Регуляторная роль фотооксидазной реакции в работе циклической редокс-цепи интактных хроматофоров пурпурной бактерии Rhodospirillum rubrum

Автор: Гирва М.В., Белевич И.О., Ременников В.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 2, 2004 года.
Бесплатный доступ
Установлено, что в аэробных условиях в результате оттока электронов на кислород и в анаэробных условиях в присутствии донора электронов наблюдается ингибирование генерации мембранного потенциала, которое устраняется в первом случае при создании анаэробных условий вследствие добавления субстратов окисления фотооксидазной реакции (сукцината), а во втором ~ при возникновении акцепторов электронов, устраняющих чрезмерное восстановление компонентов циклической редокс-цепи (фумарат, О2, ТеОз^")- Полученные данные иллюстрируют регуляторную роль фотооксидазной реакции в генерации мембранного потенциала, обусловленной работой циклической редокс-цепи клеточных хроматофоров К rubrum.
Короткий адрес: https://sciup.org/147204312
IDR: 147204312 | УДК: 579.222.121
The regulative role of the photooxidase reaction of purple popule Rhodospirillum rubrum
It was stated that in aerobic conditions due to care of electrons to oxygen, and in anaerobic conditions in presence of an electron donor generation of membrane potential was repressed. The repression was removed in the fu-st care when anaerobic conditions were created (amber acid) and in the second care when electron acceptor was added (fumarate acid, O2, ТеОз^")- The data obtained illustrate the regulative role of the photooxidase reaction of pmple bacteria in the generation of membrane potential, due to work of the cyclic redox-chain.
Текст научной статьи Регуляторная роль фотооксидазной реакции в работе циклической редокс-цепи интактных хроматофоров пурпурной бактерии Rhodospirillum rubrum
М. В. Гирева, И. О. Белевич, В. Г. Ременников
Пермский государственный университет, 614990, Пермь, ул. Букирева, 15
Установлено, что в аэробных условиях в результате оттока электронов на кислород и в анаэробных условиях в присутствии донора электронов наблюдается ингибирование генерации мембранного потенциала, которое устраняется в первом случае при создании анаэробных условий вследствие добавления субстратов окисления фотооксидазной реакции (сукцината), а во втором - при возникновении акцепторов электронов, устраняющих чрезмерное восстановление компонентов циклической редокс-цепи (фумарат, О2, ТеО32"). Полученные данные иллюстрируют регуляторную роль фотооксидазной реакции в генерации мембранного потенциала, обусловленной работой циклической редокс-цепи клеточных хроматофоров R rubrum.
Ранее нами было показано (Ежова, Мартыненко, Ременников, 2000; Белевич и др., 2001), что при выращивании R. rubrum на среде с ингибирующими концентрациями яблочной и янтарной кислот энергетический кризис проявляется в подавлении роста культуры. Окисление этих сильно восстановленных соединений происходит в результате фотоиндуцированного переноса электронов на различные акцепторы, например, на исследуемые нами О2, Н2О2, (Белевич и др., 2001), ТеО32" (Белевич и др., 2002).
В качестве акцептора электронов при изучении регуляторной роли фотооксидазной реакции интактных хроматофоров пурпурной бактерии R. rubrum использовали оксианионы теллурита.
Объекты и методы исследований
Объектом исследования служила пурпурная несерная бактерия R. rubrum, чистая культура которой была получена из коллекции кафедры микробиологии МГУ. Культивирование бактерий производили в пробирках с притертой пробкой в анаэробных условиях на свету при 35°С, pH = 6,9 на среде с малатом (Bose et al, 1969) в присутствии дрожжевого экстракта.
Поглощение проникающих анионов тетрафенил-бората (ТБ") измеряли с помощью фосфолипидной (азолектиновой) мембраны, разделяющей два отсека тефлоновой кюветы с раствором. Содержание бактериохлорофилла в клетках определяли спектрофотометрически, используя коэффициент экстинкции in vitro, равный 75 мМ"' см"1 при 770 нм.
Среда инкубации содержала 250 мМ сахарозы, 50 мМ трис-НС1-буфер pH 7,6, 1 мкМ тетрафенил-борат натрия и клетки R. rubrum с бактериохлорофиллом - 14 мкг/мл. Некоторые опыты проводили в анаэробных условиях. Для этого в оба отсека кюветы с раствором, дополненным 30 мМ глюкозой (концентрация сахарозы в этих опытах была снижена до 220 мМ), добавляли каталазу и глюкозо-оксидазу по 0,17 мг/мл, 0,1 мМ ТМФД, 5 мМ трис-аскорбат и заливали вазелиновым маслом (толщина слоя 6-8 мм). Раствор в измерительной кювете перемешивали магнитной мешалкой. Добавки: в аэробных условиях - 5 мМ трис-сукцинат; в анаэробных условиях - 5 мМ трис-фумарат, 1 мМ трис-оксалацетат. Аэрацию производили путем удаления слоя вазелинового масла или добавления 0,1 мМ ТеО32".
Для освещения суспензии хроматофоров обычно использовали свет насыщающей интенсивности с длиной волны больше 660 нм.
Результаты и обсуждение
Для изучения регуляторной функции фотооксидазной реакции использовали клетки R. rubrum, инкубируемые в аэробных и анаэробных условиях с различными донорами и акцепторами электронов. Клетки R. rubrum, инкубируемые аэробно, поглощают анионы тетрафенилбората (ТБ"). Этот процесс, протекающий в темноте, обусловлен поглощением данных анионов по градиенту концентрации и их распределением (коэффициент распределения высокий) между клеткой в гидрофоб-
Анаэробиоз Фумарат О2. ТеСК 2"
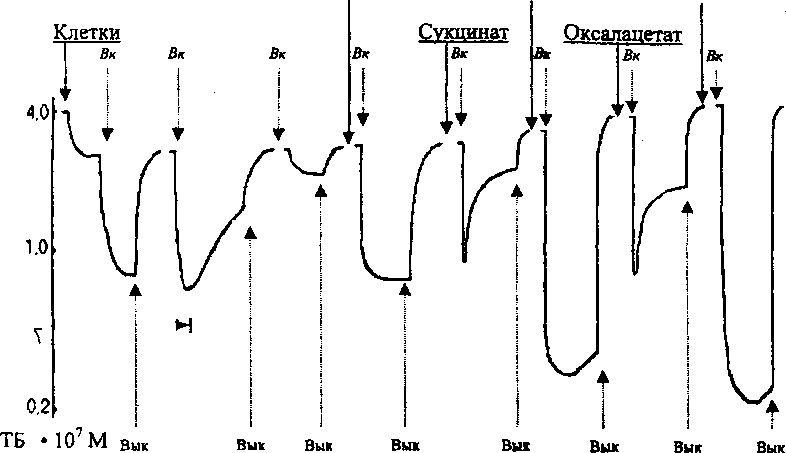
Рис. 1. Фотоиндуцированное поглощение анионов ТБ интактными хроматофорами R. rubrum в аэробных и анаэробных условиях
ной фазе и водной средой. Последующее включение света вызывает дополнительное поглощение анионов клетками против градиента концентрации, что свидетельствует об их переносе из цитоплазмы во внутриклеточные хроматофоры (Ременников, Самуилов, 1979). Снижение концентрации ТБ" в цитоплазме в свою очередь вызывает перенос этих анионов по градиенту концентрации из среды в цитоплазму клетки. Выключение света приводит к восстановлению исходной концентрации ТБ" в среде инкубации.
При длительном освещении клеток, инкубируемых аэробно в отсутствие доноров электронов, наблюдается значительное снижение светового ответа поглощения анионов ТБ". Повторное включение света вызывает дополнительное снижение величины мембранного потенциала, что обусловлено фотооксидазной реакцией, окисляющей компоненты циклической редокс-цепи. Создание анаэробных условий предотвращает спад уровня поглощения ТБ" и восстанавливает его до исходной величины. Данный эффект может быть объяснен восстановлением в анаэробных условиях компонентов циклической цепи за счет эндогенных субстратов. Однако чрезмерное восстановление компонентов циклической редокс-цепи при последующем добавлении в среду инкубации сукцината также вызывает подавление генерации мембранного потенциала (рис. 1). Этот ингибирующий эффект устраняется при добавлении фумарата - акцептора электронов на уровне убихинона. Действие фумарата снимается оксалацетатом - конкурентным ингибитором сукцинатдегидрогеназы. Таким образом, происходит очередное падение уровня поглощения
ТБ", индуцируемое светом.
При добавлении в среду инкубирования другого акцептора электронов - О2 или теллурит-аниона - вызывает максимальное фотоиндуцированное поглощение ионов ТБ".
Полученные данные иллюстрируют регуляторную роль фотооксидазной реакции в генерации мембранного потенциала, обусловленной работой циклической редокс-цепи клеточных хроматофо-ров R rubrum. В аэробных условиях вследствие оттока электронов на кислород и в анаэробных условиях в присутствии донора электронов наблюдается ингибирование генерации мембранного потенциала, которое устраняется в первом случае при создании анаэробных условий в результате добавления субстратов окисления фотооксидазной реакции (сукцината), а во втором - при возникновении акцепторов электронов, которые устраняют чрезмерное восстановление компонентов циклической редокс-цепи (фумарат, О2, ТеО32").
Согласно представлениям Мура и Каплан, при росте пурпурных бактерий в анаэробных условиях на свету и на среде, содержащей сильно восстановленные углеродные соединения, в клетках синтезируется избыточное количество восстановленного НАДН, хинонов и цитохромов, принимающих участие в функционировании циклической цепи. В результате их накопления в клетке нарушается ре-докс-баланс (Moore, Kaplan, 1994), что подавляет работу циклической редокс-цепи как генератора электрохимического протонного градиента. В клетке наступает энергетический кризис, для удаления которого используются механизмы восстановления различных акцепторов электронов (Мо-
Регуляторная роль фотооксидазной реакции в работе циклической редокс-цепи... 109
ore, Kaplan, 1994), среди которых наиболее подходящим по своим физико-химическим характеристикам может быть теллурит. Мнение Мура и Каплан согласуется с результатами наших опытов (рис. 2). Ингибирующее действие на рост культуры R. rubrum оказывает повышение концентрации малата, а также внесение сукцината в среду культивирования в концентрации 15 мМ на фоне 60 мМ малата. Снятие ингибирования происходит при последующем добавлении фумарата - акцептора электронов на уровне убихинонов, но только при повышении концентрации фумарата в среде до 110 мМ. Дальнейшее его увеличение также подавляет рост пурпурных бактерий.
ОП, % Сукцинат

30 60 90 120 150
[Малат], мМ
Рис. 2. Влияние малата, сукцината и фумарата на рост несерной пурпурной бактерии R. rubrum в анаэробных условиях на свету
(100% соответствует 9,3 ед. ОП)
Из этого следует, что вторая фаза ингибирования роста R. rubrum, проявляющаяся при концентрации более 110 мМ малата в среде, обусловлена, вероятно, осмотическим шоком, возникающим при увеличении в среде кислот.
Дальнейшие исследования показали, что ингибирующее действие яблочной кислоты и сукцината снимается при добавлении в среду инкубации теллурита в концентрации от 0,1 мкМ до 100 мкМ (рис. 2). Таким образом, теллурит, выполняя роль акцептора электронов, подобно кислороду, осуществляет регуляцию работы циклической редокс-цепи пурпурных бактерий и тем самым регулирует редокс-баланс клетки.
Следовательно, в меняющихся условиях окружающей среды, фотоиндуцированный нециклический перенос электронов на Ог и Те2О32' является адаптивным механизмом регуляции работы циклической редокс-цепи. Данный процесс можно считать и экологически важным, поскольку идет восстановление теллурита до теллура и осаждение последнего. В этой связи он может быть успешно использован для очистки водоемов от токсичных форм теллура.
Список литературы Регуляторная роль фотооксидазной реакции в работе циклической редокс-цепи интактных хроматофоров пурпурной бактерии Rhodospirillum rubrum
- Белевич И.О., Гирёва М.В., Ременников В.Г., Суворов В.И., Чудинова Л.А. Влияние Ог и Н2О2 на рост пурпурных бактерий на среде с малатом.//Вестник Перм. ун-та. 2001. Вып. 4.Биология. С. 107-120.
- Ежова Л.Е., Мартыненко И.О., Ременников В.Г. Влияние малата на рост пурпурной бактерии Rhodospirillum rubrum. II Вестник Перм. ун-та. 2000. Вып. 2. Биология. С. 143-147.
- Ременников В.Г. Роль кислорода и фотооксидазной реакции в эволюции фотосинтетического аппарата.//Вестник Перм. ун-та. 1997. Вып. 2. Биология. С. 106-110.
- Ременников В.Г, Самуилов В.Д. Нециклический перенос электронов и генерация мембранного потенциала в хроматофррах Rhodospirillum rubrum.//Биол. науки. 1979. № 5. С. 45-52.
- Ременников В.Г., Самуилов В.Д. Генерация мембранного потенциала при функционировании полной и сокращенной систем циклического переноса электронов в хроматофорах Rhodospirillum rubrum. II Биол. науки. 1979. № 10. С. 24-29.
- Moore M.D., Kaplan S. Members of the family Rhodospirillaceae reduce heavy-metal oxyanions to maintain redox poise during photosynthetic growth//ASM News. 1994. Vol. 60, № 1. P. 17-23.
