Реакции дыхания на активацию ГАМКа- и ГАМКв-рецепторов парафациальной респираторной группы специфическими агонистами

Автор: Ковалева Т.Е., Ведясова О.А.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Физиология
Статья в выпуске: 1, 2019 года.
Бесплатный доступ
Цель. Сравнительный анализ роли ГАМКА- и ГАМКВ-рецепторов парафациальной респираторной группы (пФРГ) в центральных механизмах регуляции дыхания у взрослых крыс. Материалы и методы. На наркотизированных уретаном крысах исследовали реакции внешнего дыхания и диафрагмальной мышцы на микроинъекции в область пФРГ растворов мусцимола и баклофена (10-6 М, 200 нл). Инъекции осуществляли через стеклянную микроканюлю по стерео-таксическим координатам. Регистрировали спирограмму и электромиограмму (ЭМГ) диафрагмы в исходном состоянии и в течение 60 мин после микроинъекции указанных агонистов ГАМКА- и ГАМКВ-рецепторов. Результаты. Микроинъекции в пФРГ агониста ГАМКА-рецепторов мусцимола оказывали на дыхание тормозное действие, которое развивалось с коротким латентным периодом. У крыс снижалась минутная вентиляция легких за счет изменений дыхательного объема и частоты дыхания, увеличивалось время вдоха и выдоха. На ЭМГ диафрагмы уменьшалась амплитуда осцилляций, увеличивалась длительность инспираторных залпов и межзалповых интервалов...
Парафациальная респираторная группа, гамка-рецепторы, гамкв-рецепторы, мусцимол, баклофен, внешнее дыхание, электромиограмма диафрагмы
Короткий адрес: https://sciup.org/14116360
IDR: 14116360 | УДК: 612.28 | DOI: 10.34014/2227-1848-2019-1-68-75
Respiratory response on the activation of GABAа-and GABAb-receptors of a parafacial respiratory group by specific agonists
The purpose of the paper is to conduct comparative analysis of GABAA and GABAВ receptors of the parafacial respiratory group (pFRG) in central mechanisms of respiratory regulation in adult rats. Materials and Methods. The authors studied the reactions of external respiration and diaphragmatic muscle on microinjections of muscimol and baclofen solutions (10-6 M, 200 nl) in the region of pFRG on rats, anesthetized with urethane. Injections were administered through a glass microcannula according to stereotactic coordinates. A spirogram and electromyogram (EMG) of the diaphragm were recorded in the initial state and within 60 minutes after microinjection of the abovementioned agonists of GABAA and GABAВ receptors. Results. Microinjections of muscimol, the agonist of GABAA receptors, in pFRG had an inhibitory effect on respiration. The inhibitory effect was characterized by a short latent period. Minute lung ventilation decreased in rats due to changes in tidal volume and respiratory rate, while the inspiratory and expiratory time increased...
Текст научной статьи Реакции дыхания на активацию ГАМКа- и ГАМКв-рецепторов парафациальной респираторной группы специфическими агонистами
Введение. В последние годы главным объектом при изучении центральных механизмов регуляции дыхания является ростральная вентролатеральная медуллярная область, в частности парафациальная респираторная группа (пФРГ), нейроны которой обеспечивают различные аспекты деятельности дыхательного центра [1–3]. В пределах пФРГ локализованы преинспираторные ней- роны [4], участвующие в центральной хеморецепции [5] и совместно с нейронами комплекса пре-Бетцингера генерирующие инспираторную активность [6]. Также в пФРГ имеются экспираторные нейроны Е2-типа, обеспечивающие активную экспирацию [7]. Активность дыхательных нейронов пФРГ контролируется тормозными и возбуждающими нейромедиаторами, например ацетил- холином [8], ГАМК [9, 10], глутаматом [11], глицином [12] и нейромодуляторами, такими как гастрин-рилизинг пептид и нейромедин В [13]. Среди перечисленных регуляторов наибольший интерес вызывают тормозные нейроактивные аминокислоты, особенно ГАМК, которая имеется во всех отделах дыхательного центра и принимает участие в формировании ритма и паттерна дыхания. В исследованиях на взрослых животных показано, что на уровне пФРГ влияние ГАМК на дыхание реализуется с участием ГАМКА-рецепторов [9, 12]. Что касается ГАМКВ-рецепторов, то вопрос об их участии в формировании респираторной активности ранее не изучался.
Цель исследования. Сравнительный анализ роли ГАМК А - и ГАМК В -рецепторов парафациальной респираторной группы в центральных механизмах регуляции дыхания у взрослых крыс.
Материалы и методы. Поставлены две серии острых экспериментов на половозрелых беспородных крысах массой 250–300 г, наркотизированных уретаном (1,5 г/кг, внутрибрюшинно) в строгом соответствии с этическими правилами использования лабораторных животных в научно-исследовательских целях. В ходе операции проводили трахеостомию, обнажали крышу черепа и открывали подход к диафрагмальной мышце через разрез на левом боку животного. Затем крыс переносили в стереотаксический прибор СЭЖ-3, фиксировали голову и высверливали трепанационное отверстие в окципитальной кости для доступа к пФРГ. В первой серии опытов (n=8) осуществляли микроинъекции в пФРГ раствора специфического агониста ГАМКА-рецепторов мусцимола, во второй (n=9) – специфического агониста ГАМК B -рецепторов баклофена (Sigma Aldrich). Растворы с концентрацией 10-6 М готовили разведением в искусственной спинномозговой жидкости ex tempore и вводили в пФРГ уни-латерально в объеме 200 нл через стеклянную канюлю с диаметром кончика 20–25 мкм при помощи микрошприца МШ-1. Стереотаксические координаты пФРГ: 11,3 мм каудаль-нее bregma ; 2,3 мм латеральнее срединного шва; 9,4 мм вглубь от дорсальной поверхности мозжечка [9].
Регистрировали внешнее дыхание с помощью электронного спирографа, подсоединенного к трахеотомической трубке, и электромиограмму (ЭМГ) диафрагмальной мышцы, которую отводили стальными биполярными электродами, соединенными с усилителем электромиографа. Выходные сигналы от спирографа и электромиографа подавались на аналогово-цифровой преобразователь, затем на компьютер и записывались в программе PowerGraph 3.2 Professoinal (ООО «Интероптика-С»). Запись велась непрерывно в исходном состоянии и в течение 60 мин после микроинъекции. По спиро-граммам оценивали минутный объем дыхания (МОД, мл-1), дыхательный объем (ДО, мл), длительность вдоха (Ti, с), объемную скорость инспираторного потока (Vi, мл/c), длительность выдоха (Te, с), частоту дыхания (ЧД, мин-1). На ЭМГ диафрагмы определяли длительность инспираторных залпов (ДЗ, с), межзалповые интервалы (МЗИ, с), амплитуду осцилляций (отн. ед.).
Результаты экспериментов обрабатывали в программе SigmaPlot 12.5 (Systat Software Inc.). Использовали Normality Test Shapiro– Wilk и Paired t-test, статистически значимыми считали различия при p<0,05.
Результаты и обсуждение. Основным эффектом микроинъекций в пФРГ крыс раствора селективного агониста ГАМКА-ре-цепторов мусцимола было угнетение легочной вентиляции, на что указывало снижение МОД в течение всего срока наблюдений (табл. 1). Следует отметить, что вклад частотных и объемных параметров спирограммы в данную реакцию был неоднозначным. Характерно, что на 1-10-й мин активации ГАМКА-рецепторов снижение МОД имело наибольшую выраженность (26,0 % от исходного уровня; p<0,01) и было обусловлено существенным уменьшением ДО (на 23,5 %; p<0,01). Однако эффект уменьшения ДО отличался кратковременностью, и в последующие сроки экспозиции изменения МОД начинали определяться преимущественным вкладом ЧД. Так, на 40-й мин действия ГАМКА-агониста МОД уменьшался по сравнению с исходным уровнем на 17,9 % (p<0,05), что было связано с ростом общей продолжительности дыхательного цикла и снижением ЧД на 9,5 % (p<0,05). Обращает на себя внимание тот факт, что на фоне активации ГАМКА-рецепторов в области пФРГ динамика ЧД определялась изменением длительности не только экспираторной, но и инспираторной фазы. На 1-й мин после микро- инъекции мусцимола ЧД незначительно увеличивалась, что обеспечивалось укорочением Ti на 14,9 % (p<0,01), в более поздние сроки действия мусцимола ЧД закономерно уменьшалась за счет пролонгации как Te, так и Ti на 10,2 (p<0,05) и 12,3 % (p<0,01) соответственно.
Таблица 1
Изменение параметров паттерна внешнего дыхания и ЭМГ диафрагмы у крыс после микроинъекций мусцимола (10-6 М) в пФРГ
|
Параметр |
Исх. |
1-я мин |
5-я мин |
10-я мин |
20-я мин |
30-я мин |
40-я мин |
50-я мин |
60-я мин |
|
Ti, c |
0,33± |
0,27± |
0,34± |
0,35± |
0,35± |
0,35± |
0,37± |
0,35± |
0,35± |
|
0,02 |
0,02** |
0,02** |
0,02* |
0,02 |
0,01 |
0,01** |
0,01 |
0,01 |
|
|
Te, с |
0,51± |
0,51± |
0,50± |
0,48± |
0,51± |
0,50± |
0,54± |
0,56± |
0,57± |
|
0,04 |
0,04 |
0,04* |
0,04 |
0,04 |
0,05 |
0,03* |
0,04* |
0,05 |
|
|
Vi, мл/с |
1,90± |
2,02± |
1,34± |
1,28± |
1,49± |
1,40± |
1,53± |
1,75± |
1,74± |
|
0,14 |
0,21 |
0,12* |
0,13** |
0,21* |
0,17* |
0,11 |
0,13 |
0,16 |
|
|
ЧД, мин-1 |
74,01± |
77,81± |
73,13± |
74,35± |
71,50± |
72,54± |
66,97± |
67,27± |
66,10± |
|
5,20 |
4,63* |
3,74 |
4,65 |
4,61 |
5,49 |
3,31* |
3,42* |
3,83* |
|
|
ДО, мл |
0,61± |
0,54± |
0,45± |
0,44± |
0,51± |
0,48± |
0,55± |
0,60± |
0,61± |
|
0,05 |
0,04* |
0,04* |
0,04** |
0,06 |
0,05 |
0,04 |
0,04 |
0,04 |
|
|
МОД, мл-1 |
44,85± |
41,93± |
32,63± |
32,07± |
36,80± |
35,09± |
37,29± |
40,46± |
40,34± |
|
3,92 |
3,38 |
2,14* |
2,84** |
5,99* |
5,00 |
3,41* |
3,34 |
4,39 |
|
|
МЗИ, с |
0,51± |
0,51± |
0,49± |
0,48± |
0,51±0,04 |
0,50± |
0,54± |
0,56± |
0,57± |
|
0,04 |
0,04 |
0,04 |
0,04* |
0,05 |
0,03 |
0,04* |
0,05* |
||
|
ДЗ, с |
0,35± 0,03 |
0,28± 0,02** |
0,37± 0,03 |
0,37± 0,02 |
0,37± 0,03 |
0,38± 0,03 |
0,41± 0,03* |
0,39± 0,03 |
0,38± 0,03 |
|
Амплитуда осцилляций, |
0,21± 0,02 |
0,21± 0,02 |
0,19± 0,02 |
0,19± 0,02* |
0,19± 0,02 |
0,18± 0,02* |
0,18± 0,02 |
0,19± 0,02 |
0,18± 0,02 |
|
отн. ед. |
Примечание. * – p<0,05; ** – p<0,01 (достоверные различия с исходным уровнем).
Важным показателем, подтверждающим вклад ГАМКергических структур пФРГ в деятельность инспираторных механизмов дыхательного центра, является изменение объемной скорости инспираторного потока (Vi). Достаточно быстро, уже на 5-й мин после микроинъекции мусцимола, Vi уменьшалась на 29,3 % (p<0,05) от исходного уровня, и данная тенденция сохранялась до 40-й мин наблюдений (табл. 1, рис. 1а).
Реакциям внешнего дыхания соответствовали изменения ЭМГ диафрагмы (табл. 1, рис. 1б). На первых минутах после микроинъекции мусцимола на ЭМГ отмечалось уменьшение длительности инспираторных залпов и амплитуды их осцилляций в сочетании с укорочением межзалповых интервалов. По мере действия мусцимола частота залпов диафрагмальной активности уменьшалась, а сами залпы и интервалы между ними увеличивались, что является электрофизиологическим коррелятом ослабления ритмики дыхания на фоне активации ГАМКА-рецепторов пФРГ.
i, мл/с
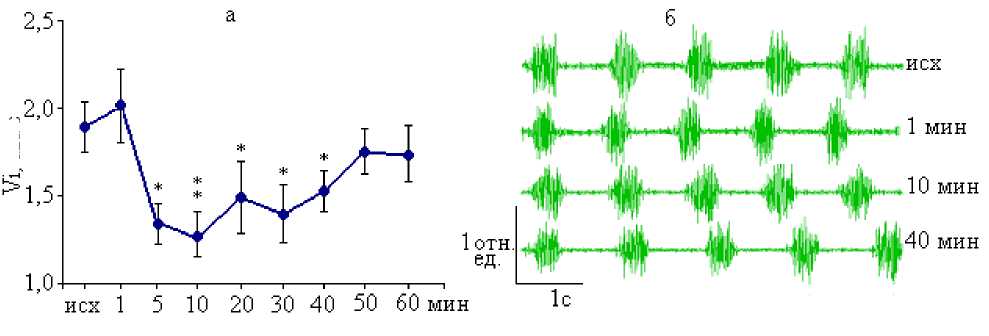
Рис. 1. Респираторные реакции на микроинъекции раствора мусцимола (10-6 М) в пФРГ: а – объемная скорость инспираторного потока; б – суммарная ЭМГ диафрагмы.
Достоверные различия с исходным уровнем при: * – р<0,05; ** – р<0,01
Микроинъекции раствора специфического агониста ГАМК В -рецепторов баклофена в пФРГ крыс также оказывали выраженное угнетающее влияние на легочную вентиляцию (табл. 2). Изменения отдельных параметров паттерна внешнего дыхания, отмеченные в экспериментах с баклофеном, совпадали по направленности с эффектами мусцимола, однако имели место некоторые различия. Во-первых, латентные периоды респираторных эффектов баклофена были более продолжительными, при этом максимальные изменения спирограммы, как и ЭМГ диафрагмы, наблюдались после 30-й мин экспозиции. Одним из объяснений такой растянутой во времени динамики реакций может быть механизм активации метаботропных ГАМКВ-ре-цепторов, включающий в себя сложный каскад обменных процессов и путей передачи сигнала в постсинаптических структурах [15].
Во-вторых, изменения МОД после микроинъекции баклофена в пФРГ, в отличие от мусцимола, в течение всего времени наблюдений определялись отклонением амплитудных параметров спирограммы, а именно ДО, уровень которого закономерно уменьшался (рис. 2а). Так, в конце экспозиции баклофена МОД у крыс снижался на 28,6 % (p<0,01) от исходного уровня (49,12±2,68 мл-1) и составлял 35,08±4,87 мл-1. В это же время достигалось минимальное значение ДО (0,49±0,07 мл), что соответствовало его уменьшению на 24,0 %
(p<0,05) от исходного уровня (0,65±0,05 мл). С этим коррелировало снижение амплитуды осцилляций диафрагмальной активности в среднем на 21,6 % (p<0,05), что хорошо видно на интегрированной ЭМГ (рис. 2б). Что касается временных параметров спирограм-мы, то при активации ГАМК В -рецепторов пФРГ значимо изменялась только величина Ti, которая наиболее выраженно (на 16,8 %; p<0,001) увеличивалась с 30-й по 60-ю мин регистрации. На ЭМГ диафрагмы в эти сроки наблюдалась пролонгация инспираторных залпов на 17,3 % (p<0,05), что совпадало с динамикой Ti. Значения ЧД и Te при воздействии баклофена на пФРГ достоверно не менялись (табл. 2). Величина Vi при этом закономерно уменьшалась в течение всего эксперимента с наибольшим эффектом во второй половине экспозиции. Минимальное значение Vi (1,43±0,24 мл/c; p<0,01) отмечалось на 60-й мин и было на 33,7 % ниже исходного уровня (рис. 2в).
Полученные экспериментальные данные в целом согласуются с современными представлениями о важной роли ГАМКергиче-ских механизмов парафациального региона в регуляции ритма и паттерна дыхания [9, 10]. В настоящей работе выявлено, что в регуляции дыхания нейронами пФРГ участвуют два класса ГАМК-рецепторов – ГАМК А и ГАМК В , которые вносят неоднозначный вклад в формирование респираторных реакций.
Таблица 2
Изменение параметров паттерна внешнего дыхания и ЭМГ диафрагмы у крыс после микроинъекций баклофена (10-6 М) в пФРГ
|
Параметр |
Исх. |
1-я мин |
5-я мин |
10-я мин |
20-я мин |
30-я мин |
40-я мин |
50-я мин |
60-я мин |
|
Ti, c |
0,31± |
0,31± |
0,31± |
0,34± |
0,33± |
0,36± |
0,35± |
0,36± |
0,36± |
|
0,01 |
0,02 |
0,01 |
0,01* |
0,01* |
0,01* |
0,01** |
0,01*** |
0,02** |
|
|
Te, с |
0,49± |
0,49± |
0,44± |
0,47± |
0,46± |
0,48± |
0,50± |
0,49± |
0,50± |
|
0,05 |
0,06 |
0,06 |
0,06 |
0,05 |
0,07 |
0,05 |
0,06 |
0,05 |
|
|
Vi, мл/с |
2,15± |
1,94± |
1,78± |
1,77± |
1,74± |
1,55± |
1,56± |
1,43±0, |
1,43±0, |
|
0,22 |
0,19 |
0,15* |
0,19* |
0,22** |
0,17* |
0,18** |
19** |
23** |
|
|
ЧД, мин-1 |
76,19± 4,33 |
76,94± 5,49 |
81,64± 6,51 |
75,91± 4,68 |
77,47± 5,14 |
73,60± 4,87 |
71,88± 3,74 |
72,37± 4,38 |
71,21± 4,28 |
|
ДО, мл |
0,65± 0,05 |
0,59± 0,04 |
0,56± 0,04** |
0,59± 0,05* |
0,56± 0,06* |
0,55± 0,05* |
0,54± 0,05* |
0,51± 0,06* |
0,50± 0,07* |
|
МОД, мл-1 |
49,12± |
45,50± |
45,35± |
44,89± |
43,32± |
40,24± |
38,54± |
36,40± |
35,08± |
|
2,68 |
3,36* |
3,95 |
4,35 |
4,16** |
4,21* |
3,65** |
3,91** |
4,87** |
|
|
МЗИ, с |
0,31± |
0,48± |
0,45± |
0,47± |
0,46± |
0,48± |
0,49± |
0,49± |
0,50± |
|
0,05 |
0,06 |
0,06 |
0,06 |
0,05 |
0,07 |
0,05 |
0,06 |
0,05 |
|
|
ДЗ, с |
0,49± 0,01 |
0,32± 0,02 |
0,32± 0,01 |
0,34± 0,01** |
0,33± 0,01* |
0,36± 0,01* |
0,35± 0,01** |
0,36± 0,01*** |
0,36± 0,01** |
|
Амплитуда |
0,29± |
0,27± |
0,27± |
0,26± |
0,25± |
0,23± |
0,23± |
0,23± |
0,23± |
|
осцилляций, отн. ед. |
0,03 |
0,03 |
0,03 |
0,03* |
0,02* |
0,02* |
0,02* |
0,02** |
0,02* |
Примечание. * – p<0,05; ** – p<0,01; *** – p<0,001 (достоверные различия с исходным уровнем).
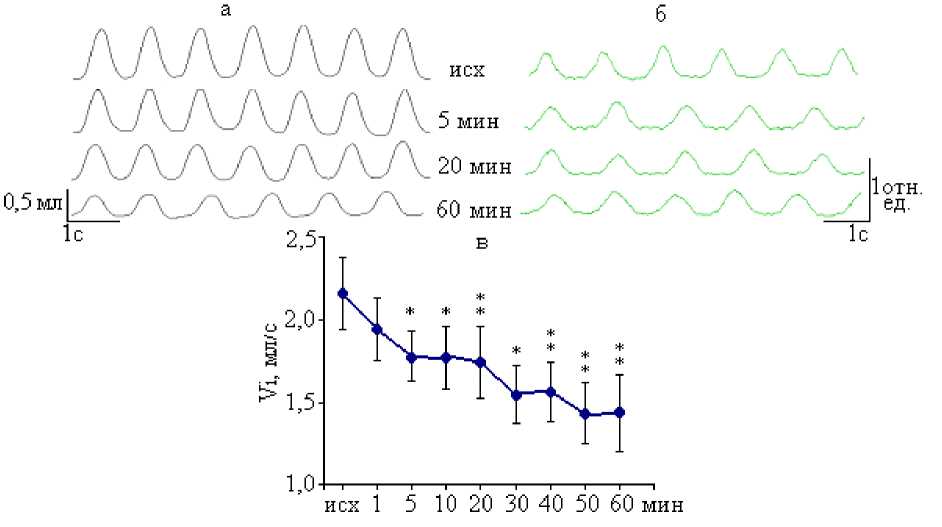
Рис. 2. Респираторные реакции на микроинъекции раствора баклофена (10-6 М) в пФРГ: а – спирограммы; б – интегрированные ЭМГ диафрагмы; в – объемная скорость инспираторного потока. Достоверные различия с исходным уровнем при: * – р<0,05; ** – р<0,01
Во-первых, эффекты активации этих рецепторов различаются по влиянию на дыхательный цикл. При действии агониста ГАМК А -рецепторов мусцимола наблюдалась пролонгация обеих фаз дыхания, тогда как при активации ГАМК В -рецепторов баклофеном удлинялся только вдох. Изменение вдоха в обоих случаях можно объяснить ГАМКер-гическим торможением преинспираторных нейронов пФРГ, которые совместно с аналогичными клетками комплекса пре-Бетцингера составляют основной респираторный ритмо-генератор [2]. Характерное для микроинъекций мусцимола увеличение фазы выдоха, вероятно, обусловлено торможением Е2 экспираторных нейронов, наличие которых в пФРГ описано в ряде работ [7, 11].
Во-вторых, различия в действии агонистов ГАМК А - и ГАМК В -рецепторов касаются скорости формирования респираторных реакций, которая при микроинъекциях мусцимола была больше. Скорее всего это связано с тем, что ГАМК А -рецепторы являются ионотропными и составляют единый комплекс с ионофором, поэтому вызываемое лигандом изменение конформации рецептора ведет к быстрому открыванию каналов для ионов Cl- и моментальной гиперполяризации постсинаптической мембраны [15]. Метаботропные ГАМК B -ре-цепторы имеют иную кинетику мембранных процессов. Их активация ведет к развивающемуся с задержкой слабому, но длительному торможению, тормозной постсинаптический потенциал в этом случае длится несколько сотен миллисекунд [16]. Этот механизм объясняет более позднее развитие ответов системы дыхания на введение в пФРГ баклофена.
При наличии различий в изменениях временных показателей дыхания, микроинъекции ГАМКА- и ГАМКВ-агонистов оказывали сходное влияние на амплитудные параметры спирограммы и ЭМГ диафрагмы. При активации обоих классов ГАМК-рецепторов происходило уменьшение дыхательного объема и амплитуды осцилляций в залпах диафрагмальной активности. Оценивая эти изменения, необходимо напомнить, что во многих публикациях отмечается особая роль пФРГ в регуляции экспираторной фазы дыхательного цикла, в частности в формировании активной экспирации [1, 3]. Что касается участия парафациального региона в регуляции инспирации, то обычно его объясняют активацией хеморецепторов ретротрапециевидно-го ядра [4], нейроны которого переслаиваются с нейронами пФРГ [12]. Наши данные об изменении дыхательного объема и продолжительности вдоха при микроинъекциях агонистов ГАМК в пФРГ свидетельствуют об участии этой области мозгового ствола посредством ГАМКА- и ГАМКВ-рецепторов в модуляции активности механизмов, обеспечивающих процессы формирования не только экспираторного, но и инспираторного моторного выхода. Морфологической основой этих процессов могут быть связи пФРГ с комплексом пре-Бетцингера [17], являющимся основным генератором инспираторного ритма, а также с комплексом Бетцингера [11, 18] и каудальной вентральной респираторной группой [5], которые содержат проприобульбарные и бульбоспинальные экспираторные нейроны.
Заключение. Таким образом, участие пФРГ в регуляции дыхания опосредуется ионотропными ГАМКА- и метаботропными ГАМКВ-рецепторами, которые включены в процессы торможения нейронных структур, контролирующих как временные, так и объемные характеристики паттерна дыхания. При этом ГАМК А -рецепторы в области пФРГ вносят вклад в регуляцию экспирации и инспирации, тогда как ГАМКВ-рецепторы преимущественно участвуют в модуляции инспираторной активности дыхательного центра.
Список литературы Реакции дыхания на активацию ГАМКа- и ГАМКв-рецепторов парафациальной респираторной группы специфическими агонистами
- Feldman J.L., Del Negro C.A., Gray P.A. Understanding the rhythm of breathing: So near, yet so far. Ann. Rev. Physiol. 2013; 75: 423-452.
- Onimaru H., Homma I. A novel functional neuron group for respiratory rhythm generation in the ventral medulla. J. Neurosci. 2003; 23 (4): 1478-1486.
- Pisanski A., Pagliardini S. The parafacial respiratory group and the control of active expiration. Respir. Physiol. Neurobiol. 2018. URL: https://www.ncbi.nlm.nih.gov/pubmed/29933053 (дата обращения: 10.12.2018). DOI: 10.1016/j.resp.2018.06.010
- Abbott S.B., Stornetta R.L., Coates M.B., Guyenet P.G. Phox2b-expressing neurons of the parafacial region regulate breathing rate, inspiration, and expiration in conscious rats. J. Neurosci. 2011; 31 (45): 16410-16422.
- Silva J.N., Tanabe F.M., Moreira T.S., Takakura A.C. Neuroanatomical and physiological evidence that the retrotrapezoid nucleus/parafacial region regulates expiration in adult rats. Respir. Physiol. Neurobiol. 2016; 227: 9-22.
- Onimaru H., Ikeda K., Kawakami K. Phox2b, RTN/pFRG neurons and respiratory rhythmogenesis. Respir. Physiol. Neurobiol. 2009; 168 (1-2): 13-18.
- De Britto A.A., Moraes D.J. Non-chemosensitive parafacial neurons simultaneously regulate active expiration and airway patency under hypercapnia in rats. J. Physiol. 2017; 595 (6): 2043-2064.
- Boutin R.C., Alsahafi Z., Pagliardini S. Cholinergic modulation of the parafacial respiratory group. J. Physiol. 2017; 595 (4): 1377-1392.
- Huckstepp R.T., Cardoza K.P., Henderson L.E., Feldman J.L. Role of parafacial nuclei in control of breathing in adult rats. J. Neurosci. 2015; 35 (3): 1052-1067.
- Vedyasova O.A., Kovaleva T.E. Role of GABAA receptors of parafacial respiratory group in control of respiration in rats. Bull. Exp. Biol. Med. 2018; 165 (6): 711-714.
- Fortuna M.G., West G.H., Stornetta R.L., Guyenet P.G. Betzinger expiratory-augmenting neurons and the parafacial respiratory group. J. Neurosci. 2008; 28 (10): 2506-2515.
- Pagliardini S., Janczewski W.A., Tan W., Dickson C.T., Deisseroth K., Feldman J.L. Active expiration induced by excitation of ventral medulla of adult anesthetized rats. J. Neurosci. 2011; 31 (8): 2895-2905.
- Li P., Janczewski W.A., Yackle K., Kam K., Pagliardini S., Krasnow M.A., Feldman J.L. The peptidergic control circuit for sighing. Nature. 2016; 530 (7590): 293-297.
- Misgeld U., Bijak M., Jarolimek W. A physiological role for GABAB receptors and the effects of baclofen in the mammalian central nervous system. Prog. Neurobiol. 1995; 46 (4): 423-462.
- Belan P.V., Kostyuk P.G. Glutamate-receptor-induced modulation of GABAergic synaptic transmission in the hippocampus. Pflugers Arch. 2002; 444 (1): 26-37.
- Cherubini E., Conti F. Generating diversity at GABAergic synapses. Trends Neurosci. 2001; 24 (3): 155-162.
- Bochorishvili G., Stornetta R.L., Coates M.B., Guyenet P.G. Pre-Betzinger complex receives glutamatergic innervation from galaninergic and other retrotrapezoid nucleus neurons. J. Comp. Neurol. 2012; 520 (5): 1047-1061.
- Smith J.C., Abdala A.P., Rybak I.A., Paton J.F. Structural and functional architecture of respiratory networks in the mammalian brainstem. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009; 364 (1529): 2577-2587.
