Разрешающая способность и оптическая чувствительность камерных глаз брюхоногих моллюсков

Автор: Шепелева Ирина Павловна
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Физиология растений и животных
Статья в выпуске: 4, 2011 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14719820
IDR: 14719820
Текст статьи Разрешающая способность и оптическая чувствительность камерных глаз брюхоногих моллюсков
Введение. Подавляющее большинство видов брюхоногих моллюсков имеет камерные глаза [31]. По устройству камерный глаз — потенциально хороший орган зрения. Он имеет лучшую разрешающую способность и оптическую чувствительность по } сравнению с другими типами глаз подобного размера благодаря длинному фокусному | расстоянию по отношению к диаметру фото-{ рецепторов и широкой апертуре, а также яв-| ное преимущество в преодолении главного § оптического дефекта — дифракции [42]. | Очевидно, что высокая разрешающая способность необходима хищникам, но хорошее | зрение так же важно другим животным для \ распознавания хищников, нахождения особей $ своего вида, маневрирования в местах обита-\ ния [36]. По сравнению с другими животными брюхоногие моллюски медлительны, к тому же части из них свойствен сумеречноночной тип активности. В связи с этим среди некоторых исследователей сенсорных органов брюхоногих моллюсков распространено мнение, что эти животные определяют объекты при помощи хеморецепции, а камерные глаза обеспечивают только реакцию фототаксиса [13; 17; 20 —21]. Однако морфологические и оптические исследования показали, что глаза некоторых видов брюхоногих моллюсков приспособлены к зрению [5 — 7; 11 — 12; 22; 26; 33 — 34]. Цель настоящего обзора — обобщить литературные и собственные данные морфологических и оптических исследований о функциональных возможностях камерных глаз брюхоногих моллюсков.
Разрешающая способность и оптическая чувствительность. Ключевыми параметрами, дающими представление о зрительной функции глаза, являются разрешающая способность и оптическая чувстви тельность. Они могут быть рассчитаны па основе анатомических и оптических данных [28].
Разрешающая способность глаза или острота зрения — это величина, обратная разрешаемому угловому расстоянию. Две близкие светящиеся точки глаз воспринимает раздельно, а не как одну слившуюся точку только при условии, что угловое расстояние между этими точками нс меньше определенного предела. Этот предел называется разрешаемым угловым расстоянием [4]. Острота зрения определяет возможность идентифицировать объекты на расстоянии и детали объектов вблизи [23]. Разрешающая способность зависит от диаметра фоторецепторов, плотности их расположения и фокусного расстояния оптической системы. Увеличение остроты зрения достигается уменьшением диаметра рецепторов до минимально возможного (примерно до 1 мкм) одновременно с увеличением плотности их расположения и/ или увеличением фокусного расстояния, что должно сопровождаться увеличением размера глаза. Разрешающая способность глаза (R, рад-1) для гексагонального расположения фоторецепторов рассчитывается по формуле [28]:
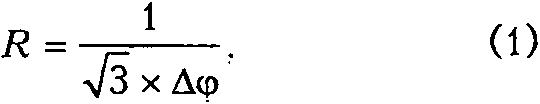
где Аф = у (град) — разрешаемое угловое расстояние между центрами соседних фоторецепторов относительно задней узловой точки оптической системы, р — анатомическое расстояние между центрами соседних фоторецепторов и f — фокусное расстояние оптической системы.
Оптическая чувствительность глаза, или чувствительность глаза к доступному свету, — это отношение количества фотонов, абсорбированных фоторецептором, к количеству фотонов, испускаемых с единицы площади протяженного источника света в телесном угле в один стерадиан. На чувствительность глаза к свету влияют диаметр апертуры, длина и диаметр фоторецепторов, а также фокусное расстояние оптической системы. Увеличение чувствительности достигается увеличением диаметра апертуры, и/или удлинением рецепторов, и/или увеличением диаметра рецепторов, и/или укорочением фокусного расстояния [28]. Чувствительность глаза к белому свету рассчитывается по формуле (5б, мкм^ - ср-*) [41]:
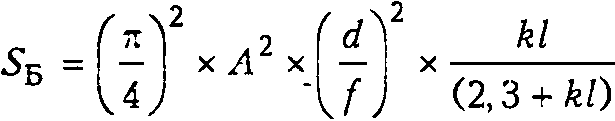
где Л — диаметр апертуры, d — диаметр фоторецептора, f — фокусное расстояние оптической системы, к — коэффициент экстинкции зрительного пигмента у ракообразных in situ, равный 0,0067 мкм-1 [16], I — длина фоторецептора, соответствующая длине мик-ровиллярного слоя.
Дополнительным параметром, характеризующим чувствительность глаза к свету, является F-число [28]:
f = L (3)
А где f — фокусное расстояние оптической системы и Л — диаметр апертуры. Чем ниже F-число, тем больше света попадает на сетчатку, тем более яркое изображение формируется на ней и тем выше чувствительность глаза [40]. Глаза с низким F-числом — те, у которых апертуры больше или почти такого же размера, что и фокусное расстояние глаз [38].
Для того чтобы выполнять определенную зрительную задачу, глаза должны быть способны к двум основным вещам: во-первых, собирать достаточное количество света, во-вторых, определять, из какого направления в пространстве падает свет. Другими словами, глаза должны иметь адекватную чувствительность к доступному свету и адекватную разрешающую способность [28; 39]. В настоящее время известно около 90 тыс. видов брю хоногих моллюсков [1], и только для 18 из них проведены соответствующие исследования глаз, необходимые для оценки их оптической чувствительности и разрешающей способности (табл. 1). Это морские переднежаберные моллюски Littorina irrorata Say, 1822 [26], Littorina littorea Linnaeus, 1758 [33], Strombus raninus Linnaeus, 1758 [34], пресноводный переднежаберный моллюск Ampularia sp. [35], пресноводные легочные моллюски Lymnaea stagnalis Linnaeus, 1758, Planorbarius corneas Linnaeus, 1758, Radix peregra Muller, 1774, Physa fontinalis Linnaeus, 1758 и наземные легочные моллюски Trichia hispida Linnaeus, 1758 из Калининградской области, Сераеа nemoralis Linnaeus, 1758 [22; 37], Arion rufus Linnaeus, 1758 [5], Cochlodina laminata Montagu, 1803, Perforatella incarnata Muller, 1774 [6], Helicigona lapicida Linnaeus, 1758 [7], Arianta arbustorum Linnaeus, 1758, Cepaea hortensis Muller, 1774 [8], Sue cine a putris Linnaeus, 1758 [11], T. hispida из Южной Швеции [12]. Как правило, если оптическая чувствительность низкая, разрешающая способность высокая, и наоборот [39]. Глаза перечисленных видов моллюсков представляют собой именно такой пример классического соотношения между оптической чувствительностью и разрешающей способностью (см. табл. 1). В глазах Л. rufus [9], С. laminata, Р. incarnata [6; 9], Н. lapicida [7; 9], A. arbustorum, С. hortensis [8 — 9], 5. putris [И], Т. hispida из Южной Швеции [12], L. stagnalis, R. peregra, Ph. fontinalis, T. hispida из Калининградской области и С. nemoralis могут быть отмечены следующие соотношения: высокая оптическая чувствительность фоторецепторов первого типа/низкая оптическая чувствительность фоторецепторов второго типа и низкая разрешающая способность фоторецепторов первого типа/высокая разрешающая способность фоторецепторов второго типа (за исключением разрешающей способности фоторецепторов второго типа у L. stagnalis, Т. hispida из Калининградской области, С. nemoralis и фоторецепторов второго типа второго вида у Н. lapicida) (см. табл. 1). Вероятно, более чувствительные фоторецепторы первого типа могут функционировать при более низком уровне освещенности, а менее чувствительные фоторецепторы второго типа — при менее высоком [19].
Табл. 1
Зрительные параметры камерных глаз брюхоногих моллюсков
|
Вид моллюска |
R, рад 1 |
Аф, град |
5В, мкм2 • ср 1 |
Г-число |
|
Морские переднежаберные |
||||
|
Littorina irrorata [26J |
31,8 |
1,04 |
0,3 |
1,75 |
|
Littorina littorea [33] |
18,28 |
1,8 |
0,37 |
1,2 |
|
5 trombus raninus (L.) [34] |
69,5 |
0,47 |
4,65 |
1,1 |
|
Пресноводные переднежаберные |
||||
|
Ampularia sp. [35] |
41,8 |
0,79 |
0,15 |
1,7 |
|
Пресноводные легочные |
||||
|
Lymnaea stagnalis [22] |
6,3/11 (л, г) 7,0/12 (д) 12/12 (в) |
5,2/3,0 (л, г) 4,7/2,7 (д) 2,7/2,7 (в) |
1,3/0,04 (л, г) 1,8/0,04 (д) 0,8/0,04 (в) |
2,9 |
|
Planorbarius corneus [22] |
13(д, в) |
2,5 (д, в) |
1,4 (д, в) |
1,8 |
|
Radix peregra [22] |
-/6,5 (л) -/7,0(г) -/11 (д) 16/16,5 (в) |
— /5,0 (л, г) -/3,0 (д) 2,0/2,0 (в) |
-/0,2 (л) -/0,1 (г) -/0,1 (д) 0,2/0,02 (в) |
1,7 |
|
Physa fontinalis [22] |
-/7,0 (д) -/8,0(в) |
-/5,0 (д) -/4,0 (в) |
- /0,2 (д, в) |
1,8 |
|
Наземные легочные |
||||
|
Trichia hispida (Калининградская область) [22] |
2,5/2,5 |
13/13 |
4,0/0,8 |
1,2 |
|
Cepaea nemoralis [22] |
4,0/4,0 |
8,0/8,0 |
8,0/1,5 |
1,3 |
|
Arion rufus [5] |
15/41 |
2,2/0,8 |
4,0/0,01 |
1,6 |
|
Cochlodina laminata [6] |
6,0/14 |
5,5/2,4 |
0,3/0,01 |
1,3 |
|
Perforatella incarnata [6] |
7,1/15 |
4,7/2,2 |
2,9/0,07 |
1,1 |
|
Helicigona lapicida [7] |
8,5/9,5/ 7,2/15,8 |
3,9/3,5/ 4,6/2,1 |
0,5/0,04/ 0,008/0,006 |
1,6 |
|
Arianta arhustorum [8] |
13,6/36 |
2.4/0,9 |
0,73/0,002 |
1,8 |
|
Cepaea hortensis [8] |
12,7/24 |
2,6/1,4 |
0,65/0,02 |
2,0 |
|
Succinea putris [11] |
4,9/12 |
6,8/2,7 |
1,2/- |
1,1 |
|
Trichia hispida (Южная Швеция) [12] |
4,0/- |
8,4/- |
1,8/- |
1,1 |
Примечание. R — разрешающая способность фоторецепторов; Аф — разрешаемое угловое расстояние фоторецепторов; 5Б — оптическая чувствительность фоторецепторов; одиночные цифры — средние значения для единственного первого типа фоторецепторов в центральной сетчатке; через дробь даны средние значения для фоторецепторов первого и второго типа в центральной сетчатке; для Н. lapicida через дробь даны средние значения для фоторецепторов первого типа, второго типа первого, второго и третьего вида в центральной сетчатке; л — латеральная область сетчатки, д — дорсальная ямка сетчатки, в — вентральная ямка сетчатки, г — гребень, разделяющий ямки.
Факторы, влияющие на разрешающую способность
Оптическая чувствительность. Первостепенным условием для реализации разрешающей способности глаза является адекватный захват фотонов, т. е. адекватная чувствительность к доступному свету. Это связано с тем, что яркость любого стимула оценивается по количеству фотонов, достигающих фоторецепторов. Если за длительный период времени фоторецепторы получают только несколько фотонов, им очень сложно точно определить яркость стимула и его локализацию в пространстве. Для того чтобы справляться с этими задачами, а также определять различия в яркости стимулов и потенциально различать больше пространственных деталей, фоторецепторам необходимо как можно большее количество фотонов. Количество фотонов, поглощенных фоторецепторами, определяется тремя основными факторами: уровнем освещенности окружающей среды, диаметром апертуры, диаметром и длиной фоторецепторов. При низких уровнях освещенности разрешающая способность достигается за счет высокой оптической чувствительности, которая в первую очередь обеспечивается большим диаметром апертуры, а также длинными и широкими рецепторами. В условиях свободного доступа фотонов их количество больше не является лимитирующим фактором разрешающей способности, поэтому необходимости в сохранении таких же размеров апертуры и рецепторов нет. При этих надпороговых уровнях интенсивности света острота зрения определяется уже возможностями оптической системы глаза и структурой сетчатки [28; 40]. Сравнительные исследования показали, что оптическая чувствительность фоторецепторов единственного первого типа у морских переднежаберных моллюсков L. irrorata [26], L. lit-torea [33], S. raninus [34], пресноводного переднежаберного моллюска Ampularia sp. [35], пресноводного легочного моллюска PL cor-neus, фоторецепторов первого и второго типа у пресноводных легочных моллюсков L. stag-nalis, R. peregra, Ph. fontinalis, наземных легочных моллюсков T. hispid а из Калининградской области, С. nemoralis [22], A. rufus [5], С. laminata, Р. incarnata [6], A. arbu-storum, С. hortensis [8], 5. putris [И], Т. hispida из Южной Швеции [12] и фоторецепторов первого типа, второго типа первого, второго и третьего вида у наземного легочного моллюска Н. lapicida [7] соответствует световым условиям их предпочитаемых мест обитания.
Положение фокуса. Для ясного видения фокус попадающих в глаз лучей от рассматриваемого объекта должен быть совмещен с сетчаткой (рис. 1, а?) [30]. Брюхоногие моллюски имеют глаза с фиксированной оптикой, т. е. ни положение, ни форма хрусталика у них не меняется [22]. У моллюсков с полусферической сетчаткой диоптрический аппарат глаза не всегда формирует отчетливое изображение на микровиллярном слое. Иногда на нем получается нечеткое изображение, как у пресноводного переднежаберного моллюска Ampularia sp. [35] и \ наземного легочного моллюска Т. hispida из | Калининградской области [22], или сильно I размытое изображение, как у наземных легочных моллюсков A. arbusiorum и С. hortensis [8]. У морских переднежаберных мол- | люсков L. irrorata [26], L. littorea [33], S. га- ( ninus [34] и наземных легочных моллюсков С. nemoralis [22], A. rufus [5], С. laminata, Р. incarnata [6], Н. lapicida [7], 5. putris [И], Т. hispida из Южной Швеции [12] оптическая система глаза создает отчетливое изображение на микровиллярном слое полусферической сетчатки. У пресноводных легочных моллюсков L. stagnalis, Pl. corneus R. pereg-ra, Ph. fontinalis в сетчатке сформировались дорсальная и вентральная ямки разной глубины, которые дают моллюскам возможность видеть и в водной, и в воздушной среде. У данных видов положение глаза в периопти-ческом синусе в воде не влияет на величину фокусного расстояния оптической системы. В воздухе фокусное расстояние оптической системы глаза, который максимально втянут в глубину периоптического синуса, длиннее фокусного расстояния оптической системы глаза, который соприкасается с эпидермисом [22; 37].
Сферическая аберрация возникает из-за того, что периферические лучи света фокусируются ближе к задней поверхности хрусталика, чем параксиальные (приосевые) лучи. Различие в положении фокуса для разных лучей делает изображение размытым (рис. 1, б). Положение наилучшего фокуса совпадает с наименьшим латеральным распространением лучей. Сферическая аберрация может быть откорректирована двумя способами: созданием линзы с асферической (параболической) поверхностью и созданием градиента показателя преломления вещества хрусталика от центра (высокий) к периферии (низкий). В отсутствие корректирующих механизмов сферическая аберрация сильнее выражена в глазах с низким F-чис-лом [28; 40].

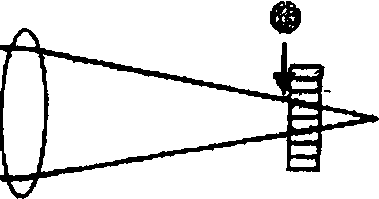
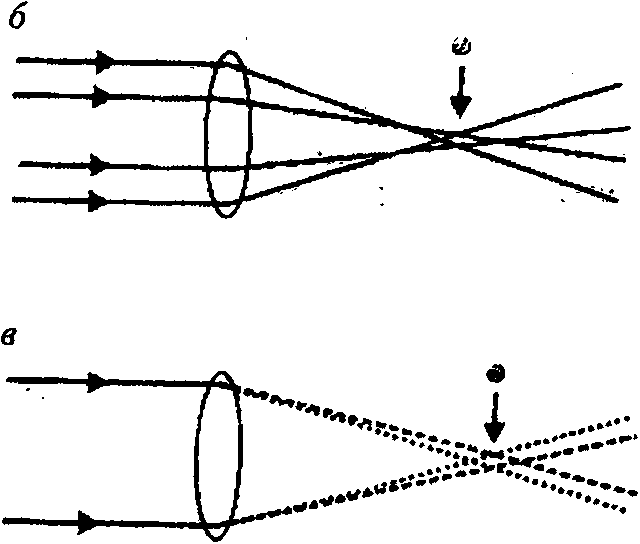
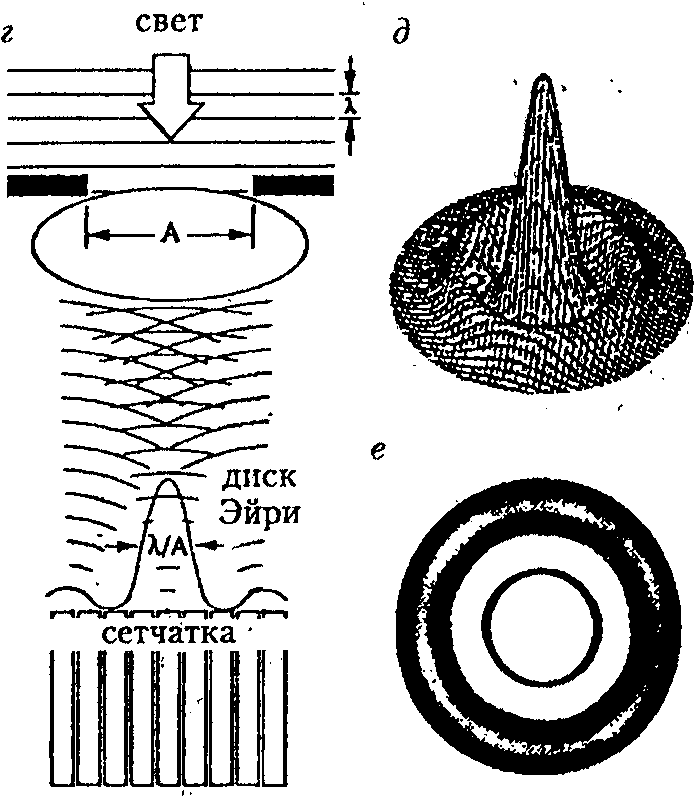
Рис. 1 ■ ,
Оптические дефекты: а — дефект фокуса; б — сферическая аберрация; в — хроматическая аберрация [30] (стрелка показывает положение размытого изображения); г — дифракция; д — трехмерное графическое изображение дифракционной картины Эйри;
е — дифракционная картина Эйри (вид сверху) [40] 1
У исследованных видов брюхоногих моллюсков поверхность хрусталиков и линз, образованных эпителием и роговицей, имеет полусферическую форму и, следовательно, не участвует в коррекции сферической аберрации. Качество изображений тестового объекта, сформированных изолированными хрусталиками, и/или рассчитанное значение отношения Меттиссена (f/r) для сферических хрусталиков позволяют предположить наличие или отсутствие в них градиента показателя преломления и, следовательно, сферической аберрации. Хрусталики морских переднежаберных моллюсков L. irrorata Q/r = = 2,7) [26], L. littorea (2,3) [33], 5. raninus (2,0) [34], пресноводных легочных моллюсков L. stagnalis (2,9), PL corneus, R. peregra (2,7), Ph. fontinalis (2,9) и наземных легочных моллюсков Т. hispida из Калининградской области (3,0), С. nemoralis (2,1) [22], A. rufus [5; 9], С. laminata (2,7) [6], Н. 1арг-cida [7], A. arbustorum, С. hortensis [8], 5. putris [И] создают отчетливые изображения и/или имеют отношение Меттиссена, близкое к идеальному (//г ® 2,55), что указывает на негомогенность хрусталиков и коррекцию сферической аберрации. Хрусталики пресноводного переднежаберного моллюска Ampularia sp. [35] и наземных легочных моллюсков Р. incarnata [6] и Т. hispida из Южной Швеции [12] формируют нечеткие изображения, что свидетельствует о гомогенности хрусталиков и присутствии сферической аберрации, но у Ampularia sp. с более высоким Г-числом глаза (1,7) она выражена в меньшей степени, чем у Р. incarnata и Т. hispida из Южной Швеции с более низким F-числом глаз (1,1).
Хроматическая аберрация ъожкка.-ет из-за того, что диоптрический аппарат глаза преломляет коротковолновой свет (ультрафиолетовый) сильнее, чем длинноволновой свет (красный)^(рис. 1, в). Изображение становится размытым, а положение наилучшего фокуса то, при котором латеральное распространение лучей наименьшее. Одним из способов коррекции хроматической аберрации является замена одного хрусталика двумя или более, каждый из которых должен иметь разные показатели преломления и индивидуальную форму. Точно подобранная комбинация доставляет лучи волн разной длины к общему фокусу, но таких примеров в природе пока нет. Другой возможный путь коррекции — использование дисперсии за счет размещения фоторецепторов определенной спектральной чувствительности в положении, соответствующем фокальной плоскости определенной длины волн: фоторецепторы с максимальной чувствительностью в коротковолновой области лежат в дистальных слоях сетчатки, а фоторецепторы с максимальной чув- ствитслыюстыо в длинноволновой области — в проксимальных. Среди беспозвоночных животных такое строение имеет только сетчатка глаз прыгающих пауков. Хроматическая аберрация влияет на разрешающую способность, когда диаметр апертуры (А) превышает 500 мкм [28; 40].
Абсолютный диаметр апертур у морских переднежаберных моллюсков L. irrorata (А = 132 мкм) [26], L. littorea (108 мкм) [33], пресноводного переднежаберного моллюска Ampularia sp. (260 мкм) [35], пресноводных легочных моллюсков L. stagnalis (60 мкм), PL corneus (140 мкм), R. регедга (73 мкм), Ph. fontinalis (65 мкм) и наземных легочных моллюсков Т. hispida из Калининградской области (92 мкм), С. nemoralis (107 мкм) [37], A. rufus (223 мкм) [5], С. laminata (70 мкм), Р. incarnata (145 мкм) [6], Н. lapicida (103 мкм) [7], A. arbustorum (125 мкм), С. hortensis (120 мкм) [8], S. putris (105 мкм) [11], Т. hispida из Южной Швеции (90 мкм) [12] указывает на то, что хроматическая аберрация нс влияет на их зрение. Диаметр апертуры морского переднежаберного моллюска 5. raninus (850 мкм) превышает 500 мкм [34], по и в этом случае хроматическая аберрация вряд ли будет ограничивать разрешающую способность глаза, поскольку все исследованные к настоящему времени брюхоногие моллюски являются монохроматами [2 — 3; 10; 14; 18—19; 24-25; 27].
Дифракция. Изображение точечного источника на сетчатке будет размытым, даже если сферическая и хроматическая аберрации выражены незначительно. Этот факт является следствием волновой природы света. Свет от точечного источника достигает хрусталика в виде плоского волнового фронта. Проходя через хрусталик, центральная область волнового фронта задерживается по сравнению с краевыми областями, потому что проходит через большую часть оптически плотного материала. В результате выходящий волновой фронт приобретает полусферическую форму, центрированную па фокусе и расходящуюся в стороны от него. В фокусе разные участки волнового фронта встречаются и, поскольку проходят друг через друга, интерферируют. Компоненты, совпадающие по фазе, усиливают друг друга, а несовпадающие — гасят. Таким образом, изображение точечного источника формируется в виде яркого пика, окруженного светлыми (возникающими от усиления) и темными (возникающими от гашения) кольцами. Изображение называется дифракционной картиной Эйри, а ее центральный пик — диском Эйри (рис. 1, г — е) [28; 30]. Глаза с большими апертурами имеют маленькие диски Эйри и в меньшей степени подвергаются дифракции. Диаметр диска Эйри (D3, мкм), образуемого на сетчатке точечным источником света, определяется по формуле [28]:
D3 = 2,44 х f х
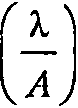
где / — фокусное расстояние оптической системы, А — диаметр апертуры и X = 500 нм — значение, близкое к максимумам чувствительности зрительных пигментов фоторецепторов у большинства исследованных видов брюхоногих моллюсков [2 — 3; 10; 14; 18 — 19; 24 — 25; 27].
У исследованных видов брюхоногих моллюсков диаметр диска Эйри в несколько раз меньше самых маленьких расстояний между центрами соседних фоторецепторов. Поэтому дифракция не ограничивает разрешающую способность глаз морских переднежаберных моллюсков L. irrorata (2,1 мкм < <4,1 мкм) [26], L. littorea (1,4 мкм < 4,0 мкм) [33], S. raninus (1,4 мкм < 7,8 мкм) [34], пресноводного переднежаберного моллюска Ampularia sp. (2,0 мкм < 5,9 мкм) [35], пресноводных легочных моллюсков L. stagnalis (3,1 мкм < 8,3 мкм), PL corneus (1,9 мкм < < 6,2 мкм), R. регедга (1,7 мкм < 4,3 мкм), Ph. fontinalis (1,9 мкм < 6,8 мкм) и наземных легочных моллюсков Т. hispida из Калининградской области (1,5 мкм <15 мкм), С. nemoralis (1,6 мкм <21 мкм) [22; 37], A. rufus (1,9 мкм <5,0 мкм) [5], С. laminata (1,6 мкм <4,0 мкм), Р. incarnata (1,3 мкм < < 6,0 мкм) [6], Н. lapicida (2,0 мкм < 6,0 мкм) [7], A. arbustorum (2,2 мкм < 3,4 мкм), С. hortensis (2,5 мкм < 6,0 мкм) [8], 5. putris (1,4 мкм <5,7 мкм) [И], Т. hispida из Южной Швеции (1,3 мкм < 5,3 мкм) [12].
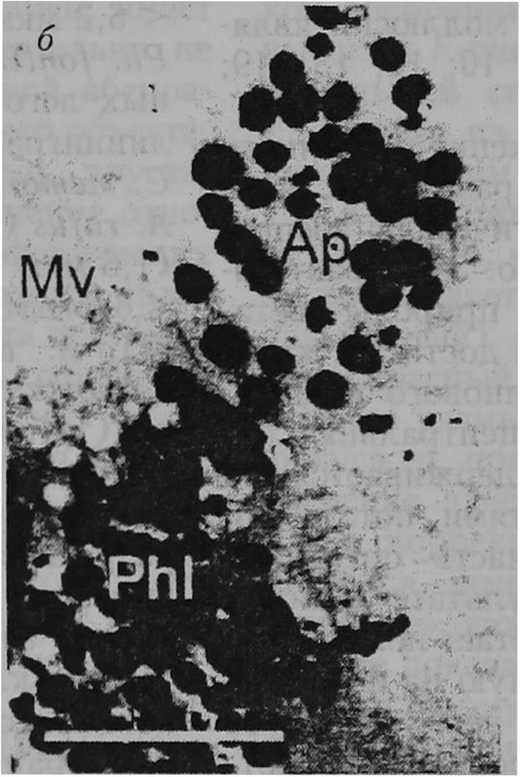
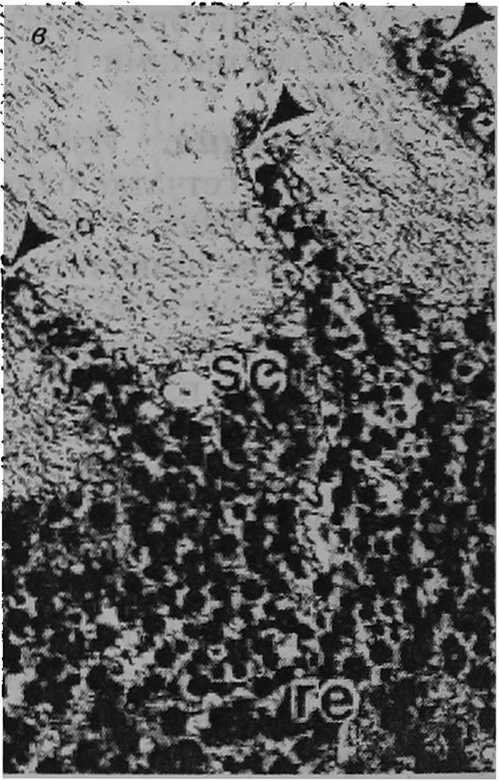
Структура сетчатки. Снижение разрешающей способности глаза может быть вызвано латеральным рассеиванием света между микровиллами соседних фоторецепторных клеток. Для ослабления эффекта такого рассеивания служит механизм миграции гранул экранирующего пигмента [28; 40] в апикальные отростки фоторецепторов. Светозависимая миграция пигментных гранул наблюдается, например, в фоторецепторах первого типа у пресноводных легочных моллюсков L. stagnalis, Pl. corneus, R. регедга, Ph. fontinalis. Отмечено, что относительная плотность пигментных гранул в пальцевидных дистальных частях фоторецепторов первого типа меняется от очень низкой при темновой адаптации (несколько гра- пул) до очень высокой при световой адаптации (множество гранул) (рис. 2, а — б) [37]. У морского переднежаберного моллюска Aporrhais pespelecani Linnaeus, 1758, так же как и у многих других исследованных видов брюхоногих моллюсков, фоторецепторные клетки не имеют апикальных отростков. Между тем только у A. pespelecani тела пигментных клеток длиннее тел фоторецепторных клеток [15]. Тем самым пигментные клетки оптически изолируют микровиллы соседних рецепторных клеток друг от друга почти по всей их длине и устраняют рассеивание света (рис. 2, в). Очевидно, что глаза брюхоногих моллюсков имеют два экранирующих механизма, направленных на повышение остроты зрения. Первый — светозависимая миграция пигментных гранул в апикальные отростки фоторецепторов — позволяет уменьшить распространение света между микровиллами соседних клеток; второй — оптическая изоляция микровилл фоторецепторов — предотвратить такое распространение. У моллюсков, в сетчатке которых нет экранирующих механизмов, рассеивание света на микровиллах соседних рецепторных клеток становится лимитирующим фактором разрешающей способности глаз. Эффект та кого рассеивания прямо пропорционален длине (0 микровиллярного слоя фоторецепторов. В связи с этим латеральное рассеивание выражено в большей степени у пресноводного легочного моллюска L. stagnalis U = 71,0 мкм) [37], морского переднежаберного моллюска 5. raninus (60,0 мкм) [34], пресноводного легочного моллюска Ph. fon-tinalis (49,1 мкм) [37], наземного легочного моллюска A. rufus (43,0 мкм) [5] и в меньшей степени у наземных легочных моллюсков A. arbustorum (24,0 мкм), С. hortensis (23,0 мкм) [8], морского переднежаберного моллюска L. littorea (20,0 мкм) [33], наземных легочных моллюсков С. nemoralis (20,0 мкм) [37], Р. incarnata (20,0 мкм) [6], пресноводных легочных моллюсков PL сог-neus (18,5 мкм), R. регедга (18,0 мкм), наземных легочных моллюсков Т. hispida из Калининградской области (15,0 мкм) [37], 5. putris (14,0 мкм) [И], Т. hispida из Южной Швеции (12,0 мкм) [12], Н. lapicida (11,0 мкм) [7], морского переднежаберного моллюска L. irrorata (10,0 мкм) [26], наземного легочного моллюска С. laminata (9,0 мкм) [6], пресноводного переднежаберного моллюска Ampularia sp. (7,0 мкм) [35].
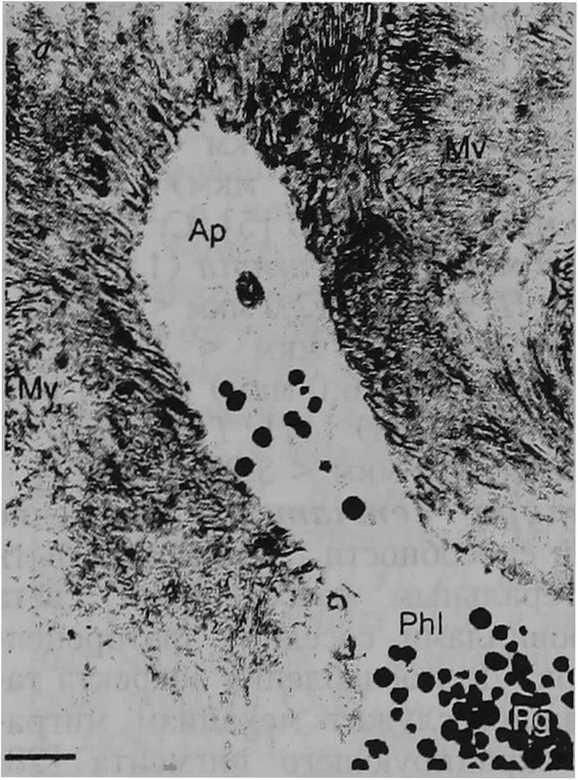
Рис. 2
Экранирующие механизмы в глазах брюхоногих моллюсков: а и 6 — гранулы экранирующего пигмента в апикальных отростках фоторецепторов первого типа пресноводного легочного моллюска Lymnaea stagnalis в условиях темновой и световой адаптации соответственно [37] (масштаб — 1 мкм): Ph I — фоторецептор; Ар — апикальный отросток фоторецептора; Mv — микровиллы; Pg — пигментные гранулы; в — участок сетчатки глаза морского переднежаберного моллюска Aporrhais pespelecani [15] (масштаб — 5 мкм): ге — сетчатка, sc — фоторецепторные клетки, стрелки показывают тела пигментных клеток между микровиллами соседних фоторецепторных клеток
Заключение. Из представленного материала видно, что возможности камерных глаз исследованных видов брюхоногих моллюсков неодинаковы. Оптическая чувствительность глаз моллюсков адекватна световым условиям их предпочитаемых мест обитания. Оптическая система глаз как таковая помещает изображение разного качества на микровиллярпом слое сетчатки. При нечетком или сильно размытом изображении разрешающая способность фоторецепторов снижается или не достигается. При отчетливом изображении разрешающая способность фоторецепторов в большей или меньшей степени снижается вследствие сферической аберрации и/или латерального рассеивания света между микровиллами соседних рецепторных клеток. Хроматическая аберрация и дифракция не влияют на зрение моллюсков. Для идентификации окружающих рбъектов, разрешаемое угловое расстояние фоторецепторов глаза не должно превышать несколько градусов [29], а оптическая чувствительность глаза должна соответствовать световым условиям мест обитания [40]. Таким образом, согласно результатам морфологических и оптических исследований камерные глаза могут функционировать как органы зрения у морских переднежаберных моллюсков L. irrorata [26], L. littorea [33], 5. raninus
[34], пресноводных легочных моллюсков L. stagnalis, PL corneus, R. peregra, Ph. fon-tinaliSj наземных легочных моллюсков C. nemoralis [22], A. rufus [5; 9], С. laminata, Р. inearnata [6], Н. lapicida [Ffr S. putris [11], T. hispida из Южной Швеции [12] или только как детекторы освещенности у пресноводного переднежаберного моллюска Ampularia sp. [35], наземных легочных моллюсков Т. hispida из Калининградской области [22], A. arbustorum, С. hortensis [8].
Исходя из данных о функциональных возможностях камерных глаз 18 видов брюхоногих моллюсков, можно выделить факторы, "ограничивающие их разрешающую способность. Во-первых, это латеральное рассеивание света между микровиллами соседних фоторецепторных клеток, поскольку оно присутствует у всех видов моллюсков, хотя и в разной степени. Во-вторых, положение фокуса, которое у четырех видов таково, что на микровиллярпом слое сетчатки формируется нечеткое или сильно размытое изображение. В-третьих, сферическая аберрация, которая наблюдается у трех видов из-за отсутствия корректирующих механизмов (табл. 2). Таким образом, из шести возможных факторов три ограничивают разрешающую способность камерных глаз исследованных видов брюхоногих моллюсков.
Табл. 2
Факторы, влияющие и не влияющие на разрешающую способность камерных глаз брюхоногих моллюсков
|
, Вид моллюска |
Адекватная оптическая чувствительность |
Положение фокуса на сетчатке |
Сферическая аберрация |
Хроматическая аберрация |
Дифракция |
Латеральное рассеивание света |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Littorina irrorata [26] |
+ |
+ |
— |
— |
*— |
4* |
|
Littorina littorea [33] |
+ |
+ |
— |
— |
— |
4- |
|
Strombus raninus [34] |
+ |
4- |
— |
— |
— |
4- |
|
Ampularia sp. [35] |
+ |
— |
+ |
— |
— |
4- |
|
Lymnaea stagnalis [22] |
+ |
+ |
— |
— |
4- |
|
|
Planorbarius corneus [22] |
+ |
+ |
— |
— |
— |
4- |
|
Radix peregra [22] |
+ |
+ |
— |
— |
4- |
|
|
Physa fontinalis [22] |
4- |
+ |
— |
— |
— |
4- |
|
Trichia hispida (Калининградская область) [22] |
4- |
+ |
— |
— |
— |
4- |
|
Cepaea nemoralis [22] |
4- |
+ |
— |
— |
— |
4- |
Окончание табл. 2
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Arion rufus [5] |
+ |
+ |
* |
- |
- |
* |
|
Cochlodina laminata [6] |
+ |
+ |
- |
- |
- |
+ |
|
Perforatella incarnata [6] |
+ |
+ |
+ |
- |
- |
+ |
|
Helicigona lapicida [7] |
+ |
+ |
- |
- |
- |
+ |
|
Arianta arbustorum [8] |
+ |
- |
- |
- |
- |
+ |
|
Cepaea hortensis [8] |
+ |
- |
- |
- |
- |
+ |
|
Succinea putris [11] |
+ |
+ |
- |
- |
- |
+ |
|
Trichia hispida (Южная Швеция) [12] |
+ |
+ |
+ |
- |
- |
+ |
Примечание. «+» — присутствие фактора, *-> — отсутствие фактора.
