Развитие миксотрофных инфузорий в водоемах Нижней, Средней Волги и Камы как отражение особых условий существования

Автор: Быкова Светлана Викторовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 3-7 т.15, 2013 года.
Бесплатный доступ
В планктоне разнотипных водоемов Средней, Нижней Волги и Камы (водохранилища, притоки, крупные и малые водоемы с различным типом перемешивания водной толщи и уровня трофности, «сульфидные» проточные озера, «болотные» озера и т.д.) зарегистрировано 39 видов миксотрофных инфузорий. Показаны неоднородность их состава, максимальный вклад в численность в стратифицированных водоемах с сульфидным гиполимнионом и максимальный вклад в биомассу в мелких полигумозных озерах. Тенденции развития и экологические предпочтения для миксотрофных инфузорий сильно различаются в зависимости от их адаптивных стратегий. Состав миксотрофных инфузорий и характер их горизонтального и вертикального распределения определяются конкретными абиотическими и биотическими условиями в водоеме и слабо зависят от зональных факторов.
Миксотрофные инфузории, симбиоз, зоохлореллы, стратегии выживания в неблагоприятных условиях, экотопы, стратификация
Короткий адрес: https://sciup.org/148202190
IDR: 148202190 | УДК: 574.52:593.17
The development of mixotrophic ciliates of waterbodies from Bottom, Central Volga and Kama river as indicator of special living conditions
39 species of mixotrophic ciliates from plankton of different reservoirs of Central, Bottom Volga and Kama (reservoirs, inflows, large and small lakes with various type of hashing of water column and of trophic level, "sulphidic" flowing waterbodies, "marsh" lakes, etc.) are registered. The heterogeneity of their structure, the maximum contribution to abundance in the stratified reservoirs with sulphidic hypolimnion and the maximum contribution to biomass in small gumic lakes is shown. Development trends and ecological preferences for the mixotrophic ciliates vary depending on their adaptive strategy.The species composition of mixotrophic ciliates and their horizontal and vertical allocation is defined by specific abiotic and biotic conditions in a reservoir and poorly depends on zonal factors.
Текст научной статьи Развитие миксотрофных инфузорий в водоемах Нижней, Средней Волги и Камы как отражение особых условий существования
М иксотрофия среди инфузорий («приобретенная фототрофия», по: [21]) известна давно; довольно широко и повсеместно распространена: около 23% общего количества морских и пресноводных инфузорий [15] содержат симбиотические водоросли («истинный симбиоз») или выделенн-ные хлоропласты («псевдосимбиоз») [11]. Миксотрофные инфузории более обычны для пресных водоемов, реже для океанов. Применимо к гетеротрофным простейшим, она означает, скорее, «симбиотический консорциум» [14] двух организмов, а не просто смешанный способ питания. Инфузории с «приобретенной фото-трофией» вносят значительный вклад в биоразнообразие водных экосистем. При этом в морских системах чаще распространены инфузории с водорослевыми пластидами, а в пресных – инфузории с внутриклеточными симбионтами. Миксо-трофия зарегистрирована, по крайней мере, в 8 крупных таксонах цилиат: Heterotrichea, Hypotri-cha, Spirotrichia, Litostomatea, Prostomatea, Penicu-lia, Peritrichia и Oligotrichida [21]. Лишь у Oligotrichida миксотрофия связана с сохранением пластид из заглоченных водорослей внутри инфузорий, у остальных же – с водорослевыми симбионтами. В целом, накопилось довольно много сведений по взаимоотношениям хозяина-инфузории и симбионтов [20], описаниям отдельных видов, а также размышлений об адаптивных преимущест-
вах инфузорий- миксотрофов в сравнении с их асимбионтными особями. И явно недостаточно обобщающих, сводных работ, аналогичных обзору Cтокер с соавторами [21], где приводятся накопившиеся данные многих авторов по миксотрофным простейшим разнообразных водных экосистем.
Первоначально развитие миксотрофов связывали с олиготрофными водоемами [16, 18]. Однако еще в середине прошлого века В.А. Догель, и Ю.И. Полянский [6] связывали наличие зоохлорелл в организмах инфузорий не только с потребностью питательных веществ, но и с потребностью улучшения газового режима, особенно в эвтрофных водоемах, что позже подтверждено работами многих авторов [10, 12-14, 19]. Кроме того, миксотрофные инфузории, наряду с другими миксотрофными простейшими, широко распространены и в экстремальных условиях антарктических и субарктических тундровых озер (продолжительное действие низких температур, пониженная освещенность и дефицит питательных веществ) [7, 17]. В целом, неоспорим тот факт, что именно в специфических, часто неблагоприятных, условиях отмечается массовое развитие миксотрофов, обеспечивающее сохранность всего сообщества инфузорий. Тем не менее, в нашей стране данные о разнообразии самих миксотрофных инфузорий, неоднородности их состава, экологии и особенностях развития в разнотипных водоемах разных географических зон, различных экотопах одного и того же водоема (в том числе и узкой зоне хемоклина стратифицированных водоемов), сезонной динамике и т.д. единичны [9]. Поэтому целью работы явилась попытка объединить и обобщить полученные нами на данный момент данные по развитию миксотрофных инфузорий в разнотипных водоемах бассейнов Средней и Нижней Волги и Камы.
РАЙОН И МЕТОДЫ ИССЛЕДОВАНИЯ
Обобщены данные многолетних, начиная с 1998 г., исследований свободноживущих инфузорий в разнотипных водоемах Средней, Нижней Волги и Камы: малые водоемы Самарской Луки, серные и «голубые» озера северо-востока Самарской области, болотные озера с. Ст. Рачейка, городские пруды г. Самара, (Самарская область, 29 озер); крупное оз. Кандры-Куль (респ. Башкорто стан), озера Раифского участка и пойма Саралин-ского участка Волжско-Камского государствен -ного природного биосферного заповедника (респ. Татарстан, 9 водоемов); 5 водоемов респ. Марий Эл; водохранилища Камского каскада (Камское, Воткинское, Нижнекамское, камская ветвь Куйбышевского водохранилища), 28 притоков р. Камы и 2 болотных озера в бассейне р. Вишеры и незарегулированного участка р. Камы (Пермский край). Большая часть озер, за исключением пойменных, - карстового происхождения. Район исследования очень обширен и охватывает область от лесостепной географической зоны (у границы со степной) до тайги.
Таблица 1. Характеристика некоторых исследованных водоемов (составлена по: [3-5]])
|
Водоем |
6 о X X S3 X о о |
к X 1 X X 5 |
S3 в о ч К |
S S S3 X X ю |
Л о X И |
S ” 5 л о X X S |
CU |
X 4 С? >э X О Е^ S >. § s й О а и |
X X ^ О е |
*s3 Ч Ч X о |
X о X о л X о И о >> |
I. Нестратифицированные мелкие и стратифицированные крупные без сульфидного гиполимниона озера
|
Кандры-Куль |
РБ |
с |
1560 |
16,5 |
10 |
984 |
8,7 |
0 |
0,34 |
1,93 |
олиго-, мезо-трофный |
|
Саратовское |
СО |
нс |
1831 |
31,0 |
63 |
231 |
7,8 |
0 |
0,06 / 0,08 |
- |
мезотрофный |
|
водохранилище |
х105 |
II. Полигумозные ацидные ("болотные озера") и зарастающие
|
Харовое |
СО |
нс |
0,04 |
1,9 |
42 |
278 |
7,8 |
0 |
0,32 |
27,20 |
эвтрофный |
|
Серебрянка |
СО |
нс |
0,11 |
0,9 |
51 |
31 |
7,1 |
0 |
0,45 |
39,70 |
эвтрофный |
|
Клюквенное |
СО |
нс |
2,93 |
1,7 |
- |
113 |
8,0/7,5 |
0 |
0,23/0,37 |
28,4/50,3 |
гиперэвтрофный |
|
Лизинка |
СО |
нс |
- |
0,9 |
344 |
142 |
6,6 |
0 |
1,28 |
- |
эвтрофный |
|
Журавлиное |
СО |
с |
- |
1,5 |
202 |
88 |
6,2/6,0 |
0 |
0,050/0,079 |
- |
эвтрофное |
|
Илантово |
РТ |
нс |
4,8 |
2,4 |
160 |
- |
6,9/6,4 |
0 |
0,06* |
- |
|
|
Гнилое |
РТ |
с |
0,8 |
4,5 |
- |
- |
7,5/5,7 |
0,41 |
0,05/0,99* |
- |
эвтрофный |
|
Долгое |
РТ |
с |
0,4 |
12 |
220 |
- |
6,6/5,6 |
0,059 |
0,05/0,52* |
- |
эвтрофный |
III. Серные «голубые» озера
|
Голубое 1 |
СО |
нс |
0,25 |
18,0 - |
2260 |
6,83 |
8,78 |
0,006* |
0,06-0,18 |
олиготрофный |
|
|
Солодовка |
СО |
нс |
- |
1,0 |
- |
1910 |
6,9 |
112,6 |
0,03* |
0,53-4,08 |
мезотрофный |
|
Б. Голубое |
РТ |
нс? |
4,6 |
15,7 |
- |
2743 |
6,3 |
3,2 |
0,018/0,035* |
- |
олиготрофный |
|
Зеленый Ключ |
РМЭ |
нс |
0,3 |
3,8 |
- |
1393 |
7,7 |
3,9 |
0,04* |
- |
олиготрофный |
IV. Стратифицированные (димиктические и меромиктические)
|
Голубое 3 |
СО |
с |
0,27 |
5,0 |
- |
1160 |
7,74 |
60 |
0,015/0,015* |
3,60-42,20 |
мезо-, эвтрофный |
|
Подгорское |
СО |
эс |
0,31 |
2,6 |
74 |
354 |
8,8/8,0 |
следы |
0,52/0,81 |
87,5/204,3 |
гипертрофный |
|
Большое Шелехметское |
СО |
эс |
28 |
6 |
61 |
370 |
7,6/7,7 |
следы |
0,13/0,16 |
56,4/18,5 |
мезо-, эвтрофный |
|
Бездонное |
СО |
с |
0,14 |
8,0 |
56 |
157 |
8,0/6,5 |
0,26 |
0,45/0,76 |
47,3/51,9 |
высокоэвтрофный |
|
Пруд Нижний Ботанический |
СО |
м |
0,55 |
5,7 |
- |
1342 |
- |
163 |
0,16/1,83 |
50,0 |
высокоэвтрофный |
|
Черный Кичиер |
РМЭ |
м |
9,0 |
- |
- |
7,1 |
123 |
0,051/3,37* |
- |
мезотрофный |
|
|
Шунгалдан |
РМЭ |
м |
2 |
13,5 |
- |
2413 |
7,1/7,0 |
309 |
0,002/1,04* |
- |
эвтрофный |
Примечание: РТ - республика Татарстан, РМЭ - республика Марий Эл, СО - Самарская область. Наличие стратификации: нс - отсутствует, эс - эпизодическая, с - продолжительная, м - меромиктический характер перемешивания; прочерк - нет данных.
Все водные объекты (и экотопы в них), огромное разнообразие которых в рамках данной статьи не позволяет рассмотреть особенности развития миксотрофов в каждом из них, были для удобства анализа условно разбиты на группы: I. нестратифицированные и стратифицированные без сульфидного гиполимниона озера, водохранилища и реки с невысокой трофно-стью; II. полигумозные с пониженной кислотностью и мелкие зарастающие высшей водной растительностью водоемы; III. серные, «голубые озера»; IV. эвтрофные стратифицированные (эпизодически стратифицированные, стратифицированные с высоким содержанием сероводорода и сульфидов в гиполимнионе, меромиктические как частный случай таких водоемов) (табл. 1).
Таблица 2. Видовой состав миксотрофных инфузорий бассейнов Нижней, Средней Волги и Камы и их примерная приуроченность к экотопам
|
Виды |
Водоемы и экотопы |
|||
|
I 1 1 2 3 ab |
II 4 1 5 a b a b c |
III 6 a b |
IV 7 1 8 a b c |
|
|
Actinobolina smalli Holt, Lynn & Corliss 1973 Actinobolina wenrichii Wang & Nie, 1933 Askenasia chlorelligera Krainer & Foiss., 1990 Caliptotricha pleuronemoides Phillips, 1882 Climacostomum virens Ehrb., 1833 Coleps hirtus viridis Ehrenberg, 1831 |
• •• • • |
• • • • • • • • |
• |
• • • • • |
|
Cristigera sp. Cyclotrichium viride Gajewskaja, 1933 Cyrtolophosis sp. Didinium chlorelligerum Kahl, 1935 Disematostoma butschlii Lauteborn, 1894 Enchelys simplex Kahl, 1926 Euplotes diadaleos Diller, Kounarius, 1966 Frontonia sp. Histiobalantium natans Clap.& Lachm., 1858 |
• • •• • • • • • • |
• • • • • • • • • • • |
• • • • |
• • • • • • •
|
|
Leptopharynx eurystoma Kahl,1931 Limnostrombidium viride (Stein, 1867) Litonotus sp. Monodinium chlorelligerum Krainer,1995 Ophrydium versatile Muller, 1786 Oxytricha sp. Pelagostrombidium falla x (Zach., 1895) P. mirabile (Penard,1916) Parammecium bursaria (Ehrb.,1831) Pelagohalteria viridis (From., 1876) Foissner, Skogstad & Pratt, 1988 Paracondylostoma setigerum chlorelligerum ? Pelagothrix plancticola Foissner et al., 1995 Pseudoblepharisma tenue (Kahl, 1926) Pseudovorticella fasciculata (Мull., 1773) Rimostrombidium velox (Faure-Fr., 1924) Spathidium viride Kahl, 1926 Spirostomum semivirescens Stentor amethystinus Leidy, 1889 St. polymorphus (O.F. Muller, 1773) Stichotricha secunda Perty, 1852 Stokesia vernalis Wenzich, 1929 Strongylidium lanceolatum Kowal., 1882 Vorticella chlorostigma (Ehrenberg, 1831) |
• • • •• • • • • •• • • •• • • • • • • • • • • |
• • • • • • • • • • • • • • • • • • •
• • • • • • • |
• • • • • |
• • • • • • • • • • • • • • |
Примечание. I. Водоемы пониженной трофности: 1 – олиго-, мезотрофное озеро (а – пелагиаль, b – фиталь); 2 – водохранилища; 3 – притоки; II. зарастающие и полигу-мозные с пониженной кислотностью водоемы: 4 – мелкие макрофитные (а – пелагиаль, b – фиталь); 5 – стратифицированные полигумозные (а – эпилимнион, b – оксиклин, с – горизонты, примыкающие снизу к границе кислородной и бескислородной зон; III. сульфидные, «голубые» озера: (6а – пелагиаль, 6b – бактериальные обрастания и маты, опрес- ненные участки на литорали); IV. эвтрофные и высокоэвтрофные водоемы разной степени стратификации и с высоким содержанием сероводорода: 7 – с устойчивой стратификацией и меромиктические (а – эпилимнион, b – оксиклин, с – ниже границы кислородной и бескислородной зон), 8 – придонный горизонт водоемов с неустойчивой стратификацией.
Для исследования инфузорий отбор проб осуществляли стандартными гидробиологическими методами. С целью учесть гетерогенность распределения сообщества инфузорий в разнотипных местообитаниях, пробы отбирали в разных экотопах водоема: пелагической части, открытой литорали, фитали и т.д. Кроме того, в стратифицированных водоемах отбор проб производили с разных горизонтов: шаг отбора составлял 0,5-1 м, а в области градиентов (оксиклина, термоклина, хемоклина и т.д.) и прилегающих зонах некоторых водоемов – 0,1 м. Количественный учет проводили на фиксированном сулемой материале, видовую идентификацию – в живом виде и применяя стандартные гистохимические (импрегнации серебром, окрашивания ядер по Фельгену) и протозоологические методы. Ординацию полученных данных в зависимости от факторов среды проводили на основе Canonical Correspondence Analysis (CCA) с помощью программы Canoco for Windows 4,5.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Всего в планктоне зарегистрировано 39 видов миксотрофных инфузорий (12,5% от общего числа зарегистрированных нами видов) (Табл. 2). При этом, в список вошли, помимо эупланктон-ных и эпипланктонных, бентосные и некоторые перифитонные виды, попавшие в планктонные пробы в результате смыва (в данной работе исследованиями прикрепленных миксотрофов на естественных субстратах и стеклах обрастания специально не занимались).
Состав инфузорий, содержащих симбиотические водоросли или выделенные хлоропласты, довольно неоднороден. Явно выделяются «эпилим-нические» аэрофильные миксотрофы (Rimostrom-bidium velox, Limnostrombidium viride, Pelagohalte-ria viridis, Pelagostrombidium mirabile, Enchelys simplex, Disematostoma butschlii, Actinibolina smalli, Spathidium viride и др.) со стратегией обеспечения себя необходимыми питательными элементами, и микроаэрофильные и анаэробные миксотрофы (Pelagothrix plancticola, Frontonia sp., Euplotes diadaleos, Histiobalantium natans, Coleps hirtus viridis и др.) со стратегией обеспечения себя кислородом. Первые встречаются в водоемах с пониженным (довольно условно) уровнем трофно-сти; неустойчивой стратификацией; в эпилимнионе стратифицированных водоемов; в водоемах с действием неблагоприятных факторов, (за исклю- чением дефицита кислорода), таких как цветность, ацидность, колебание уровня воды и т.д.; вторые предпочитают микроаэробные и анаэробные условия стратифицированных и меромиктических эвтрофных водоемов. Наиболее распространенные миксотрофные инфузории первой группы (родов Limnostrombidium и Pelagostrombidium) содержат выделенные хлоропласты, в то время как у большинства миксотрофов второй группы, за небольшим исключением (например, Histiobalantium natans, имеет «приобретенную фототрофию» за счет выделенных хлоропластов [13]), эндосимбионты - Chlorella.
Особенности развития миксотрофных инфузорий в разнотипных условиях (табл. 3) . По вкладу в общую численность (до 51-75%) развитие миксотрофов максимально в эвтрофных стратифицированных водоемах с сульфидным гиполимнионом (пруд Нижний Ботанический, озера Голубое 3 и Бездонное); по вкладу в общую биомассу (до 91-98%) - в глубоких полигумозных водоемах (оз. Гнилое, Долгое, Карасиха) и в сообществах зарослевой части мелких зарастающих водоемов (оз. Харовое, фиталь оз. Журавлиного) (табл. 3). Во всех озерах вклад миксотрофов в биомассу как правило выше, чем вклад в численность. Индекс плотности (показатель, интегрирующий в себе абсолютные значения как численности, так и биомассы) максимален в фитали оз.
Журавлиное, где в массовом количестве развиваются крупные полуподвижные Stentor amethystinus . Наиболее богаты в плане количества видов миксотрофные инфузории совершенно различных водоемов: олиго-, мезотрофного оз. Кан-дры Куль, высоко-эвтрофного меромиктического пруда Нижний Ботанический и эвтрофного ацид-ного оз. Журавлиное (табл. 3).
I. Водоемы пониженной трофности. Типичный водоем данной группы - крупное олиго- -мезотрофное оз. Кандры-Куль. Особенность развивающихся в нем миксотрофов заключается в том, что они представлены, в основном, мелкими и средними по размерам эупланктонными видами (табл. 3, рис. 1). Чаще других и во всех экотопах встречаются (в порядке убывания): Pelagostrom-bidium mirabile ; Pelagohalteria viridis, Monodinium chlorelligerum, Askenasia chlorelligera (рис. 1). В пелагиали и открытой литорали вклад миксотро-фов в численность и биомассу выше, чем в заросшем мелководье. Развитие миксотрофов весной начинается с прибрежной зоны, перемещаясь летом в пелагическую, но не самую глубоководную (а, следовательно, лучше освещенную) часть водоема. В целом, вклад миксотрофов максимален весной и летом (57% и 50% от общей численности), к осени же значительно снижается по всей акватории озера (5,5%).
пелагиаль
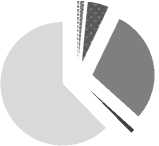
Askenasia chlorelligera
Pelagostrombidium mirabile
Вклад в биомассу, % открытая литораль

Monodinium chlorelligerum прочие миксотрофы
Pelagohalteria viridis прочие инфузории
Частота встречаемости, %
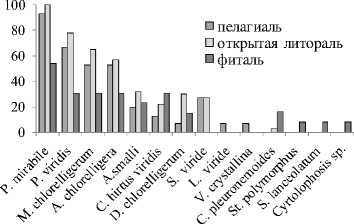
Рис. 1. Доля миксотрофных инфузорий в биомассе сообщества планктона (средние показатели май-июль-сентябрь) и их частота встречаемости в оз. Кандры-Куль в 2012 г.
В водохранилищах, по сравнению с другими водоемами, роль миксотрофов менее значительна. Так, вклад в численность и биомассу сообществ инфузорий планктона в Саратовском водохранилище составляет 5% и 6,4% (табл. 3), соответственно, а в водохранилищах камского каскада, включая камскую ветвь Куйбышевского водохранилища, - 5,4-15% (в среднем, 9,6%) от общей численности и 5,4-28% (в среднем, 14%) от общей биомассы. Кроме того, довольно часто некоторые виды, например Disematostoma butschlii, Stokesia vernalis, Frontonia leucas развиваются вообще без симбиотических водорослей, однако роль таких факультативных миксотрофов в сообществе незначительна. Так же, как и в оз. Кандры-Куль, вклад миксотрофов в развитие всего сообщества инфузорий водохранилищ выше в открытой части, по сравнению с зарослевой: доля миксотрофов по численности в пелагили и фитали составляет 29 и 14%, доля по биомассе 12 и 8%, соответственно. В водохранилищах основные миксотрофы представлены: Limnostrombidium viride (1,8% от общей численности); Rimostrombidium velox (1,8%); P. mirabile (0,9%); Spathidium viride (0,5%) (Саратовское водохранилище); Pelagostrombidium mirabile (2,2%) Spathidium viride (1,6%), Didinium chlorelligerum (1,4%), Cyclotrichium viride (0,9%), Enchelys simplex (0,6%), Monodinium chlorelligerum (0,6%) (камские водохранилища).
Таблица 3. Вклад миксотрофных инфузорий в количественные показатели развития сообществ инфузорий р азнотипных вод оемов бассейнов Нижней, Средней Волги и Камы
|
Водоем |
N, тыс.экз. /м3 |
B, мг/м3 |
√N×B |
n |
Основные виды миксотрофов* |
|
абс. % |
абс. % |
Нестратифицированные и стратифицированные крупные без сульфидного гиполимниона озера
|
Кандры-Куль |
356 |
20 |
6,96 |
39 |
49,8 |
19 |
Pelagostrombidium mirabile (14% / 82%); Pelagohalteria viridis (6% 80%); A skenasia chlorelligera (0,3% / 4,2%) |
|
Cаратовское водохранилище |
2,3 |
5 |
0,16 |
6 |
0,6 |
5 |
Limnostrombidium viride (1,8%/11,1%); Rimostrombidium velox (1,8% / 3%); P. mirabile (0,9% / 3,4%); Spathidium viride (0,5% / 3%) |
Полигумозные ("болотные") и зарастающие озера
|
Харовое |
911 |
61 |
1088 |
98 |
994,4 |
7 |
Stentor amethystinus (16,5%/53%), Vorticella chlorostigma (2,4% / 25%), P. mirabile (2,0%/15%), Euplotes diadaleos (1,4% / 21%); Paramecium bursaria (0,2% / 1,1%); S. viride (0,3% / 1,9%); Pelagothrix plancticola (у дна 26% / 45%) |
|
Серебрянка |
167 |
5,4 |
23,5 |
13 |
63,2 |
7 |
Actinobolina wenrichii (1,5% / 7,5%); P. plancticola (0,8% / 22%); Para-condylostoma setigerum chlorelligerum (1% / 11%); Stentor polymorphus (<0,1% / 1,1%); P. bursaria (0,5% /9%); Frontonia sp. (0,2% / 1,4%); Spathidium viride (0,2% / 1,4%) |
|
Клюквенное |
148 |
28 |
10,3 |
40 |
38,9 |
8 |
L. viride (20% / 53%); R. velox (3,5% / 56%); Actinobolina smalli (1,8% / 5,8%); P. mirabile (1,2% / 4,8%); Stokesia vernalis (0,3% / 2,3%); Disema-tostoma butschlii (0,5% / 2,9%); Enchelys simplex (0,4% / 1,2%) |
|
Лизинка |
38 |
54 |
2,7 |
51 |
10,1 |
4 |
L. viride (37,4% / 50); P. mirabile (8,4% / 38,9%); R. velox (5,6% / 27,8%); P. bursaria (2,8% / 9,1%) |
|
Журавлиное |
251 |
10 |
4,9 |
19 |
35,2 |
5 |
C. hirtus viridis (6,3 %); P. mirabile (1,3%); P. viridis (1,3 %); A. smalli |
|
(пелагиаль) |
(0,9%) |
||||||
|
Журавлиное |
3186 |
17 |
2244 |
96 |
2673 |
19 |
S. amethystinus (10,1%/45,2%); P. mirabile (1,9%/2,7%); R. velox (1,5% / |
|
(фиталь) |
3,2%); P. viridis (1,5% / 2,8%); Vorticella chlorostigma (0,1%/ 0,3%); Leptopharynx eurystoma (Kahl,1931) (<0,1%); P. setigerum chlorelligerum (<0,1%) |
||||||
|
Гнилое |
1856 |
54 |
1278 |
97 |
1541 |
4 |
D. butschlii (35% / 43%); Frontonia sp. (12% / 43%); P. mirabile (6% / 27%); H. natans (<0,1% ); R. velox (0,3%) |
|
Долгое |
472 |
46 |
194,7 |
91 |
303,2 |
10 |
Pseudovorticella fasciculata (17 / 30%); D. butschlii (6% / 10 %); R. velox (8% / 11%); P. viridis (5% / 21%); H. natans (4% / 45%); S. amethystinus (2% / 7%); P. plancticola (2% / 26%); Frontonia sp. (1% / 14%); L. viride (0,2% / 0,3%) |
|
Серные «голубые» озера |
|||||||
|
Голубое 1 (П) |
1 |
41 |
0,2 |
75 |
0,4 |
4 |
E. diadaleos (21%); P. bursaria (10,3%); H. natans (5,2%); |
|
Голубое 1 (Л) |
16 |
43 |
- |
- |
- |
3 |
P. bursaria (21,6%); Monodinium chlorelligerum (16,2%); E. diadaleos (5,4%) |
|
Солодовка (пр) |
7 |
3 |
0,9 |
9 |
2,6 |
2 |
C. hirtus viridis (2%); P. bursaria (1%) |
|
Солодовка (з) |
12 |
11 |
12,5 |
21 |
12,2 |
2 |
Frontonia sp. (11,3%); C. hirtus viridis (0,7%) |
|
Большое |
Стратифицированные (димиктические и меромиктические) |
||||||
|
104 |
14 |
9,7 |
26 |
28,9 |
10 |
R. velox (6% / 41%); L. viride (4% /271%); P. mirabile (0,6% / 8%); E, |
|
|
Шелехметское |
simplex (0,5% / 5%); D. butschlii (<0,1 / 4%) |
||||||
|
Подгорское |
695 |
32 |
79,7 |
11 |
232,8 |
15 |
P. plancticola (20%); L. viride (6%); H. natans (1,9%); E. diadaleos (0,9%); R. velox (0,8%); A. smalli (0,3%); D. butschlii (<0,1%) |
|
Бездонное |
5175 |
44 |
884,1 |
68 |
2139 |
11 |
P. plancticola (36%); Frontonia sp. (2,8%); E. diadaleos (2,5%); D. butsch-lii (2%); H. natans (0,2%); E. simplex (0,2%); A. smalli (0,3%) |
|
Голубое 3 |
1016 |
62 |
129,7 |
80 |
363 |
10 |
P. plancticola (27% / 76%); P. viridis (22% / 41%); C. hirtus viridis (5,8%) / 32%); H. natans (3% / 14%); R. velox (2% / 9%); A. chlorelligera (1,4% / 8%); D. butschlii (0,4% / 3%); E. diadaleos (0,1% / 0,6%) и др. |
|
Пруд Н. Бота- |
3407 |
75 |
456,7 |
53 |
1247 |
16 |
P. plancticola (56% / 92%); H. natans (10% / 79%); E. diadaleos (4% / |
|
нический |
15%); L. viride (0,9% / 27%); C. hirtus viridis (0,6% / 34%); D. chlorel-ligerum (0,2% / 16%) и др. |
||||||
|
Ч. Кичиер |
4 |
0,5 |
0,1 |
0,4 |
0,5 |
1 |
C. hirtus viridis (0,5% / 4, 8%) |
|
Шунгалдан |
119 |
51 |
7,2 |
35 |
29,3 |
6 |
P. viridis (32 / 52%); Rimostrombidium sp. (5% / 28%); P. mirabile (4,6% / 16,3%); A. chlorelligera (4% / 11%); C. hirtus viridis (0,7% / 1,3%); Оxy-tricha sp. (0,3 % / 4,7%) |
Примечание. * – в скобках указан вклад миксотрофов (средний / максимальный, в % от общей численности инфузорий); N – численность, B – биомасса, n – количество видов, √N×B – индекс плотности; П – пелагиаль, Л – литораль, пр – проточный экотоп, з – заросле-вый экотоп
В притоках (в основном, малых реках) вклад миксотрофов в количественные характеристики еще ниже (6,2% в численность и 3,2% в биомассу). Основная роль среди миксотрофов принадлежит Coleps hirtus viridis (4% от общей численности). Кроме того, здесь более значимы, по сравнению с водохранилищами, Monodinium chlorelligerum (4,7%) и Pelagohalteria viridis (4%). В Чусовском заливе как экотонной зоне представлены «миксотрофные доминанты» как водохранилищ - Pelagostrombidium mirabile (1,6% от общей численности), так и их притоков - C. hirtus viridis (0,6%) и Monodinium chlorelligerum (0,5%).
В целом, особенностью миксотрофов данной группы водоемов является то, что они представлены эупланктонными видами со стратегией обеспечения себя питательными веществами. Характер связи с симбионтами для некоторых из них может быть факультативным, а некоторые (в основном, олиготрихи) содержат выделенные хлоропласты.
-
II. Зарастающие олиго-, мезогумозные (озера Харовое, Серебрянка, Клюквенное ) и полигу-мозные с пониженной кислотностью водоемы (мелкие оз. Лизинка, Журавлиное и глубокие стратифицированные оз. Гнилое и Долгое). В по-лигумозных (темноводных) (цветность 100-250°Pt) и сильно гумифицированных (цветность >250°Pt) водоемах из-за затенения вследствие высокой цветности складываются неблагоприятные трофические условия, поскольку, несмотря на то, что болотные озера (например, Журавлиное) отличаются большим количеством органического вещества, доля в них легкоокисляемого вещества невелика [5]. Поэтому в этих водоемах тоже функционально более значимы миксотрофы с первой стратегией («обеспечения себя питательными веществами в водоемах пониженной трофности»).
Особенность фауны мелких макрофитных озер состоит в том, что в них миксотрофы представлены средними и довольно крупными пери-фитонными (полуподвижными Stentor polymorphus, Stentor amethystinus и прикрепленными Vorticella chlorostigma) и бентосными видами (Paramecium bursaria, Spirostomum semivirescens, Climacostomum virens), попадающими в планктон за счет смыва с макрофитов и с поверхности грунтов, куда проникает достаточное для симбионтов количество света (табл. 3). В отличие от водоемов первой группы, в фитали данных водоемов, по сравнению с открытой водой, вклад миксотрофов в биомассу всего сообщества значительно выше за счет крупных видов. При этом их распределение по акватории водоема может быть очень агрегированным (мозаичным). Например, это характерно для популяции Stentor amethystinus в оз. Харовое и Журавлиное. Так, в пелагиали озера Журавлиное данный вид не обнаружен, а в планктоне зарослей макрофитов его численность распределена крайне неравномерно: в центре зарослей погруженно-водного макрофита (рдеста) она составила 2380 и 16580 тыс. экз./м3, на границах открытой воды с зарослями рдеста - 40 и 380 тыс. экз./м3, соответственно, в зарослях прибрежно-водного макрофита (тростника) - 60 тыс. экз./ м3. Интересны находки довольно редких видов Vorticella chlorostigma (оз. Харовое, Журавлиное), Actinobolina wenrichii, Paracondylostoma setigerum chlorelligerum (оз. Клюквенное), Leptopharynx eurystoma, Spirosto-mum semivirescens (оз. Журавлиное), Pseudoble-pharisma tenue var. viride (болота Моховое, Узи-лово) . В целом, вклад миксотрофов в количественные показатели всего сообщества в данного типа водоемах колеблется в широких пределах: от 5% в численность и 13% в биомассу в оз. Клюквенное и до 61% и 98% , соответственно, в оз. Ха-ровое. При создании аноксичных условий в придонной воде мелких водоемов увеличивается роль миксотрофов со второй стратегией (обеспечение кислородом). Например, в оз. Харовое возрастает (до 45%) вклад Pelagothrix plancticola в общую численность.
В качестве особенности состава миксотро-фов в глубоких стратифицированных полигумоз-ных озерах (оз. Долгое, Гнилое) необходимо отметить значительную роль факультативных мик-сотрофов, которые, в отличие от водохранилищ-ных видов, в данном водоеме всегда с зоохлореллами, и миксотрофов, содержащих выделенные хлоропласты. Так, в приповерхностных горизонтах оз. Долгое 27% численности и 37% биомассы приходятся на Pelagostrombidium mirabile («приобретенная фототрофия» за счет выделенных хлоропластов), а у границы кислородной и бескислородной зон в массовых количествах развиваются Disematostoma butschlii (42% от общей численности и 46% от общей биомассы) и Frontonia leucas с зоохлореллами (14% и 54%, соответственно).
-
III. В серных, «голубых озерах» действует целый комплекс «неблагоприятных» для инфузорий факторов: пониженная трофность, низкая температура, высокая проточность, повышенная минерализация, высокая концентрация сероводорода и сульфидов и т.д. В оз. Голубое 1 развитие всего сообщества инфузорий и, миксотрофов в частности, в пелагическом планктоне сильно ограничено и полностью отсутствует в области источников. Тем не менее, в пелагиали, в толще зарегистрированы единичные экземпляры Euplotes diadaleos и Histiobalantium natans , что составило 42% от общей численности. Основное развитие инфузорий в таких экосистемах приурочено к бактериальным матам и обрастани-
- ям в литоральной зоне, при этом инфузории используют данный специфический субстрат в качестве биотопа, т.е. здесь не идет речь об истинно перифитонных (прикрепленных) видах, за исключением Ophrydium sp., Pseudovorticella sp. Основу сообщества здесь составляют анаэробные сапропелевые виды. Наряду с ними в слизистых матах на харе с преобладанием нитчатых зеленых бактерий и бесцветных серобактерий можно встретить немногочисленные миксотрофы Paramecium bursaria, Histiobalantium natans, Monodinium chlorelligerum, Ophrydium sp. (оз. Голубое 1 и Солод овка); в воде вокруг матов с преобладанием пурпурных бактерий и цианобактерий -Climacostomum virens (оз. Коржовское). Практически все миксотрофы в данных условиях - виды с «кислородной стратегией». В сравнительно более опреснённых экотопах водоемов с меньшим содержанием сероводорода присутствуют Coleps hirtus viridis и Frontonia sp. В общем, в серных озерах, по сравнению с маломинерализованными аэробными озерами, значительно увеличена доля по числу видов (но не по численности) миксотро-фов: в серных озерах она составляет 37%, в «несерных» - 4%.
-
IV. Развитие миксотрофных инфузорий в эвтрофных и высокоэвтрофных водоемах с разной степенью стратификации и высоким содержанием сероводорода и сульфидов уже рассматривалось нами ранее [1] . Показано, что даже в пределах одного водоема состав миксотрофов неоднороден. В эпилимнионе они представлены видами с «пищевой» стратегией («обеспечения себя питательными веществами»): Limnostrombi-dium viride, Pelagostrombidium mirabile, Pelagohal-teria viridis, Rimostrombidium velox, Enchelys simplex, Actinibolina smalli и др. В зоне градиентов химических факторов (оксиклин в термически стратифицированном и хемоклин в меромиктическом водоемах) и прилегающих к ним областях, а также в придонных горизонтах неглубоких водоемов с эпизодической неустойчивой стратификацией (оз. Подгорское), в которых свет проникает до дна, в массовом количестве развиваются мик-сотрофы со второй («кислородной»)стратегией: C. hirtus viridis (в верхней части оксиклина), P. plancticola , H. natans , E. diadaleos , и Frontonia sp. (под границей кислородно-бескилородной зон), P. bursaria Frontonia sp. ( vernalis ?) (у дна в мелких водоемах) и т.д.. Все эти виды (за исключением, возможно, Frontonia sp.) являются облигатными миксотрофами.
Абсолютные максимумы численности (206540 экз./л, 84% от общей численности) и биомассы (7264 мг/м3, 51%) миксотрофов были зарегистрированы в период стратификации в высокоэвтрофном меромиктическом пруду. В подобных экосистемах миксотрофы так или иначе функционально значимы (13-87% от средней в толще воды общей численности и 4-91% от биомассы) на протяжении всего годового цикла, за исключением периода перемешивания воды в миксолимнионе (разрушение оксиклина), когда вклад миксотрофов минимален (2,8% от численности и 1,9% от биомассы) и в конце подледного периода (март), когда значительные толщина льда и снежный покров препятствуют проникновению света и протеканию фотосинтеза подо льдом и, соответственно, развитию миксотрофов. Все это позволяет считать миксотрофных инфузорий неотъемлемой чертой данного типа водоема, по аналогии с аноксигенными фототрофными бактериями, являющимися «визитной карточкой» меромиктических водоемов [Рогозин].
Особенностью водоемов данной группы является и то, что в них часто биомассы миксотрофных инфузорий сопоставимы с биомассами бактерий и фитофлагеллят, в отличие от других водоемов, где биомассы всего сообщества инфузорий на несколько порядков меньше биомассы бактерий [2]. Интересны взаимоотношения миксотрофных инфузорий и с другими компонентами фототрофной составляющей сообщества одноклеточных организмов в целом. Максимумы развития инфузорий-миксотрофов всегда располагаются над максимумами фототрофных аноксигенных (пурпурных и зеленых серных) бактерий и часто совпадают или лежат под максимумами других. миксотрофных простейших (например, криптофитовых или динофитовых), определяя общий хлорофильный максимум [2].
В целом, такие виды, как P. plancticola, Frontonia sp., E.diadaleos, H. natans, C. hirtus viridis широко распространены и составляют основу численности в микроаэробных и анаэробных условиях устойчиво стратифицированных и меромиктических эвтрофных водоемов не только Самарской области, (степная и лесостепная зоны), но и некоторых водоемов Англии, Испании, Литвы, Эстонии и др. [13, 22], что свидетельствует о космополитизме данной фауны. Тем не менее, в исследованных нами водоемах со схожими условиями в республиках Марий Эл и Татарстан (лесная зона) эти виды либо отсутствуют, либо вклад их очень незначителен (табл. 3). Так, в меромиктическом оз. Черный Кичиер, где велико содержание сероводорода и сульфидов (табл. 1) и отмечается ярко выраженная бактериальная пластина, казалось бы, созданы все условия для миксотрофов, однако их развитие сильно ограничено (3,7% от общей численности) и представлены они лишь C. hirtus viridis . Для выяснения причины таких различий необходимы дальнейшие исследования.
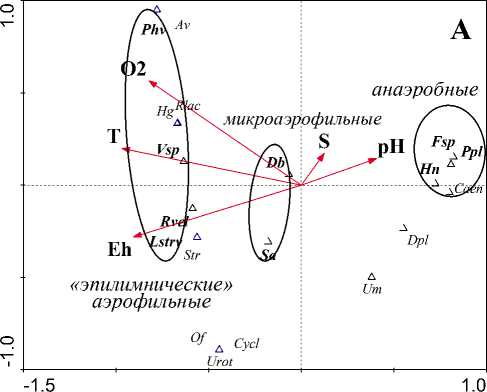
Ординация сообществ инфузорий пелагического планктона разных стратифицированных водоемов – полигумозного озера (рис. 2А) и меромиктического пруда (рис. 2В) – в пространстве факторов среды с использованием канонического анализа соответствий CCA показала, что распределение миксотрофов в толще воды сходно и обусловлено, в первую очередь, их экологическими требованиями (рис. 2). Так, «эпилимнические» аэ-рофильные миксотрофы предпочитают горизонты с повышенными температурой, содержанием кислорода и окислительно-восстановительного потенциала. Анаэробные миксотрофы – горизонты с повышенным содержанием фосфора и минерализацией и т.д. Промежуточное положение занимают микроаэрофильные виды. При этом наблюдается
взаимозаменяемость близких видов, имеющих сходные экологические требования и функции, в близких сообществах: так, в эпилимнионе оз Долгое развивается Limnostrombidium viride, а в эпи-
лимнионе оз. Гнилое
–
Pelagostrombidium
mirabile. Другой пример: в сильно различающихся (по трофности и кислотности) водоемах, но в сходных (по газовому режиму) условиях у верхней границы оксиклина, тоже отмечаются близкие виды Stentor amethystinus (оз. Долгое) и S. polymorphus (пруд Н. Ботанический) (рис. 2А, В). Вероятно, они выполняют сходные функции в условиях, соответствующих их экологическим оптимумам.
микроаэрофильные D L c o
Mb
Fl Me Hr
Asph
В
Stpol
Uro2
Ed Hn Ea
Ppl2 Ppl
Miner. Prov
Per Cm
UtSt
Ptot Pl
Fsp анаэробные
Vn
Ch
O2
«эпилимнические» аэрофильные sm Eh
TrapLac r Rstr1
'д Lstrv pH
Hg
Af T
Pstrf
Phv Rstr2
Csp
Spa
Uro1 A Bep
Av^ Lisp
Rstrv
-1.0
1.5
Рис. 2. Ординационные диаграммы ССА связи факторов среды и состава миксотрофных инфузорий в планктоне стратифицированных водоемов: полигумозного оз Долгое (А) и с признаками меромиктии пруда Нижний Ботанический (В). Обозначения: О 2 – содержание кислорода, Miner. – минерализация, Ptot – содержание общего фосфора; коды видов-миксотрофов (выделены жирным шрифтом): Asm – Actinobolina smalli , Chv – Coleps hirtus viridis , Ed– Euplotes diadaleos , Fsp – Frontonia sp., Hn – Histiobalantium natans , Lstrv – Limnostrombidium viride , Phv – Pelagohalteria viridis,Pstrf – Pelagostrombidium falla x, Rstrv –
Rimostrombidium velox , Sa – Stentor amethystinus , Spa – Spathidium viride, Stpol – Stentor polymorphus
Стратегия МИКХО1рифиН
I Получение легкодоступной opiлипки
0=
трофностъ НЯ
I Илбсттшпе пресс* кислорад» - ’’пш™» 1
II. Получение
z ..... v „адшумозныс
I мезогумозньге I ,
^ серные, «голубые» озера
водоемы noHKASHwll 1р^т^стн]^ктря^г6-Qjwro^2 w^oTpo^we)"
ЭГОЛЮТ1Ы'
Petagos rm и i buTiuni Limnosmwbktiu»! nrkte Mojeo*air<»in Wzh^ RVMOSTTOMlbldlUlM 1 A'fll S’atesia 1 ®tmIu Enchchi simplex JcJimfboItna smaJh XsJtenasra cManlbgcra
i ne amran'- BBS
Mill-. ЫСПШНМШЮН
^KIi^MHLlbHM** млты и оОрастшны
✓ мста-гшюлкмпннон, хемоклин высокоэнтрофньтх водоемов
придонные горепошы atuucuMiieciai сгро.тифи1шроыю1Ы1 псооемод
Ваши:
DiMfmatoTtoma bulschlb Psmdovorhcrlla fasciculate Pdagvhaittna
vjndu SJtviror am
СДОпчйшм wwM РдяттАчит bursana.
Pxeudovartictila /ascxvlata
Виды: P^ay-ch1 1 y plcvvcncola Eupkn^t dlatktiex» HiMiotxricwTluw natat# ^/Muub/iiNm Man>JhpfrtjM Coieps hirrus wutu Eranionxi sp.
Dzhmum chloiTlJigerum

Рис. 3. Схематическое изображение развития различных по жизненным стратегиям миксотрофных инфузорий водоемов Средней Волги и Камы
ЗАКЛЮЧЕНИЕ
Таким образом, на основании обобщения данных по исследованию миксотрофных инфузорий в разнотипных водоемах Средней, Нижней Волги и Камы можно утверждать следующее: зарегистрировано 39 видов-миксотрофов (только в планктоне); установлено максимальное количественное развитие (индекс плотности) в полигумозном и эвтрофных стратифицированном димиктическом и меромиктическом водоемах, при этом максимальный вклад в численность – в стратифицированных, а в биомассу – в мелких зарастающих полигумозных водоемах. Составлена схема приуроченности разных видов миксотрофных инфузорий к различным водоемам, экотопам и условиям в них (рис. 3). Тем не менее, привязка миксо-трофов с различными адаптивными стратегиями (получение питательных веществ или получение от симбионтов кислорода) к определенному типу водоема или экотопу довольно условна, поскольку вид реагирует на локальные условия, которые при определенных обстоятельствах бывают схожими в разных экотопах и разнотипных водоемах, возможно разных географических зон.
В целом, показана неоднородность состава миксотрофных инфузорий в разнотипных водоемах Волжского бассейна. Тенденции развития и экологические предпочтения для мик-сотрофных инфузорий сильно различаются в зависимости от их адаптивных стратегий. Виды одной стратегии взаимозаменяемы. Характер связи с симбионтами (факультативный или облигатный), возможно, определяется типом стратегии: факультативная связь чаще наблюдается в водоемах пониженной трофии: водохранилищах и полигумозных стратифицированных озерах. Состав миксотрофных инфузорий и характер их горизонтального и вертикального распределения определяются конкретными абиотическими и биотическими условиями в водоеме и слабо зависят от зональных факторов.
Список литературы Развитие миксотрофных инфузорий в водоемах Нижней, Средней Волги и Камы как отражение особых условий существования
- Быкова С.В. Миксотрофные инфузории в стратифицированных водоемах бассейна Средней и Нижней волги // Мат-лы X Междунар. науч.-практ. конференции «Татищевские чтения: актуальные проблемы науки и практики» // Актуальные проблемы экологии и охраны окружающей среды. Тольятти: Волжский университет им. В.Н. Татищева, 2013. С. 19-27.
- Быкова С.В., Уманская М.В. Микромасштабное распределение простейших и бактерий в хемоклине меромиктического водоема//Вода: химия и экология. 2012. № 9. С 43-49.
- Горбунов М.Ю., Уманская М.В. Абиотические условия в водной толще озер//Жариков В.В., Горбунов М.Ю., Быкова С.В., Уманская М.В., Шерышева Н.Г. Экология сообществ бактерий и свободноживущих инфузорий малых водоемов Самарской Луки./Под редакцией д.б.н. В.В. Жарикова. Тольятти, 2007. C. 17-41.
- Горбунов М.Ю., Уманская М.В. Абиотические условия водной толщи озер Самарской области//Жариков В.В., Горбунов М.Ю., Быкова С.В., Уманская М.В., Тарасова Н.Г., Буркова Т.Н., Шерышева Н.Г., Ротарь М.Ю. Протисты и бактерии озер Самарской области/Под редакцией д.б.н. В.В. Жарикова. Тольятти: Кассандра, 2009. C. 33-60.
- Горохова О.Г, Номоконова В.И. Фитопланктон и условия его развития в болотных озерах юга лесостепного поволжья (Самарская область)//Изв. СНЦ РАН. Т.20, № 1, 2011. С. 68-75.
- Догель В.А., Полянский Ю.И., Хейсин Е.М. Общая протозоология. Изд-во АН СССР. 1962. 592 с.
- Лаврентьев П.Я. Сообщества инфузорий субарктических тундровых озер. Особенности структуры и развития, роль в экосистеме и реакция на антропогенное воздействие//Автореф. дис. …канд. биол. наук. СПб, 1991. 21 с.
- Рогозин Д.Ю., Зыков В.В., Трусова М.Ю., Белолипецкий В.М. Фототрофные серные бактерии в меромиктических озерах Хакасии: пространственные распределения и сезонная динамика//X Съезд Гидробиологического общества при РАН. Тез. докл. (г. Владивосток, 28 сентября -2 октября 2009 г.)/отв. ред. Алимов А.Ф., Адрианов А.В. Владивосток: Дальнаука, 2009. С. 338-339.
- Хлебович Т.В. Вклад хлорофиллсодержащих миксотрофных инфузорий в общую автотрофную биомассу в пресноводных водоемах//Структурно-функциональная организация пресноводных экосистем разного типа. Тр. ЗИН РАН. 1999. Т. 279. С. 93-99.
- Berninger U-G., Finlay B.J., Canter H.M. The Spatial Distribution and Ecology of Zoochlorellae-Bearing Ciliates in a Productive Pond//J. Protozool. 1986. 33, № 4. P. 557-563.
- Dolan J. Mixotrophy in ciliates: a review of Chlorella symbiosis and chloroplast retention//Marine Microbial Food Webs. 1992. 6(2). P. 115-132.
- Esteban G.F., Fenchel T., Finlay B.J. Mixotrophy in Ciliates//Protist. 2010. V. 161, Issue 5 P. 621-64.
- Esteban G.F., Finlay B.J. Morphology and Ecology of the Cosmopolitan Ciliate Prorodon viridis//Archiv für Protistenkunde. 1996. Bd. 147. P. 181-188.
- Finlay B.J. Protist taxonomy: an ecological perspective//Phil Trans. R. Soc. Lond. B 2004. 359. P. 599-610.
- Foissner W., Berger H., Schaumdurg J. Identification and Ecology of Limnetic Plancton Ciliates. Informationsberichte des Bayer. Landesamtes für Wasserwirtschaft, Heft 3/99. 1999. 793 pp.
- Hecky, R. E. & Kling, H. J. 1981. The phytoplankton and protozooplankton of the euphotic zone of Lake Tanganyika: species composition, biomass, chlorophyll content, and spatio-temporal distribution. Limnol. Oceanogr., 26:548-564.
- Laybourn-Parry J. Survival mechanisms in Antarctic lakes//Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2002. 357(1423). P. 863-869.
- Modenutti B.E., Balseiro E.G. Mixotrophic ciliates in an Andean lake: dependence on light and prey of an Ophrydium naumanni population//Freshwater Biol. 2002. V. 47. P. 121-128.
- Sanders R. Mixotrophic Protists in Marine and Freshwater Ecosystems//J. Protozool. 1991. V. 38, № 1. P. 76-81.
- Stabell Tr, Andersen T., Klaveness D. Ecological significance of endosymbionts in a mixotrophic ciliate -an experimental test of a simple model of growth coordination between host and symbiont//J. of Plankton Research 2002, V. 24. N. 9. P. 889-988.
- Stoecker D.K., Johnson M.D., de Vargas, Not F. Acquired phototrophy in aquatic protists//Aquat. Microb. Ecol. 2009. Vol. 57. P. 279-310.
- Zingel P. Vertical and seasonal dynamics of planktonic ciliates in a strongly stratified hypertrophic lake//Hydrobiologia. 2005. 547.
