Развитие Lathurys japonicus Willd. (сем. Fabaceae) в первый год жизни в условиях культуры
 в первый год жизни в условиях культуры")
Автор: Калинкина Валентина Андреевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 4, 2016 года.
Бесплатный доступ
В статье рассмотрены биологические особенности реликтового вида сем. Бобовые - чины японской ( Lathyrus japonicus ). Описана динамика прорастания семян в условиях культы, определена энергия и всхожесть семян. Охарактеризованы этапы развития особей: тип прорастания, развитие надземной и подземной сфер особи в виргинильном периоде.
Чина японская, бобовые, онтогенез, семена, проросток, развитие
Короткий адрес: https://sciup.org/146116669
IDR: 146116669 | УДК: 581.4:582.73:633.378
The development of Lathyrus japonicus Willd. (Fabaceae) in the first year of life in the cultivated condition
We describe some of the biological peculiarities of the relict species of Fabaceae - Lathyrus japonicus. We pay special attention to the dynamics of germination of cultivated plants, as well as to the energy of germination and its success. The description of developmental stages from the germination to the appearance of the above- and underground parts at virginal period is given.
Текст научной статьи Развитие Lathurys japonicus Willd. (сем. Fabaceae) в первый год жизни в условиях культуры
Введение. Чина японская (L japonicus) - реликтовый амфиокеанический представитель семейства Legumosaea (Федченко, 1948), широко распространенный на побережьях морей и океанов с умеренным и умеренно холодным климатом. Встречается на тихоокеанском побережье Азии (от Чукотки до п-ова Корея и Японии), в Северной Америке (от Аляски до штата Орегон), в Приатлантической Европе (от арктической Скандинавии до Испании), на атлантическом побережье Северной Америки (от о. Ньюфаундлен до мыса Хаттерас) (Арктическая флора..., 1986).
Вид является хозяйственно-ценным растением, содержащим в траве кумарины и гамма-метилглутаминовую кислоту, в семенах -5-оксипипеколиновую кислоту. Надземные части растений (стебли, листья и семена) в Китае используют в пищу, отвары употребляют при сердечнососудистых заболеваниях, а так же в качестве мочегонного средства; водный настой травы полезен при эпилепсии; семена, как наркотическое средство, используют при психических нарушениях (Dragendorff, 1898; Шретер, 1975).
Чина японская, имея обширный ареал, характеризуется высокой степенью морфологической изменчивости, которая проявляется в изменении формы, размеров листочков и прилистников, длине и степени разветвления усика, числе цветков в соцветии, длине побега и степени опушения. Особенности морфологии послужили основанием к выделению разновидностей, подвидов, географических рас и новых видов (Федченко, 1948; Чефранова, 1971; Allkin et al., 1983; Арктическая флора..., 1986; Павлова, 1989; Рыжова и др., 2007; Вишнякова и др., 2008; The Plant List; Lathyrus japonicas; Sea Pea). He смотря на обширный ареал и хозяйственное значение в литературе (Туленкова, 1968а, б, 1970, 1974а, 1977; Петухова и др., 2003; Sadao, 2005 и др.) на данный момент отсутствуют сведения о начальных этапах развития чины японской.
В рамках изучения структурных особенностей представителей бобовых в условиях востока Азии изучена динамика прорастания семян и начальные этапы развития чины приморской в первый год жизни в условиях культуры. Исследования проводили на базе ФГБУН Ботнический сад-институт ДВО РАН.
Материал и методика. Материал для исследований был собран 1 августа 2014 г. в Приморском крае, Тернейском р-не, в районе урочища Благодатное, на восточном берегу оз. Благодатное, на прибрежных галечниковых участках (GPS координаты 44° 93'49,4118" с.ш; 136°54'99,9184"в.д., высота=31 м. н.у.м.).
Собранные семена высушивались и хранились в бумажных конвертах. Эксперимент по проращиванию семян был поставлен 26 января 2015 г. Перед посадкой, для разрушения твердых оболочек семенной кожуры в течение 30 мин. проводили обработку концентрированным раствором серной кислоты. После семена промывали в проточной воде и высеивали в чашки Петри на влажную фильтровальную бумагу. Пробы по 50 шт. семян взяты в трехкратной повторности. Проращивание проводили в климокамерах при следующих условиях: Т = 22,0°С, влажность = 80,5%, концентрация углекислого газа = 80,5%, продолжительность фотопериода= 16,5 ч.
Семя считали проросшим при формировании корешка размером с семя. Подсчет проросших семян проводили от начала прорастания ежедневно. Процент семян, проросших за десять дней со дня закладки опыта, рассматривали как энергию прорастания, а на пятнадцатый день определяли общую всхожесть семян.
Морфометрические параметры семян, этапы прорастания, начальные периоды развития растений описывали и измеряли при помощи стереомикроскопа Stemi-2000C и Stemi DV4 производства фирмы Carl Zeiss (Германия).
После прорастания особи пересаживались в грунт.
В связи с небольшой повторностью опыта мы считаем, что целесообразно указывать диапазон размерных характеристик побеговой и корневой системы растений. График динамики прорастания семян построен в программе Microsoft Office Excel.
В работе применены биоморфологический и онтогенетический методы. Онтогенетические (возрастные) состояния выделяли согласно методике Т.А. Работнова (1950).
Результаты и обсуждение. Латентный период. Плод боб. Семена округлые, шаровидные, некоторые неправильно-шаровидные, от коричневых до темно коричневых и темно-бурых, шероховатые. Длина семени 4,5+0,04 мм, ширина 4,3+0,04 мм. Масса 1000 семян 35,58+0,28 г. (Воронкова и др., 2003). Анатомические и ультраструктурные исследования семенной кожуры некоторых видов рода Lathyrus (Баранов, Бурляева, 1990; Ворончихин, 1997) показали, что поверхность семян L. japonicus представлена слабозвездчатыми бугорками различных размеров, покрытых эпикутикулой. Эпидерма представлена длинными узкими клетками, вытянутыми в радиальном направлении, гиподерма на поперечном срезе представлена слабо утолщенными стенками, вытянутыми в радиальном направлении. Индекс эпидерма / гиподерма составляет 3/1. Эпидерма и гиподерма на начальных стадиях своего развития представлены живыми тонкостенными клетками (Баранов, Бурляева, 1990). По мере созревания семени, клеточные стенки эпидермы и гиподермы утолщаются, протопласт в них разрушается, клетки эпидермы преобразуются в столбчатые макросклереиды, а клетки, расположенные под ней - в призматические склереиды. Образующаяся плотная водонепроницаемая кожура защищает семена при неблагоприятных условиях. Твердосемянность является особым типом физического или экзогенного покоя, свойственная многим представителям сем. Fabaceae (Николаева и др., 1985). Согласно литературным данным (Нестерова и др., 1997) семена чины японской при хранении в лабораторных условиях в бумажных пакетах сохраняют всхожесть в течении как минимум 12 лет. Для выведения семян из состояния покоя в лабораторных условиях используют метод скарификации.
Виргинильный период - характеризуется началом развития особи и объединяет возрастные состояния проросток, ювенильное, имматурное и молодое вегетативное.
Проросток. Чина приморская относиться к растениям с подземным типом прорастания семян, что обусловлено недоразвитием гипокотиля. Прорастание семян дружное, начинается на 6 день после скарификации, и длиться в течение 5 дней (рис. 1). Энергия прорастания и общая всхожесть семян в опыте составили 33,3%, что согласуется имеющимися в литературе сведения. Так, по данным Н.М. Воронковой и др. (2003), при скарификации семян L. japonicus раствором серной кислоты в течение 1 часа процент проросших семян составил 37%, 10 % оставались твердыми, остальные были не жизнеспособными и сгнивали. Вероятно, это связано с тем, что в стручках чины приморской только треть семян обладает хорошей жизнеспособностью, 2/3 - при хороших внешних показателях оказываются нежизнеспособными (Gurusamy et al., 2000). Прорастание последнего семени в нашем опыте отмечено на 49 день, что возможно обусловлено недостаточной скарификацией.
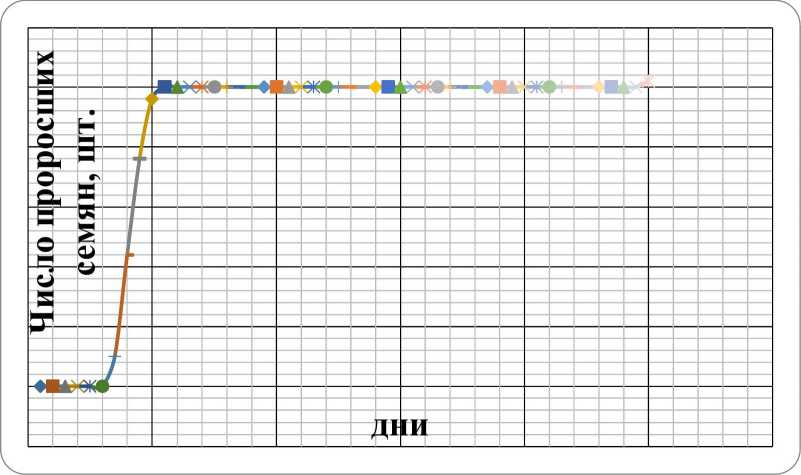
Рис. 1. Динамика прорастания семян L. japonicas в условиях культуры.
Прорастания семян начинается с роста зародышевого корешка, который дает начало главному корню. Далее в рост трогается конус нарастания побега; на 3-5 день развивается первый низовой лист, он, как и 2-3 последующих - чешуевидный, недоразвитый (рис. 2). Длина эпикотиля составляет 2,1-3,5 мм. Первые два чешуевидных листа сближены, длина междоузлия 7-8 мм; третий-четвертый (при их развитии) - обычно удалены от предыдущих, длина междоузлия достигает 3,5-4 см.
Первый настоящий лист развивается на 6-7 день. Он сложный, парноперистый, с одной парой листочков, черешковый с прилистниками; рахис заканчивается простым усиком. Листочки эллиптической или обратнояйцевидной формы. Редко наблюдается редукция доли сложного листа, или отсутствие усика. Длина листочка 8-10 (12) мм, ширина - 4-7 (9) мм; длина усика - 1 мм; длина черешка - 2-3 мм. Прилистники ни формой, ни размером не имеют ничего общего с взрослыми растениями. У проростков они ланцетной или шиловидной формы, длиной 2-3 мм. Тип побега - прямостоячий. Общая длина особи составляет 4-6 см.

Рис. 2. Этапы развития L. japonicus в первый год жизни в культуре.



Главный корень у проростков нарастает до 2-3,5 см. Ветвление его слабое. По мере роста растения несколько удлиняется гипокотиль, отличающийся от главного корня по цвету и толщине. В этом возрастном состоянии связь растения с семенем сохраняется, особи переходят к смешанному питанию (гетеротрофному и автотрофному), рост их идет за счет использования запасных веществ семядолей, ассимилятов листьев и корневого питания.
Маркером перехода особи в ювенильное возрастное состояние в надземной сфере является развитие второго настоящего листа, форма листочков, которого яйцевидно-округлая. Листья ювенильных растений сложные, парноперистые; нижние два-три листа имеют одну пару листьев (иногда наблюдается недоразвитие одного листочка второй пары листьев); рахис заканчивается простым усиком длиной 12 мм. Следующие один-два листа развивают две (редко одну) пары листьев, длина усика достигает 2,6 см. Длина листочков 9-13 мм, ширина 9-11 мм, длина черешком 7-8 мм. Прилистники ланцетные, длиной 3-4 мм. Длина побега достигает 10-15 см. Тип побега -прямостоячий или полегающий. Связь с семенем сохраняется.
Соответственно изменению надземных органов, усложняется структура и размер подземных. Длина главного корня достигает 3,23,9 см, ширина 0,5-1,3 мм. На гипокотиле наблюдается развитие придаточных корней, длина которых может достигать 1-2 см (рис. 3).
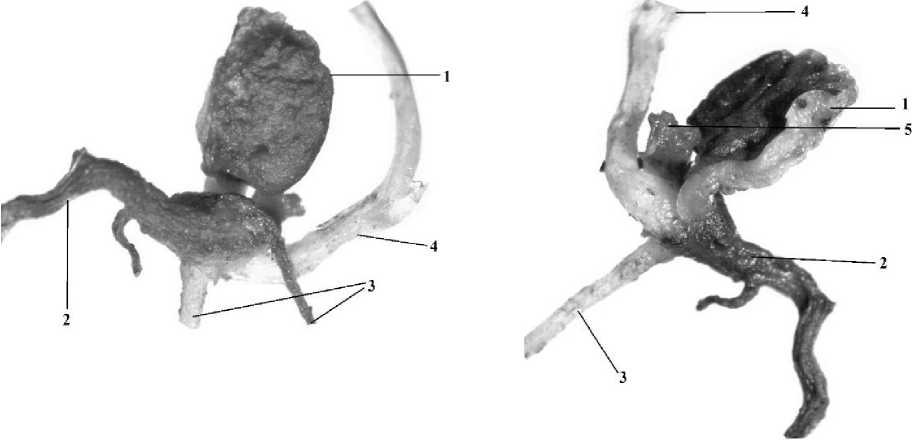
Рис. 3 . Процесс отмирание главного корня и семядолей у L. japonicas: X - вид спереди; Б - вид сзади; 1 - семядольный лист, 2 - главный корень, 3 - придаточный корень, 4 - главный побег, 5 - черешок семядольного листа.
У чиня японской, как и у многих других представителей сем. Бобовые (Туленкова, 1968а, 19746; Калинкина, 2008), возможно образование нескольких сериально или коллатерально расположенных почек. В ювенильном возрастном состоянии у особей L. japonicus в пазухе первого-третьего чешуевидных листьев закладываются одиночные, а в основании эпикотиля внутри семядольных листьев -сериальные почки (рис. 4).
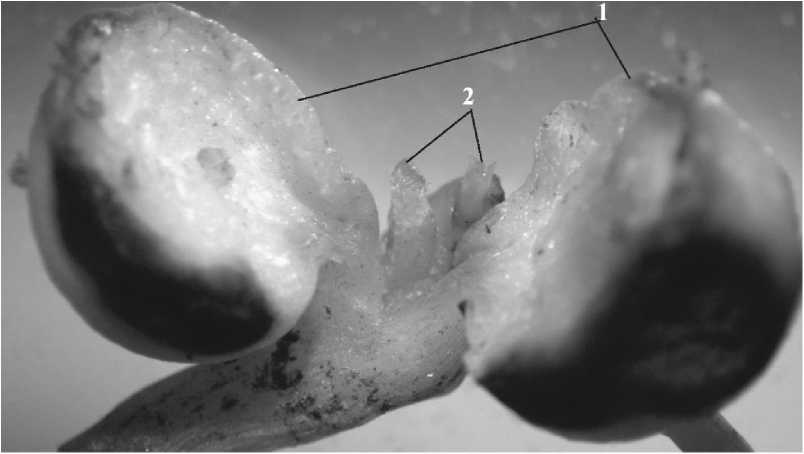
Рис. 4 . Сериальные почки L. japonicus.
1 - семядольные листья; 2 - сериальные почки.
Длительность ювенильного возрастного состояния 1,5 месяцев.
В возрасте около 2 мес. особи L. japonicus переходят в имматурное возрастное состояние, которое в надземной сфере характеризуется формированием боковых побегов, а в подземной -интенсивным развитием корневой системы (рис. 2). Формирующиеся в этот период листья парноперистые, имеют две (редко одну) пары листьев, рахис заканчивается простым усиком, длиной до 3 см. Размер листочков увеличивается, по сравнению с ювенильным возрастным состоянием на 1-2 мм. Длина усика уже позволяет особи при соприкосновении с опорой, по возможности фиксировать годичный побег, однако побеги не обвивают опору, подобно травянистым лианам. Тип побега - стелющийся. Кроме развития более сложных листьев в основании гипокотиля наблюдается закладка почек возобновления.
На этом этапе связь с семенем минимальна. У некоторых особей происходит полный переход с автотрофного и смешанного питания на гетеротрофный.
Главный и придаточный корни начинают интенсивно ветвиться, образовывая боковые корни второго порядка. Длина главного корня достигает 5-8 см, ширина 0,8-1 мм. Число придаточных корней варьирует от 1 до 5, их длина может достигать 5-8 см.
В начале лета наблюдается переход особей в вегетативное возрастное состояние , которое характеризуется не только увеличением общей массы надземной и подземной сферы, но и качественными морфологическими изменениями обоих сфер (рис. 2).
Главный побег интенсивно ветвиться, однако отмечено, что боковые побеги 1-го порядка берут на себя лидирующую роль, более интенсивно развиваются и ветвятся по сравнению с материнским. Длина главного побега достигает 16-18 см, длина боковых - 90 см. Тип побега — стелющийся. Число долей развивающихся в этот время парноперистых листьев увеличивается до 3 (однако могут развиваться листья с 2 парами листочков); рахис так же простой, длиной до 3 см. Листочки овальные, длиной до 1,7 см, шириной - 1,1 см. Форма прилистников листовидная или копьевидная, близка к форме взрослых растений. Длина прилистников 9 мм, ширина в основании - 5 мм.
Благодаря контрактильной деятельности главного корня происходит втягивания базальной части главного побега глубже в почву. Заложенные на гипокотиле и эпикотиле почки возобновления трогаются в рост, подземные побеги около месяца развиваются плагеотропно. Длина их за этот период достигает 15-25 см. Междоузлия удлиненные, число узлов 8-10. Листья недоразвитые, чешуевидные. В конце августа в основании некоторых почек возобновления, закладывающихся в пазухах этих листьев, развиваются тонкие придаточные корни. При выходе на поверхность, верхушечная почка плагеотропного побега формирует надземный ассимилирующий побег, морфометрические параметры которого соизмеримы с таковыми у главного побега.
Главный и придаточные корни интенсивно растут и ветвятся до 2-3-го порядка. Длина главного корня достигает - 20 см, ширина в основании - 2,5 мм. Боковые корни проникают на глубину 25 см, придаточные - 10-12 см.
Развитие азотфиксирующих клубеньков, характерных для взрослых особей (Gurasamy, Davis, Bal, 2000) в вегетативном возрастном состоянии нами не отмечена.
В конце сентября особи надземная вегетативная масса L. japonicus отмирает. Сохраняется базальная часть побегов с нереализованными в первый год развития почками возобновления, которая в дальнейшем войдет в состав каудекса, а так же подземные плагеотропные побеги. В зависимости от степени ветвления главного побега, образуется одно- или двух-трехглавый каудекс. Благодаря способности главного корня сокращаться и вкручиваться в почву, почки возобновления располагаются на глубине 0,2-0,3 см под землей, что защищает их от неблагоприятных условий среды.
Заключение. Морфологический анализ особей чины японской, развивающихся в течение первого года жизни в условиях культуры, позволил выделить два периода и четыре возрастных состояния. В каждом из них растение характеризуется определенной морфологической структурой надземных и подземных органов. У особей L. japonicus уже в виргинильном возрастном периоде наблюдается смена жизненной формы: стержнекорневая жизненная форма, характерная для проростков, в конце вегетационного сезона модифицируется в стержне-кистекорневую с подземными плагеотропными побегами, которые на второй год жизни особи станут одним из звеньев гипогеогенного корневища.
Надземный побег так же претерпевает ряд морфологических изменений. В течение вегетационного сезона наблюдается усложнение листьев - от простых чешуевидных до сложных, парноперистых с простым усиком. Прямостоячий тип побега сменяется стелющимся, характерным для взрослых особей этого вида.
Развитие большого числа почек возобновления позволит особи на следующий год дать массу годичных надземных и многолетних подземных побегов, обеспечив формирование у взрослых растений явно-полицентрической жизненной формы (Смирнова и др., 1976).
THE DEVELOPMENT OF LATHYRUS J APONICUS WILLD. (FABACEAE) IN THE FIRST YEAR OF LIFE
IN THE CULTIVATED CONDITION
Botanical Garden-Institute, Far East Branch of the Russian Academy of Sciences, Vladivostok
Far East Federal University, Vladivostok
We describe some of the biological peculiarities of the relict species of Fabaceae - Lathyrus japonicas. We pay special attention to the dynamics of germination of cultivated plants, as well as to the energy of germination and its success. The description of developmental stages from the germination to the appearance of the above- and underground parts at virginal period is given.
Калинкина В.А. Развитие Lathyrus japonicas Wild. (сем. Fabaceae) в первый год жизни в условиях культуры / В.А. Калинкина // Вести. ТвГУ. Сер.: Биология и экология. 2016. №4. С. 184-194.
Список литературы Развитие Lathurys japonicus Willd. (сем. Fabaceae) в первый год жизни в условиях культуры
- Арктическая флора СССР. 1986.//Семейство Leguminosae/Сост. А.А. Коробков, М.В. Соколова, Н.Н, Тараскина, Б.А. Брцев. Л.: Наука. Вып. 9. Ч. 2. 188 с.
- Баранов М.П., Бурляева М.О. 1990. Анатомическое строение семенной кожуры некоторых видов рода Lathyrus L.//Исходный материал, генетика, систематика и селекция зерновых бобовых культур (Сб. науч. тр. по прикладной ботанике, генетике и селекции, том 135). Л.: ВИР. С. 125-135.
- Вишнякова М.А., Бурляева М.О., Алпатьева Н.В., Чесноков Ю.В. 2008. RAPD анализ видового полиморфизма рода Lathyrus L. семейства Fabaceae Linl.//Вестник ВОГиС. Т. 12. № 4. С. 595-607.
- Воронкова Н.М., Бурундукова О.Л., Бурляева Е.В. 2003. Морфобиологические особенности семян некоторых видов морских побережий российского Дальнего Востока//Ботанические исследования в Азиатской России. Матер. XI съезда Русского ботанического общества (18-22 августа 2003 г., Новосибирск-Барнаул). Барнаул. Т. 2. С. 32-33.
- Ворончихин В.В. 1997. Анатомия и ультраструктура семенной кожуры некоторых представителей родов Lathyrus и Orobus//Бюлл. ГБС РАН. № 174. С. 115-126.
- Гуленкова М.А. 1968а. Становление жизненной формы в процессе онтогенеза у Lathyrus vernus (L.) Bernh.//Вопросы биологии и экологии доминантов и эдификаторов растительных сообществ: Матер. I межвуз. конф. по биол. и экол. доминантов и эдификаторов естеств. и искусств. фитоценозов, Пермь, 21-26 сентября 1967 г. Уч. зап. Пермь. Т. 64. С. 91-93.
- Гуленкова М.А. 1968б. Становление жизненной формы в процессе онтогенеза у чины гороховидной (Lathyrus pisiformis L.)//Морфология высших растений. М.: Наука. С. 128-141.
- Гуленкова М.А. 1970. Характеристика морфологических преобразований в виргинильный период развития у Lathyrus verus (L.) Bernh. и L. pannonicus Garcke//Рефераты докладов всесоюз. симпозиума по изучению морфологических основ онтогенеза травянистых растений. г. Ставрополь. С. 19-20.
- Гуленкова М.А. 1974а. Становление жизненной формы Lathyrus pannonicus (Kramer) Garcke в онтогенезе//Бюл. Моск. о-ва испыт. прир. Отд. биол. Т. 79. Вып. 4. С. 105-114.
- Гуленкова М.А. 1974б. Сериальные почки у некоторых бобовых//Научн. докл. высш. шк. Биол. науки. Т. 12. С. 55-58.
- Гуленкова М.А. 1977. Становление жизненной формы у Lathyrus sylvestris L. (Fabaceae) в онтогенезе//Ботан. журн. Т. 62. № 5. С. 704-715.
- Калинкина В.А. 2008. Особенности большого жизненного цикла клевера люпиновидного//Вестник ОГУ. № 6. С. 150-155.
- Николаева М.Г., Разумова М.В., Гладкова В.Н. 1985. Справочник по проращиванию покоящихся семян. Л.: Наука. 358 с.
- Нестерова С.В., Холина А.Б., Воронкова А.А. 1997. Всхожесть семян некоторых представителей семейства бобовых после длительного хранения//Животный и растительный мир Дальнего Востока. Сб. науч. тр. Уссурийск. Вып. 3. С. 207-209.
- Павлова Н.С. 1989. Сем. Бобовые -Fabaceae Lindl s. l.//Сосудистые растения советского Дальнего Востока. М.: Наука. Т. 4. С. 313.
- Петухова Л.В., Черноброва О.В., Нотов А.А. 2003. О биоморфологии чины гладкой (Lathyrus laevigatus (Waldst. et Kit.) Gren.)//Ботанические исследования в Тверском регионе. Тверь. Вып. 1. С. 100-107.
- Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах//Тр. БИН АН СССР. Сер. 3. Геоботаника. М.-Л., Вып. 6. С. 7-204.
- Рыжова Н.Н., Бурляева М.О., Кочиева Е.А., Вишнякова М.А. 2007. Использование ITS-последовательностей для оценки таксономических отношений у представителей трибы Vicieae (Adans.) Bronn сем. Fabaceae Lindl.//Экологическая генетика. Том V. № 3. С. 5-14
- Смирнова О.В., Заугольнова Л.Б, Топорова Н.А. 1976. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф//Ценопопуляции растений (основные понятия и структура). М.: Наука. С. 14-43.
- Федченко Б.А. 1948. Род Lathyrus L.//Флора СССР. М.-Л.: АНСССР. Т. 13. 507 с.
- Чефранова З.В. 1987. Род Lathyrus L. Флора Европейской части СССР/Ред. А.А. Федорова. Л.: Наука. Т. 6. С. 147-172.
- Шретер А.И. 1975. Лекарственная флора советского Дальнего Востока. М.: Медицина. 166 с.
- Allkin R., Macfarlane R.D., White R.J, Bisby F.A., Adey, M.E. 1983. The geographical distribution of Vicia. Issue 1. Vicieae Database Project Publication. № 5. 35p.
- Dragendorff G. 1898. Die Heilpflanzen der verschiedenen Völker und Zeiten. Ihre Anwendung, wesentlichen Bestandteile und Geschichte. Stuttgart. 884 p.
- Lathyrus japonicas Willd. Go Botany Discover thousands of Now England plants . https://gobotany.newenglandwild.org/species/lathyrus/japonicas.
- Gurusamy C., Davis P.J., Bal A.K. 2000. Seasonal changes in perennial nodules of Beach pea (Lathyrus maritimus (L.) Bigel.) with special reference to oleosemes//Int. J. Plant Sci. V. 116. No. 4. P. 631-638.
- The Plant List . Режим доступа: http://www.theplantlist.org/tpl1.1/search?q=lathyrus+japonicus.
- Sadao A. 2005. Biological flora of Japan. Japan. P. 366-367.
- Sea Pea //Nature Gate. Режим доступа: http://www.luontoportti.com/suomi/en/kukkakasvit/sea-pea.
