67 (см. табл. 2, рис. 1, а ). Этот кластер, наряду с N-Y3195, характеризуется хорошо вы-
раженной филогеографической спецификой, ранее уже отмеченной другими авторами [Пономарев и др., 2024]. Гаплогруппа N-P43 присутствует в мужском генофонде всех основных тюркоязычных и финно-угорских популяций Приуралья, Зауралья и Западной Сибири, но соотношение субкластеров N-VL67 и N-Y3195 меняется: по направлению с запада на восток уменьшается частота «западного» субкластера N-Y3195 и растет частота «восточного» N-VL67. Условной границей распространения этих двух кластеров является бассейн Иртыша на юге и Оби севернее устья Иртыша. Западнее этой границы доминирует N-Y3195, а восточнее преобладает N-VL67. Значительно западнее обозначенной границы варианты N-VL67 присутствуют с небольшой частотой в генофондах основных тюркоязычных групп, вплоть до Волго-Уральского региона [Пономарев и др., 2024], в то время как восточнее этой границы присутствуют только варианты кластера N-VL67 (т. е. N-Y3195 полностью отсутствует). В целом среди современных тюркоязычных и финно-угорских популяций наибольшей частоты интересующая нас гаплогруппа N-VL67 достигает в популяции хакасов [Харьков и др., 2020] и некоторых локальных группах хантов (наиболее восточных и южных) [Пономарев и др., 2024].
Отдельного внимания заслуживают самодийские народы Западной Сибири: в генофонде северных самодийцев отмечена наибольшая суммарная частота кластера N-P43, значительная часть которого представлена именно вариантами N-VL67 [Харьков и др., 2021; 2025; Rootsi et al., 2007]. Из-за столь высоких частот у северных самодийцев кластер N-P43 (и N-VL67, в частности) часто рассматривают как генетический компонент, связанный с самодийским вкладом в генофонд населения Западной и Южной Сибири (см., например, [Харьков и др., 2020]). На наш взгляд, его распространение в лесостепной и южно-таежной зонах Западной Сибири можно связывать с процессами тюркизации региона – распространения тюркоязычных групп населения в регионе, которое по археологическим данным наблюдается в различных районах Западной Сибири на протяжении большинства периодов эпохи Средневековья (с середины I тыс. н. э. до второй половины II тыс. н. э.).
Потенциальными источниками проникновения тюркоязычных групп в регион являются территория Алтае-Саянской горной системы и степные пространства, расположенные на территории современного Казахстана: в генофонде современного тюркоязычного населения Алтая и Саян [Харьков и др., 2020], а также у части родовых групп современных казахов Казахстана [Zhunussova et al., 2025] со значительной частотой представлен кластер N-VL67. О распространении с тюркоязычными группами свидетельствует и отмеченное выше наличие N-VL67 у тюркоязычных (но не у финно-угорских) популяций вплоть до Волго-Уральского региона на западе. Наиболее ранним из установленных в палеогенетических исследованиях носителей гаплогруппы N-VL67 является представитель пазырыкской культуры раннего железного века с территории Горного Алтая [Пилипенко и др., 2015]. Таким образом, данный кластер присутствовал в популяции Алтая за почти 1000 лет до начала процессов тюркизации населения Западной Сибири. Следует отметить, что с территорией северной части Алтае-Саянского нагорья часто связывают и прародину некоторых протосамодийских племен [Этногенез…, 1980; Народы Западной Сибири…, 2005]. Возможно, в процессе тюр-кизации происходило распространение тюркскими популяциями на север и северо-запад компонентов, имеющих в том числе протосамодийское происхождение.
Альтернативным сценарием могло бы быть проникновение вариантов N-VL67 в генофонд тюркоязычных популяций Западной Сибири (сибирских татар) в результате непосредственного контакта со средневековыми самодийскими группами. Однако, по нашим данным, в генофонде позднесредневековых южных самодийцев, которые, наиболее вероятно, контактировали с тюркоязычными группами – южных (нарымских) селькупов, варианты N-VL67 представлены с очень низкой частотой и только в могильниках, отражающих наибольшую интенсивность этнокультурных контактов с позднесредневековыми тюрками (сибирскими татарами, чулымцами и хакасами) (неопубликованные данные авторов).
Таким образом, мы считаем высокую частоту N-VL67, выявленную в генофонде группы сибирских татар, сформировавшей могильник Абрамово-10, следствием процессов тюркиза- ции региона в предшествующие формированию могильника периоды развитого и начала позднего Средневековья, а также вероятными контактами непосредственно в период формирования могильника с тюркоязычным населением Саяно-Алтайского региона, для которого характерна высокая представленность N-VL67 [Харьков и др., 2020]. В данном отношении результаты по мужскому генофонду хорошо коррелируют с результатами по мтДНК, фило-географический анализ которых свидетельствует об интенсивных контактах популяции из Абрамово-10 с тюркоязычными группами Алтая и Саян [Черданцев и др., 2025].
В то же время не все выводы, сделанные нами по мтДНК из Абрамово-10, коррелируют с данными по мужскому генофонду. Это касается результатов сравнения серии с другими группами современных сибирских татар, прежде всего тоболо-иртышскими [Агджоян и др., 2016], а также томскими [Валихова и др., 2022]. В генофонде томских татар линии гапло-группы N-Y3037 представлены крайне слабо: они были выявлены лишь в 4 из 167 исследованных образцов (в пос. Эушта и Черная речка) [Там же]. В контексте данной работы представляет интерес очень высокая частота вариантов гаплогруппы N-Y3037 в некоторых группах тоболо-иртышских татар, в частности у заболотных (ясколбинских) татар, в генофонде которых ее линии представлены с частотой более 80 % (т. е. сопоставимо или даже больше, чем в серии из Абрамово-10), хотя в других локальных группах частота намного ниже [Агджоян и др., 2016]. Ранее при анализе серии из Абрамово-10 на уровне мтДНК мы установили, что по структуре генофонда мтДНК эта группа ближе к сводной серии тоболо-иртышских татар [Naumova et al., 2008], чем к барабинским [Gubina et al., 2018]. Таким образом, и в данном случае можно было бы видеть определенную корреляцию между результатами анализа генофонда мтДНК и Y-хромосомы. Однако рассмотрение на уровне субкластеров гаплогруппы N-Y3037 опровергает такой вывод: высокая доля линий N-Y3037 в генофонде ясколбинских тоболо-иртышских татар полностью сформирована за счет вклада «западного» субкластера N-Y3195, отсутствующего в серии из Абрамово-10. В качестве минорного компонента «восточный» субкластер N-VL67, доминирующий в Абрамово-10, выявлен лишь у искеро-тобольской группы тоболо-иртышских татар, ареал которой вплотную примыкает к Барабе с запада [Пономарев и др., 2024]. Таким образом, мужская часть популяции, сформировавшей могильник Абрамово-10, демонстрирует явное сходство с барабин-скими, а не с тоболо-иртышскими татарами.
Вторым по значимости в исследованной серии образцов из Абрамово-10 является гапло-группа Q-M346/Q-L56/Q1a2 (ранее Q1a2; рис. 1, б ). Этот кластер является одним из наиболее распространенных компонентов мужского генофонда центральноазиатских популяций, включая современные популяции, населяющие Алтае-Саянский регион, а также Западной Сибири, где он широко представлен как в популяциях тюркоязычных групп (современные сибирские татары), так и у угров (ханты), а особенно кетов и ряда самодийцев. По-видимому, первичное распространение этого кластера по обширным территориям Южной и Западной Сибири происходило в результате древних миграций задолго до формирования современных этнокультурных групп региона. В результате присутствие данного компонента в той или иной степени было характерно для предковых групп всех основных этнокультурных образований региона. В современных популяциях различной этнической принадлежности выявляются субкластеры Q-M346/Q-L56, демонстрирующие этноспецифичность [Харьков и др., 2013; 2020; 2023]. Для прояснения вопросов происхождения вариантов гаплогруппы Q-M346/Q-L56, обнаруженных в составе серии из Абрамово-10, необходимо проверить их близость к этноспецифическим субкластерам. Для этого необходим анализ статуса ряда ОНП Y-хромосомы и / или расширение анализируемого набора YSTR (эти задачи в настоящее время выполняются авторами). На данном этапе можно отметить, что присутствие значительной доли вариантов гаплогруппы Q-M346/Q-L56 характерно также и для современных сибирских татар лесостепной зоны Западной Сибири, включая как барабинские, так и тобо-ло-иртышские локальные группы [Агджоян и др., 2016; Пономарев и др., 2024].
Линии гаплогруппы R-M198/R1a-M198 (рис. 1, в ) распространены на территории лесостепной зоны Западной Сибири начиная с эпохи бронзы. Их первое появление в южных районах Западной и Южной Сибири, включая Барабинскую лесостепь, связано с миграционными волнами носителей андроновской (федоровской) культуры, происходившими в первой половине – середине II тысячелетия до н. э. [Keyser et al., 2009; Трапезов и др., 2020; 2022]. Впоследствии этот компонент сохранился в генофонде мужского населения южных районов Сибири. Непосредственно на территории Барабы он играет значимую роль в мужском генофонде носителей саргатской культуры раннего железного века [Пилипенко и др., 2017]. Таким образом, данный компонент мужского генофонда может быть связан с сохранением в структуре генофонда древних линий Y-хромосомы, присутствовавших в популяциях Бара-бинской лесостепи в периоды, предшествующие тюркизации населения. С другой стороны, распространенность вариантов R-M198/R1a-M198 в макрорегионе Западной и Южной Сибири к концу раннего железного века – начальным этапам Средневековья была очень широка. Поэтому нельзя исключить и более поздние повторные миграции его носителей из сопредельных с Барабой территорий. Установить время и механизм проникновения этого компонента в генофонд сибирских татар, сформировавших могильник Абрамово-10, потенциально позволит проводимый нами в настоящее время углубленный анализ данного образца Y-хромосомы по расширенному набору YSTR.
При рассмотрении этногенетических процессов, протекавших на юге Западной Сибири в период формирования могильника Абрамово-10 (XVI–XVIII вв. н. э.) и в последующее время, вплоть до современности, важным с точки зрения динамики генетического состава популяций становится фактор влияния на коренное население пришлых европеоидных групп. Состав исследованной нами мужской серии из Абрамово-10 свидетельствует, что проникавшее в регион пришлое европеоидное население в период функционирования погребального памятника не оказывало значимого влияния на генофонд Y-хромосомы: потенциально с такого рода влиянием можно связать лишь единственный вариант R-M198/R1a-M198 гаплогруппы, которая является доминирующей в русском (восточнославянском) мужском генофонде [Balanovsky et al., 2008]. Как отмечено выше, мы пока не можем исключить связь обнаруженного варианта с предшествующими популяциями Сибири, но, даже если этот компонент связан с пришлым европеоидным населением, он представляет собой лишь минорный компонент в генофонде популяции, оставившей могильник Абрамово-10. Что касается современных популяций сибирских татар из Барабинской лесостепи, то их генофонд содержит помимо R-M198/R1a-M198 еще ряд минорных кластеров (R1b-M73, I2a1-P37), однако даже суммарная частота кластеров, потенциально имеющих недавнее западно-евразийское происхождение, не превышает четвертой части всего состава мужского генофонда барабинцев [Пономарев и др., 2024]. Таким образом, можно констатировать, что, несмотря на некоторое увеличение разнообразия и доли западно-евразийских вариантов Y-хромосомы, барабинские татары сохранили основные черты структуры мужского генофонда, включая состав и общую долю доминирующих филогенетических кластеров (N-VL67 и Q-M346/Q-L56). Этот вывод хорошо коррелирует с полученным нами ранее заключением о сохранении значительной частью современных барабинских татар основных черт генофонда мтДНК [Черданцев и др., 2025]. При этом, как и в случае с мтДНК, необходимо учитывать, что при формировании выборок для популяционно-генетического исследования современного населения из их состава были исключены индивиды, являющиеся потомками недавних смешанных браков, что не позволяет распространить вывод о сохранении основных исконных черт структуры генофонда на всю современную популяцию барабинцев.
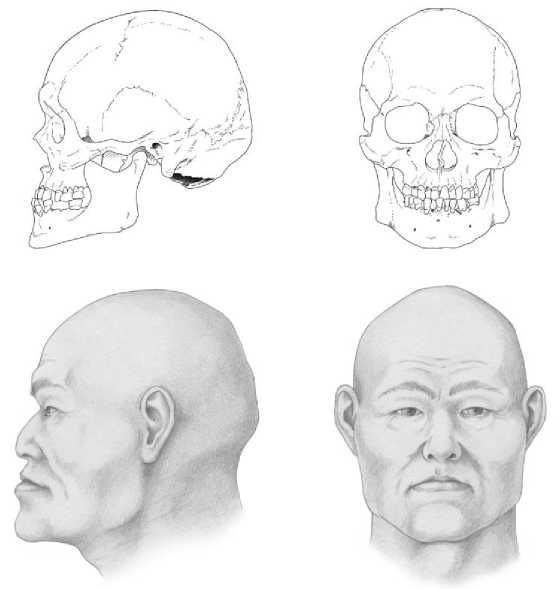
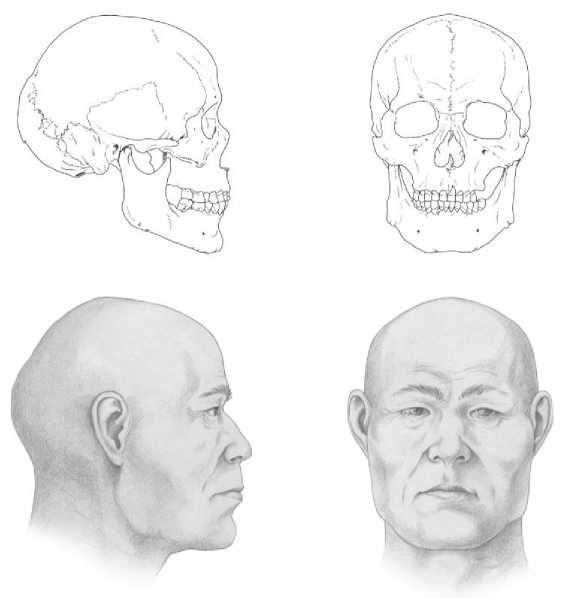
В рамках междисциплинарного исследования материалов могильника Абрамово-10 помимо палеогенетического анализа также выполнены графические портретные реконструкции ряда индивидов мужского пола из исследуемого памятника, которые входят в состав формируемой нами портретной галереи представителей разновременных групп средневекового населения Западной Сибири (рис. 3).
а
б
Рис. 3 . Портретная реконструкция индивидов из могильника Абрамово-10: а – мужчина, возраст 40–45 лет, погр. 2; б – мужчина, возраст 40–45 лет, погр. 3 Автор реконструкции канд. ист. наук Д. В. Поздняков
Fig. 3. Portrait reconstruction of individuals from the Abramovo-10 burial ground: a – male, 40–45 years old (burial 2); b – male, 40–45 years old (burial 3) The author of the reconstruction is Candidate of Historical Sciences D. V. Pozdnyakov
Заключение
Лесостепная и южно-таежная зоны Западной Сибири в эпоху Средневековья были охвачены сложными этногенетическими процессами, лежащими в основе формирования многокомпонентного современного коренного населения, включающего представителей нескольких культурно, лингвистически и генетически контрастных популяций – тюркских, угорских и самодийских. Объективная реконструкция этих процессов требует реализации комплексного междисциплинарного исследования всего корпуса источников, касающихся истории населения региона. Одним из наиболее перспективных подходов является популяционно-палео-генетическое исследование накопленных палеоантропологических материалов из разновременных могильников, ассоциированных с конкретными локальными популяциями. Сопоставление репрезентативных палеогенетических результатов с накопленными археологическими и палеоантропологическими данными о древнем населении, а также данными этногеномики о современных популяциях позволяют рассчитывать на объективную реконструкцию поздних этапов истории формирования населения Западной Сибири в ее этнокультурных и популяционно-генетических аспектах. К настоящему моменту препятствием для реализации такого рода исследовательской программы является слабый уровень исследованности средневековых популяций методами палеогенетики. Данная работа продолжила серию статей коллектива авторов, посвященную введению в научный оборот первых палеогенетических данных о средневековых популяциях лесостепной и южно-таежной зон Западной Сибири. Статья служит развитием опубликованной нами ранее работы о разнообразии мтДНК сибирских татар из позднесредневекового могильника Абрамово-10 – наиболее крупного исследованного археологами погребального памятника этого периода в западно-сибирской лесостепи. Полученные результаты – это первые в научной литературе данные о составе мужского генофонда позднесредневекового населения лесостепной зоны Западной Сибири.
Исследованная выборка ( N = 18) позволила определить состав основных компонентов мужского генофонда средневековых сибирских татар Барабы. Результаты свидетельствуют о возможном вкладе нескольких источников в формирование состава мужской части популяции. При этом, на наш взгляд, определяющую роль могли играть компоненты, связанные с многостадийными процессами тюркизации населения региона (т. е. компоненты, расселение которых связано с разновременными миграционными волнами тюркоязычных групп), а также сохранившиеся в составе генофонда древние компоненты, распространявшиеся в Западной Сибири в эпохи, предшествующие Средневековью, и присутствующие в качестве общих составляющих в предковых генофондах этнокультурно контрастных групп населения региона. В период формирования могильника Абрамово-10 (XVI–XVIII вв.) состав основных компонентов мужского генофонда сибирских татар уже сложился, и далее происходило в основном лишь изменение их соотношения. Оба генетических маркера с однородительским наследованием (мтДНК и Y-хромосома) свидетельствуют, что значительная часть популяции сибирских татар Барабинской лесостепи сохранила специфику своего генофонда в период массовой миграции в регион европеоидного населения в последние несколько веков. Вместе с тем выявлены и расхождения в выводах, полученных для материнского и мужского генофонда популяции, сформировавшей могильник Абрамово-10: в отличие от генофонда мтДНК, структура которого отражает большее сходство серии из могильника с современными тоболо-иртышскими татарами, серия Y-хромосомы однозначно сближается с барабинской группой сибирских татар и отличается от основных тоболо-иртышских локальных групп. На наш взгляд, этот результат не является противоречием, а скорее отражает особенности формирования женского и мужского генофонда, в частности выраженную патрилокальность населения, благодаря которой близкая к современной филогеографическая структура мужского населения формировалась раньше, чем особенности состава материнского генофонда (мтДНК), продолжавшие меняться на протяжении Нового времени.
Очевидно, что для более детальной интерпретации подобных результатов остро не хватает репрезентативных данных по генофонду других разновременных локально-территориальных групп средневекового населения Западной Сибири. Такие данные предполагается получить в рамках анализа обширной диахронной модели (выборки) населения лесостепной и южно-таежной зон Западной Сибири, выполняемого нами в настоящее время. Учитывая полученный нами объем первичных лабораторных результатов в этом направлении, мы полагаем, что в ближайшие годы западносибирская лесостепная и южно-таежная зоны могут стать одним из наиболее хорошо исследованных регионов в отношении вариабельности Y-хромосомы в разновременных популяциях. Другим перспективным направлением является углубленный анализ структуры мтДНК и Y-хромосомы, а также расширение спектра анализируемых ядерных маркеров.
