Ремоделирование костной ткани: возможный вклад адреналина

Автор: Пасатецкая Н.А., Лопатин А.И., Кипенко А.В., Лопатина Е.В.
Журнал: Волгоградский научно-медицинский журнал @bulletin-volgmed
Рубрика: Экспериментальные исследования
Статья в выпуске: 4 (56), 2017 года.
Бесплатный доступ
В последнее время накоплено большое количество данных, подтверждающих участие симпатической нервной системы и ее медиаторов в регуляции процессов ремоделирования костной ткани. Однако результаты фармакологических и генетических экспериментов демонстрируют неоднозначные, порой противоположные, данные. Целью исследования являлся анализ возможного вклада адреналина в ремоделирование костной ткани в условиях органотипического культивирования. В условиях органотипического культивирования нами обнаружен ингибирующий рост эксплантатов ткани кости 10-12-дневных куриных эмбрионов эффект адреналина в дозе 10-4 М. Ингибиторный анализ с помощью кардиоселективного ßi-адреноблокатора атенолола (10-4 М) доказал отсутствие вклада этого типа адренорецепторов в обнаруженный эффект адреналина (10-4 М).
Адреналин, ßi-адренорецепторы, органотипическая культура ткани, эксплантаты костной ткани
Короткий адрес: https://sciup.org/142214556
IDR: 142214556 | УДК: 612.753
The remodeling of bone: possible contribution of adrenaline
More recently, there has been evidence that the sympathetic nervous system and its mediators play a significant role in the regulation of bone remodeling. However, the results of pharmacological and genetic studies demonstrate ambiguous or even conflicting data. The aim of the study was to analyze the potential contribution of adrenaline in bone remodeling under organotypic culture conditions. It was shown that adrenaline at a dose of 10-4 М inhibited the growth of 10-12-day-old chicken embryo bone explants. The inhibitory analysis using a cardioselective ßi-adrenoblocker, atenolol (10-4 M), proved that these adrenergic receptors do not participate in the effect of adrenaline (10-4 M) to inhibit the growth of bone explants.
Текст научной статьи Ремоделирование костной ткани: возможный вклад адреналина
Ремоделирование костной ткани это физиологический процесс, основанный на балансе двух разнонаправленных факторов: формирование новой кости остеобластами и разрушение старой костной ткани остеокластами.
В последнее время накоплено большое количество данных, подтверждающих участие симпатической нервной системы и ее медиаторов в регуляции процессов ремоделирования ткани кости. Гистологические исследования по- казывают, что и кость и надкостница обильно снабжены сенсорными и симпатическими нервными волокнами, максимальная плотность которых регистрируется около ростовой пластинки и метафизах длинных костей. Нервные окончания, содержащие везикулы катехоламинов, прямо контактируют с остеобластами in vivo, что также свидетельствует в пользу наличия нейроэндокринной регуляции ремоделирования ткани кости [8]. Доказано наличие α- и β-адрено- рецепторов на поверхности остеобластов и остеокластов.
Однако, не смотря на большое количество доказательств вовлеченности адренергической системы в процесс регуляции ремоделирования костной ткани, результаты фармакологических и генетических экспериментов демонстрируют неоднозначные, порой противоположные, данные.
ЦЕЛЬ РАБОТЫ
Проанализировать возможный вклад адреналина в ремоделирование костной ткани в условиях органотипического культивирования.
МЕТОДИКА ИССЛЕДОВАНИЯ
Исследования проводили на эксплантатах ткани кости 10–12-дневных куриных эмбрионов. Каждая серия экспериментов включала 120 контрольных эксплантатов и 120 экспериментальных эксплантатов на каждую исследованную концентрацию действующих веществ.
Фрагменты бедренной кости, очищенные от надкостницы, аккуратно переносили на коллагеновую подложку чашек Петри, добавляли питательную среду и культивировали в СО 2 -инкубаторе («Sanyo», Япония) в течение 3 суток при 37 °С и 5 % СО 2 . Препаровку осуществляли инструментами для микрохирургии глаза. Каждая чашка содержала 20 эксплантатов. Контрольные эксплантаты культивировали в питательной среде следующего состава: 40 % раствора Хенкса, 40 % среды Игла, 15 % фетальной телячьей сыворотки. В культуральную среду также добавляли глюкозу (0,6 %), глютамин (2 мМ), гентамицин (100 ЕД/мл) [1]. В питательную среду экспериментальных эксплантатов добавляли адреналин в диапазоне концентраций от 10-4 до 10-14 М, атенолол, в диапазоне концентраций от 10-4 до 10-8 М. В отдельной серии опытов экспериментальные эксплантаты культивировали в питательной среде, содержащей адреналин (10-4 М) и атенолол (10-4 М).
Для визуализации объектов использовали микроскоп «Axiostar Plus» («Carl Zeiss», Германия). Полученные изображения анализировали при помощи программы ImageJ. Для количественной оценки степени роста эксплантатов применяли морфометрический метод. Морфометрический критерий индекс площади (ИП) рассчитывали, как отношение общей площади эксплантата к площади исходной зоны. За условную единицу площади принимали квадрат окуляр-сетки микроскопа, сторона квадрата при увеличении 3,5 х 10 составляла 150 мкм. Значение ИП контрольных эксплантатов принимали за 100 %.
Статистическую обработку результатов проводили с помощью программы STATISTICA 8.0.
При сравнении контрольной и экспериментальной групп использовали критерий Манна-Уитни. Различия считались достоверными при р ˂ 0,05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Влияние адреналина на рост эксплантатов ткани кости 10–12-дневных куриных эмбрионов исследовали в диапазоне концентраций от 10-4 до 10-14 М (рис. 1). В концентрации 10-4 М адреналин угнетал рост эксплантатов ткани кости на 53 % (р ˂ 0,01) по сравнению с контрольным значением. В диапазоне концентраций от 10-6 до 10-14 М адреналин на рост эксплантатов костной ткани практически не влиял. ИП экспериментальных эксплантатов не отличался от контрольного значения.
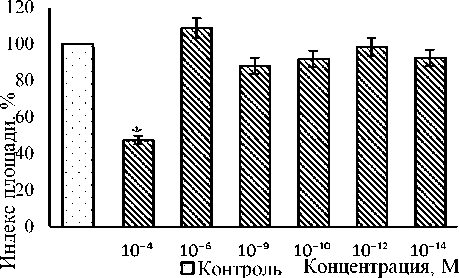
Рис. 1. Оценка участия адреналина в регуляции роста эксплантатов ткани кости 10–12-дневных куриных эмбрионов
* Достоверные различия относительно контроля, p ˂ 0,05.
В следующей серии экспериментов изучали действие селективного β 1 -адреноблокатора атенолола на рост эксплантатов ткани кости в диапазоне концентраций от 10-4 до 10-8 М (рис. 2). Ни в одной из исследуемых концентраций препарат не оказывал влияния на рост эксплантатов ткани кости. ИП экспериментальных эксплантатов не отличался от контрольного значения.
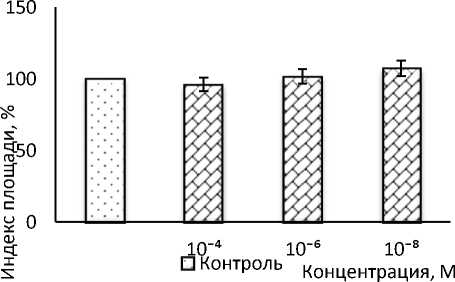
Рис. 2. Влияние атенолола на рост эксплантатов ткани кости 10–12-дневных куриных эмбрионов
* Достоверные различия относительно контроля, p ˂ 0,05.
Для того чтобы проверить возможную связь ингибирующего действия адреналина с влиянием на β 1 -адренорецепторы эксплантаты ткани кости культивировали в питательной среде, содержащей адреналин (10-4 М), в присутствии β 1 -адреноблокатора атенолола (10-4 М) (рис. 3). Наблюдали практически такое же ингибирование роста экспериментальных эксплантатов ткани кости, как при введении одного адреналина. ИП экспериментальных эксплантатов был ниже контрольного значения на 47 % (р ˂ 0,05).
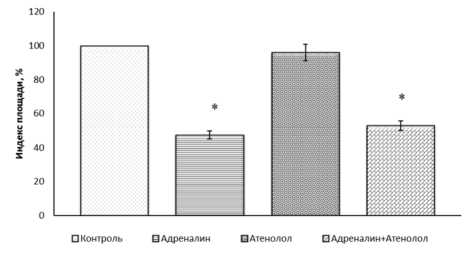
Рис. 3. Атенолол (10-4 М) не устраняет ингибирующее действие адреналина (10-4 М) на рост эксплантатов ткани кости 10–12-дневных куриных эмбрионов
* Достоверные различия относительно контроля, p ˂ 0,05.
Maassen впервые продемонстрировал участие катехоламинов в регуляции роста ткани кости в 1950-х гг. [10]. Позднее было доказано наличие адренорецепторов на поверхности остеокластов и остеобластов. Разные клеточные линии, используемые при исследовании механизмов ремоделирования костной ткани, имеют не одинаковый состав подтипов β -адренорецепторов на поверхности клеток. Экспериментально показано наличие β 2 -адренорецепторов на поверхности клеток линий ROS17/2.8, SaOS-2, HOS, MG-63, и первичной культуры клеток остеобластов мыши. Экспрессия β 2 -адренорецепторов также зарегистрирована в культуре остеокластов [3; 8]. Kellenberger et al. зарегистрировали экспрессию β 1 -адренорецепторов в культуре клеток SaOS-2, TE-85, OHS-4, однако не обнаружили их в культуре первичных остеобластов человека [7]. На основании сравнительного анализа наличия β -адрено-рецепторов в клеточных линиях Kellenberger выдвинул предположение о том, что различные композиции подтипов β -адренорецепторов могут отражать стадии дифференцировки остеобластов в источнике культуры клеток. В недифференцированных остеобластах наблюдается высокий уровень экспрессии β 2 -адренорецепторов и наличие β 1 -адренорецепторов. По мере дифференцировки происходит уменьшение количества β 2 -адрено-рецепторов и исчезновение β 1 -адренорецеп-торов [7].
Полученные in vivo и in vitro экспериментальные данные о механизме действия катехоламинов на ремоделирование ткани кости весьма противоречивы.
Исследования Suzuki позволили зарегистрировать дозозависимое действие адреналина на пролиферацию клеток линии остеобластов мыши MC3T3-E1 в диапазоне концентраций 1 нМ/л – 10 мкМ/л. Максимальный стимулирующий эффект адреналина наблюдался в концентрации 1–10 мкМ/л. Количество клеток увеличилось на (48,4 ± 5,8) % после 48 часов инкубации [5].
Takeuchi в работе на клетках той же линии обнаружил способность адреналина (0,1– 10 мкМ/л) модулировать остеокластогенез. Эффект, по мнению автора, основан на стимуляции адреналином синтеза фактора дифференцировки остеокластов, последнее способствует резорбции костной ткани [2].
В экспериментах in vivo зарегистрировано уменьшение числа остеобластов, скорости формирования кости на трабекулярных поверхностях и уровня экспрессии коллагена I типа при введении изопротеренола или других агонистов адренорецепторов. Напротив, при блокаде адренорецепторов неселективным β -адреноблокатором пропранололом или в исследованиях с использованием генетически модифицированных мышей лишенных β 2 -адренорецепторов наблюдалось увеличение числа остеобластов и массы костной ткани [4].
При изучении мышей лишенных как β 2 -адре-норецепторов, так и β 1 -адренорецепторов, Pierroz обнаружил значимое уменьшение формирования костной ткани (скорость формирования кости снижалась на 79 % по сравнению с мышами дикого типа) [7]. Данные свидетельствуют о том, что, по-видимому, увеличение костной массы при блокаде или отсутствии β 2 -адренорецепторов может наблюдаться лишь при сохранности β 1 -адрено-рецепторов. Хотя точный вклад β 1 -адрено-рецепторов в регуляцию процессов остеогенеза не установлен. Есть лишь предположение, что активация β 1 -адренорецепторов может стимулировать рост костной ткани опосредованно через системные влияния [4].
Преимуществом метода органотипической культуры ткани, который мы применяли в представленной работе, является возможность оценить влияние фармакологических и иных агентов на развитие клеточного сообщества, составляющего определенную ткань, в строго контролируемых условиях. Необходимо отметить, что в органных культурах сохраняется ци-тоархетектоника, присущая исследуемой ткани в условиях in vivo, и связи между клетками, формирующими определенную ткань. При этом исключается не прямое (системное) воздей- ствие со стороны других органов и систем организма на изучаемые процессы.
В нашей работе зарегистрировать стимулирующий рост эксплантатов костной ткани эффект адреналина не удалось. Исследуемые концентрации адреналина от 10-6 до 10-14 М на рост эксплантатов практически не влияли. В концентрации 10-4 М адреналин оказывал выраженное ингибирующее действие. Селективный β 1 -адреноблокатор атенолол ни в одной из исследованных концентраций на рост экспериментальных эксплантатов не влиял.
По-видимому, в этот период онтогенеза (10– 12-й день эмбрионального развития куриного эмбриона) плотность β 1 -адренорецпторов ничтожна. Возможно, особую роль в полученных результатах играет этап эмбриогенеза экспериментального объекта.
Ингибиторный анализ с помощью кардио-селективного β 1 -адреноблокатора атенолола (10-4 М) доказал отсутствие вклада этого типа адренорецепторов в обнаруженный эффект адреналина (10-4 М).
Список литературы Ремоделирование костной ткани: возможный вклад адреналина
- Лопатина, Е. В. Исследование участия Na+, К+-АТФазы в регуляции роста эксплантатов ткани сердца в органотипической культуре/Е. В. Лопатина, В. А. Пеннияйнен, А. А. Зайка//Бюллетень эксперим. биологии и медицины. -2005. -Т. 140. -С. 150-153.
- Adrenergic stimulation of osteoclastogenesis mediated by expression of osteoclast differentiation factor in MC3T3-E1 osteoblast-like cells/T. Takeuchi //Biochem Pharmacol. -2001. -Vol. 61. -P. 579-586.
- Beta2-adrenoreceptor ligands regulate osteoclast differentiation in vitro by direct and indirect mechanisms/S. J. Aitken //Arch Biochem Biophys. -2009. -Vol. 482. -P. 96-103.
- Bonnet, N. Adrenergic control of bone remodeling and its implications for the treatment of osteoporosis/N. Bonnet, D. D. Pierroz, S. L. Ferrari//J. Musculoskelet Neuronal Interact. -2008. -Vol. 8, № 2. -P. 94-104.
- Catecholamines stimulate the proliferation and alkaline phosphatase activity of MC3T3-E1 osteoblast-like cells/A. Suzuki //Bone. -1998. -Vol. 23, № 3. -P. 197-203.
- Evidence for a dense and intimate innervation of the Bone tissue, including glutamate-containing fibers/C. M. Serre //Bone. -1999. -Vol. 25. -P. 623629.
- Formoterol and isoproterenol induce c-fos gene expression in osteoblast-like cells by activating beta2-adrenergic receptors/S. Kellenberger //Bone. -1998. -Vol. 22. -P. 471-478.
- Kondo, H. Beta-adrenergic signaling stimulates osteoclastogenesis via reactive oxygen species/H. Kondo, S. Takeuchi, A. Togari//Am J Physiol Endocrinol Metab. -2013. -Vol. 304. -P. E507-E515.
- Low cortical bone mass in mice lacking beta 1 and beta 2 adrenergic receptors is associated with low bone formation and circulating IGF-1/D. D. Pierroz //J Bone Miner Res. -2006. -Vol. 21. -P. S277.
- Maassen A. P. The influence of adrenalectomy on the growth of rats/A. P. Maassen//Arch Int Pharmacodyn Ther. -1952. -Vol. 88. -P. 473-481.
