Репродуктивная биология живородящей ящерицы Zootoca vivipara (Reptilia, Lacertidae) в Пермском крае
 в Пермском крае")
Автор: Епланова Г.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.13, 2011 года.
Бесплатный доступ
В работе представлены характеристики репродуктивной биологии живородящей ящерицы Zootoca vivipara из популяций Пермского края. У самок живородящей ящерицы длина и масса тела положительно коррелируют с плодовитостью. В процессе яйцеживорождения отмечено длительное (до 2 суток) пребывание новорожденных в яйцевых оболочках. Сравнение полученных данных с опубликованными показало, что плодовитость живородящей ящерицы уменьшается, а размеры новорожденных увеличиваются по направлению к северной границе ареала в европейской части России.
Живородящая ящерица, репродуктивная биология, яйцеживорождение, плодовитость, новорожденный
Короткий адрес: https://sciup.org/148200316
IDR: 148200316 | УДК: 591.526:598.113.6(470.
Reproductive biology of Zootoca vivipara (Reptilia, Lacertidae) in the Perm region
In this article the characteristics of reproductive biology of Zootoca vivipara (Lichtenstein, 1823) from the Perm region are presented. There is a positive significant correlation between body weight with fecundity and body length with fecundity of female of viviparous lizard. The long duration (up to two days) of newborn being in eggshell membranes was observed in process of ovoviparity. Comparison our data and published resulted that the number of newborns decreases and body weight and body size of newborns increases towards the northern border area in the European part of Russia.
Текст научной статьи Репродуктивная биология живородящей ящерицы Zootoca vivipara (Reptilia, Lacertidae) в Пермском крае
Живородящая ящерица Zootoca vivipara (Lichtenstein, 1823) – широко распространенный вид в Северной Евразии [1]. Несмотря на широкое распространение живородящей ящерицы, ее репродуктивная биология остается недостаточно выясненной. Специальных публикаций на эту тему со статистически сравнимыми данными немного [4-6, 8, 11, 15].
Актуальность изучения биологии размножения ящериц из разных регионов обусловлена межпопуляционными различиями репродуктивных характеристик. Например, как показали исследования, проведенные на юго-востоке Западной Сибири, наступление половозрелости самок происходит при достижении определенной длины тела, специфичной для разных популяций [4]. Сроки фенологических событий в жизни ящериц – выхода с зимовки, спаривания, появления новорожденных, продолжительности сезона активности – зависят непосредственно от биотопических и климатических условий конкретных местообитаний и подвержены сезонным колебаниям.
Данная работа является продолжением исследования экологии и биологии живородящей ящерицы на территории Волжского бассейна. Изучение биологии размножения вида на территории Пермского края представляет интерес с точки зрения значительной протяженности данной территории в широтном направлении. Вследствие этого популяции живородящей ящерицы из южной части края расположены ближе к центральной части ареала, а из северной – ближе к его северной периферии.
МАТЕРИАЛ И МЕТОДЫ
Наблюдения за живородящими ящерицами в полевых условиях и их отлов проводился 17-22 мая 2008 г на территории Добрянского и Кунгурского районов и 4-5 июля 2009 г. в Чердынском районе Пермского края.
В лабораторных условиях содержалось 25 беременных самок из следующих пунктов: 1) Добрянский район, биостанция Верх-Кважва, 58°23' с.ш., 56°24' в.д., n =6; 2) Кунгурский район, окрестности с. Плеханово, урочище Спасская гора, 57°28' с.ш, 56°55' в.д. n =3; 3) Чердынский район, окрестности пос. Чепец, 60°43' с.ш., 55°63' в.д., n =16.
С учетом небольшого количества самок из двух первых местообитаний, одинаковых сроков отлова и территориальной приуроченности Добрянского и Кунгурского районов к южной части Пермского края, живородящие ящерицы из этих местообитаний были объединены в «южную» выборку. Ящерицы из Чердынского района, расположенного на севере края, выделены в «северную» выборку.
Самок содержали в террариумах изолировано для наблюдения процесса яйцеживорождения. Родившихся молодых взвешивали и измеряли. В качестве характеристик самок и новорожденных использовались: масса тела W (г), длина туловища с головой L. (мм), длина хвоста L.cd. (мм), общая длина тела L.+L.cd. (мм), индекс L./L.cd . Статистическую обработку проводили общепринятыми методами [8] с использованием программы MS EXEL.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Длина L. размножающихся самок, отловленных в мае 2008 г. в южной части Пермского края, варьировала в пределах 50-65 мм, в среднем составляя 57,4±1,48.
Яйцеживорождение у ящериц южной выборки зафиксировано в период с 25 июня по 3 июля 2008 г. (табл. 1). Наши данные совпадают со сведениями для Северо-Востока европейской России, где самая ранняя откладка яиц была отмечена в конце июня (Печеро-Илычский заповедник, 25 июня 1957 г.) [2]. На основании этого факта было высказано предположение о том, что период эмбрионального развития в данном регионе не такой длинный, как в других частях ареала. Наши наблюдения, полученные в 2008 г. могут служить тому подтверждением.
Таблица 1. Репродуктивные показатели самок живородящей ящерицы из Пермского края
|
№ п/п |
Дата родов |
L ., мм |
Масса после родов, г |
Количество эмбрионов* |
Время выхода новорожденных |
|
1 |
26.06.2008 |
54 |
2,55 |
4 |
– |
|
2 |
25.06.2008 |
60 |
4,24 |
4 (2) |
– |
|
3 |
27.06.2008 |
65 |
4,12 |
6 (1) |
1 ч |
|
4 |
26.06.2008 |
54 |
2,4 |
3 |
– |
|
5 |
29.06.2008 |
50 |
2,35 |
2 |
– |
|
6 |
03.07.2008 |
58 |
2,62 |
1 (4) |
– |
|
7 |
28.06.2008 |
60 |
3,93 |
5 |
– |
|
8 |
29.06.2008 |
56 |
2,925 |
6 |
Обнаружены бегающими |
|
9 |
29.06.2008 |
60 |
4,82 |
9 |
39 ч 30мин |
|
10 |
17.07.2009 |
60 |
4,15 |
6 |
1 ч 30 мин |
|
11 |
18.07.2009 |
55 |
3,18 |
6 |
40 ч |
|
12 |
19.07.2009 |
54 |
2,88 |
4 (3) |
18 ч |
|
13 |
19.07.2009 |
65 |
3,8 |
5 |
Обнаружены бегающими |
|
14 |
20.07.2009 |
61 |
4,1 |
8 |
1 ч 10 мин |
|
15 |
20.07.2009 |
57 |
3,32 |
6 |
36 ч |
|
16 |
20.07.2009 |
58 |
3,24 |
5 |
Обнаружены бегающими |
|
17 |
20.07.2009 |
57 |
2,98 |
2 (3) |
47 ч |
|
18 |
21.07.2009 |
67 |
4,06 |
8 |
7 ч 30 мин |
|
19 |
22.07.2009 |
68 |
4,69 |
9 |
Обнаружены бегающими |
|
20 |
22.07.2009 |
60 |
3,87 |
8 |
8 ч |
|
21 |
22.07.2009 |
63 |
4,04 |
7 |
34 ч |
|
22 |
22.07.2009 |
56 |
2,96 |
5 |
Обнаружены бегающими |
|
23 |
23.07.2009 |
59 |
3,05 |
5 |
25 ч 30 мин |
|
24 |
25.07.2009 |
58 |
3,5 |
6 |
2 |
|
25 |
25.07.2009 |
55 |
3,1 |
4 |
21 ч 30 мин |
|
n |
25 |
25 |
25 |
– |
|
|
min - max |
50-68 |
2,4-4,8 |
1-9 |
– |
|
|
M ± m |
58,8±0,87 |
3,5±0,14 |
5,4±0,42 |
– |
|
Примечание: * – в скобках указано количество жировых яиц в кладке.
Из-за отсутствия наблюдений по срокам выхода и спаривания живородящей ящерицы весной 2008 г., мы не можем точно определить длительность беременности самок. Но, учитывая, что 17-22 мая у всех самок с длиной тела ≥ 50 мм отмечались свежие следы от укусов самцов, можно предположить, что спаривание проходило с начала мая. В этом случае период беременности составляет, вероятно, не более 60 дней.
Определить этот срок по имеющимся в публикациях датам массового выхода ящериц после зимовки и соответствующим им срокам брачного периода не представляется возможным из-за значительного расхождения результатов наблюдений в разные годы. Например, массовый выход в популяции живородящей ящерицы в Пермской области (58°23‘ с.ш.) фиксировался 16.04.1969 г. [3]. В Лысьвенском районе Пермской области массовое появление самцов отмечалось с 6.05.1961 г. и 15.05.1966 г, самки появлялись на 10 дней позднее самцов [11]. На основании этих сведений и наших данных о появлении детенышей, срок беременности живородящей ящерицы в исследуемом регионе очень сильно варьирует (от 31 до 70 дней), что кажется маловероятным.
В целом, по данным разных авторов, продолжительность беременности у живородящей ящерицы в разных регионах России составляет от 46 до 60
дней [6], 60 дней и более [16], 60-70 дней [4], 90 дней [7].
В потомстве ящериц из «южной» выборки Пермского края наблюдалось значительное количество мертвых детенышей – 65% (26 из 40 новорожденных). Только в выводках 3 из 9 самок потомство оказалось жизнеспособным. Эти детеныши ( n =14) появлялись сразу без оболочек или в оболочках, освобождаясь от них в интервале от 1 до 40 часов. Масса тела их составила в среднем 0,19±0,006 г, длина тела 19,1±0,16 мм (табл. 2). 13 новорожденных из кладок разных самок были полностью сформированными, но они либо рождались мертвыми, либо умирали вскоре после родов. Длина их тела достоверно меньше, чем у жизнеспособных детенышей ( L . – 18,0±0,23 мм, t ф = 3,71, р <0,001). Еще 13 детенышей, представляли собой эмбрионы с остатками желточных мешков, покрытые оболочкой. После родов отмечалось их шевеление, но затем они погибли.
В работе Литвинова и Шатненко по Камскому Предуралью [10] приводится информация о 100%-ной гибели отложенных яиц с эмбрионами у живородящих ящериц. К сожалению, возможная причина этого авторами не указана. По другим сведениям для ящериц с юго-востока Западной Сибири, значительная встречаемость в пометах (32,7%) неоп-лодотворенных яиц и недоразвитых эмбрионов в
2004 г была обусловлена аномальным ходом весенних и летних температур [4].
В нашем случае на физиологическое состояние самок неблагоприятное влияние оказала, вероятно, транспортировка и адаптация к условиям в неволе. И в целом, длительное, около месяца, пребывание в лабораторных условиях могло негативно отразиться на формировании эмбрионов.
Плодовитость самок южной популяции составила 4,4 эмбрионов на самку. По полученным данным, масса тела ( t ф =0,77, р >0,05), длина туловища ( t ф =1,17, р >0,05) и плодовитость самок ( t ф =1,68, р >0,05) двух исследуемых популяций не отличаются достоверно, что позволило нам объединить их в одну выборку (см. табл. 1).
Роды в популяции ящериц с севера Пермского края отмечались в период с 17 по 25 июля 2009 г, что на три недели позже, чем у самок из «южной» выборки (см. табл. 1). Вероятно, это можно объяснить более низкими температурами данного сезона наблюдений и смещением сроков яйцеживорожде-ния. Наши наблюдения согласуются с указываемыми для Печеро-Илычского заповедника – там в годы с прохладным летом первых молодых ящериц наблюдали в третьей декаде июля (21-25 июля 1987 г.) [2].
Таким образом, полученные сведения о сроках появления молодых в двух выборках из Пермского края, с учетом их сезонных отличий, совпадают с таковыми, известными для европейского Северо-Востока России [2]. Период яйцеживорождения у ящериц «южной» пермской выборки в 2008 г. (25 июня по 3 июля) совпал также со сроками появления молодых в Среднем Поволжье (с 20 июня по 13 июля 2001, 2003-2007 гг.) [6].
Плодовитость живородящих ящериц из северной части Пермского края составила 5,9 эмбриона на самку. В кладках двух самок отмечено по 3 жировых яйца. В числе 92 новорожденных зафиксировано два погибших детеныша у разных самок:
один родился без яйцевой оболочки уже мертвым и второй умер после того, как освободился от оболочки. Эти детеныши внешне и по параметрам тела не отличались от остальных.
Как показало наблюдение, у самок «северной» популяции ( n =16), потомство 3 ящериц освобождалось от яйцевых оболочек в течение 2 часов, у 9 – процесс выхода занимал от 7 ч 30 мин до 47 ч. У 4 ящериц все детеныши уже бегали к моменту их обнаружения (см. табл. 1). Как правило, такое появление детенышей было приурочено к ночному времени.
Таким образом, в ходе яйцеживорождения выявлено длительное пребывание новорожденных в яйцевых оболочках, отмечавшееся ранее у живородящих ящериц Среднего Поволжья [6]. Косвенным подтверждением существования этого явления в природных условиях может служить защитное поведение, которое наблюдалось у самки из «южной» популяции в процессе откладки яиц. Роды у этой самки длились 40 мин, после чего она оставалась неподвижной еще в течение 35 мин, а затем стала совершать копательные движения мордой и передними лапами, надвигая мох на кладку. Такое поведение, вероятно, является целесообразным, когда имеет место откладка яиц с эмбрионами, которые какое-то время после родов находятся в оболочках, а не сразу освобождаются от них.
Данные наблюдений за процессом яйцеживоро-ждения у ящериц из Пермского Края отличаются от таковых для других частей ареала. По опубликованным сведениям, детеныши живородящей ящерицы покидают яйцевые оболочки в течение более короткого времени. В Ивановской области выход молодых из оболочек фиксировался в интервале от момента родов до 4 часов [8], в Карелии – через несколько минут до получаса [7], на юго-востоке Западной Сибири – тотчас или в течение 1–3 ч [4], в средней и северной тайге Западной Сибири – через 17–40 мин [14].
Таблица 2. Сравнительная таблица параметров тела новорожденных живородящей ящерицы Пермского края
|
Показатель |
Район исследования |
t ф |
p |
|
|
Север Пермского края |
Юг Пермского края |
|||
|
W, г |
92 0,22±0,002 0,17–0,27 |
14 0,19±0,006 0,15-0,22 |
5,80 |
<0,001 |
|
L. , мм |
92 20,8±0,09 19–22 |
14 19,1±0,16 18–20 |
7,35 |
<0,001 |
|
L.cd. , мм |
92 24,4±0,20 20–29 |
14 21,6±0,63 18–25 |
4,94 |
<0,001 |
|
L./L.cd. |
92 0,86±0,006 0,73–1,00 |
14 0,89±0,023 0,79–1,06 |
1,80 |
>0,05 |
Размерные характеристики новорожденных живородящей ящерицы Пермского края приведены в табл. 2. Сопоставление этих данных показало, что потомство самок «северной» выборки крупнее, чем «южной». Детеныши с севера края отличаются на высоком уровне значимости (р<0,001) по массе, длине туловища и длине хвоста. Достоверных отличий не выявлено только по индексу L./L.cd. (р>0,05).
Проводилось сравнение средних значений параметров тела новорожденных из Пермского края с имеющимися литературными данными из ряда регионов (табл. 3) Сравнение популяций из европейской части ареала, в направлении с севера на юг, выявило следующее. Так, новорожденные из Архангельской области отличаются большей длиной туловища ( t ф =3,02, р <0,01) и общей длиной тела ( t ф =2,65, р <0,05) от детенышей с севера Пермского края. Масса тела у них достоверно не отличается ( t ф =0,49, р >0,05). По всем параметрам детеныши из Архангельской области крупнее детенышей из южных популяций Пермского края ( W - t ф =2,54;
р <0,05; L. - t ф =7,35, р <0,001; L.+L.cd - t ф = 3,86, р <0,01). Масса ( t ф =6,94, р <0,001) и общая длина тела новорожденных ( t ф = 5,21, р<0,001) с севера Пермского края достоверно больше, чем у детенышей из Ивановской области. При этом отличий по длине тела не выявлено ( t ф =1,0, р >0,05). У расположенных примерно на одной широте популяций Ивановской области и юга Пермского края, потомство по массе не отличается ( t ф =0,35, р >0,05), но размеры тела достоверно меньше у пермских детенышей ( L. - t ф = 5,85, р <0,001; L.+L.cd - t ф = 2,23, р <0,05).
Таблица 3. Сравнительная таблица параметров тела новорожденных из Пермского края и других регионов
|
Регион |
Показатель ( n , Um , M ± m ) |
||
|
Длина туловища с головой ( L .), мм |
Общая длина тела ( L .+ L . cd .), мм |
Масса ( W), г |
|
|
Север Пермского края (наши данные) |
92 19-22 20,8±0,09 |
92 39-50 45,2±0,24 |
92 0,17-0,27 0,222±0,0020 |
|
Юг Пермского края (наши данные) |
14 18-20 19,1±0,16 |
14 37-45 40,7±0,74 |
14 0,15-0,22 0,190±0,0057 |
|
Архангельская область [8] |
4 21,3-23,7 22,1 ± 0,54 |
4 42,8-52,7 48,6 ± 3,00 |
4 0,200-0,285 0,227 ± 0,0197 |
|
Ивановская область [8] |
22 19-22,2 20,6 ± 0,18 |
22 39,3-46,3 42,4 ± 0,38 |
22 0,167-0,214 0,192 ± 0,0028 |
|
Среднее Поволжье [6] |
114 16-21 19,1 ± 0,10 |
114 34-43 41,5 ± 0,24 |
114 0,14-0,24 0,180 ± 0,0020 |
|
Хабаровский край [8] |
3 21,5-22,5 22,0 ± 0,29 |
3 42,5-44,0 43,2 ± 0,44 |
3 0,171-0,207 0,193 ± 0,0110 |
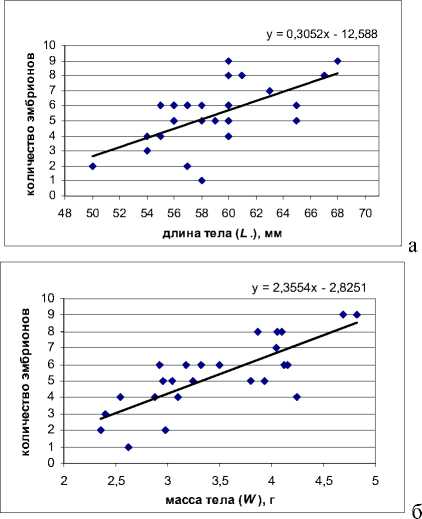
Рис. Корреляция плодовитости с длиной L. (а) и массой W (б) тела у самок живородящей ящерицы из Пермского края
Новорожденные с севера Пермского края в высшей степени достоверно крупнее новорожден- ных из Среднего Поволжья по всем параметрам: массе (tф=14,92, р<0,001), длине тела (tф =12,58, р<0,001) и длине тела с хвостом (tф =10,66, р<0,001). Между потомством «южной» выборки ящериц из Пермского края и Среднего Поволжья достоверных отличий не выявлено.
Сопоставление параметров новорожденных из Пермского и Хабаровского краев показало, что детеныши из юго-востока азиатской части ареала отличаются от детенышей обеих пермских выборок большей длиной туловища («северная» - t ф =2,47, р <0,05; «южная» - t ф =7,63, р <0,001), общая длина тела при этом у них не отличается («северная» - t ф =1,45, р >0,05; «южная» - t ф = 1,51, р >0,05). Масса потомства из Хабаровского края меньше, чем у детенышей с севера Пермского края ( t ф =2,57, р <0,05) при отсутствии различий с массой потомства из «южной выборки» ( t ф =0,22, р >0,05).
Таким образом, при сравнении потомства живородящей ящерицы из европейской части ареала прослеживается тенденция уменьшения размеров тела и массы новорожденных по направлению к южной границе ареала. Детеныши ящериц с севера европейского ареала (северная выборка из Пермского края) отличаются от новорожденных юго- востока азиатской части ареала (Хабаровский край) пропорциями тела и меньшей массой.
Средняя плодовитость живородящих ящериц из Пермского края составила 5,4 эмбриона на самку (см. табл. 1). Выявлена положительная достоверная корреляция количества эмбрионов с массой ( r =0,784, t ф =6,05, р <0,001) и длиной тела самок ( r =0,629, t ф =3,88, р <0,001) (рисунок).
По имеющимся сведениям, прямая положительная зависимость плодовитости с длиной и массой тела ящериц зафиксирована для некоторых районов Ивановской области ( r =0,6) [8]. Но, к сожалению, из текста не ясно, достоверен ли коэффициент корреляции. Положительная достоверная корреляция между плодовитостью и размерами тела самок выявлена в азиатской части ареала для объединенной выборки из Томской области ( r =0,68, р <0,001) [15] и для популяций с юго-востока Западной Сибири ( r=0,72, p =0) [4]. Зависимость числа развивающихся яиц от размера самки ( r =0,507) отмечена для ящериц из Костромской области [12].
Сравнение плодовитости ящериц из Пермского края с опубликованными данными для других регионов европейской части России показало следующее (табл. 4). У ящериц из популяций, расположенных севернее или приблизительно на той же широте (европейский Северо-Восток России, Архангельская, Новгородская, Пермская, Костромская, Ивановская области), что и обследуемый регион, плодовитость достоверно не отличается. Большей плодовитостью отличаются живородящие ящерицы из Среднего Поволжья ( t ф =3,12, р <0,01), обитающие вблизи южной границы ареала [6]. Между плодовитостью ящериц из Пермского края и Саратовской области, находящейся на юге ареала, отличий не выявлено.
Как видно из табл. 4, большинство популяций из азиатской части ареала по плодовитости не отличается (№ 10, 11, 12, 16, 17, 19, 20, 21, 22) или превышает (№ 13, 14, 18) плодовитость ящериц из Пермского края. Только в двух азиатских популяциях этот показатель достоверно ниже (№ 15, 23).
Таблица 4. Сравнительная таблица плодовитости живородящей ящерицы из Пермского края и других регионов европейской части России
|
№ п/п |
Регион |
Плодовитость |
t ф |
p |
||
|
n |
M ± m |
lim |
||||
|
1 |
Пермский Край (наши данные) |
25 |
5,4±0,42 |
1-9 |
- |
- |
|
2 |
Европейский Северо-Восток России, средняя тайга [2] |
18 |
6,4±0,43 |
3-10 |
1,62 |
>0,05 |
|
3 |
Архангельская область [8] |
6 |
5,5±0,67 |
4-8 |
0,11 |
>0,05 |
|
4 |
Новгородская область [11] |
8 |
4,9±0,52 |
4-8 |
0,62 |
>0,05 |
|
5 |
Пермская область [11] |
22 |
5,4±0,31 |
3-9 |
0 |
>0,05 |
|
6 |
Костромская область [12] |
26 |
5,8±0,37 |
3-9 |
0,71 |
>0,05 |
|
7 |
Ивановская область [8] |
6 |
6,5±0,84 |
4-9 |
1,16 |
>0,05 |
|
8 |
Среднее Поволжье [6] |
19 |
7,8±0,69 |
3-14 |
3,12 |
<0,01 |
|
9 |
Нижнее Поволжье, Саратовская область [16] |
17 |
6,2±0,41 |
3-11 |
1,31 |
>0,05 |
|
10 |
Южный Урал [11] |
8 |
6,6±0,68 |
5-9 |
1,43 |
>0,05 |
|
11 |
Южный Алтай, Маркагольская котловина, с. Успенка, 1500-1550 м н.у.м. [11] |
28 |
4,7±0,21 |
2-7 |
1,54 |
>0,05 |
|
12 |
Южный Алтай, Маркагольская котловина, с. Урунхайка, 1500-1600 м н.у.м. [11] |
11 |
5,2±0,40 |
4-8 |
0,29 |
>0,05 |
|
13 |
Южный Алтай, предгорья Курчумского хребта, до 800 м н.у.м. [11] |
5 |
8,2±0,38 |
7-9 |
2,90 |
<0,01 |
|
14 |
Северный Алтай, 900-1100 м н.у.м. [11] |
9 |
7,7±1,00 |
3-13 |
2,51 |
<0,05 |
|
15 |
Северо-Восточный Алтай, Прителецкий р-н, Артыбаш, 450-500 м н.у.м. [11] |
7 |
3,6±0,30 |
3-5 |
2,20 |
<0,05 |
|
16 |
Северо-Восточный Алтай, Кебезень, 450-500 м н.у.м. [11] |
4 |
5,8±0,63 |
4-7 |
0,36 |
>0,05 |
|
17 |
Кузнецкий Алатау, Гавриловка, 550-600 м н.у.м. [11] |
24 |
6,0±0,32 |
3-10 |
1,12 |
>0,05 |
|
18 |
Кузнецкий Алатау, оз. Чудное, 650-600 м н.у.м. [11] |
11 |
8,6±0,31 |
7-10 |
4,78 |
<0,001 |
|
19 |
Томская обл., Причулымье, Тегульдет, 350 м н.у.м. [11] |
7 |
5,1±0,99 |
2-10 |
0,31 |
>0,05 |
|
20 |
Объединенная выборка, окрестности г. Томска [11] |
106 |
6,3±0,21 |
5,6-7,1 |
1,88 |
>0,05 |
|
21 |
Западная Сибирь, северная тайга [11] |
18 |
6,1±0,34 |
4-9 |
1,22 |
>0,05 |
|
22 |
Западная Сибирь, средняя тайга [11] |
15 |
6,5±0,61 |
4-11 |
1,53 |
>0,05 |
|
23 |
Хабаровский край [8] |
4 |
2,25±0,47 |
1-3 |
2,91 |
<0,01 |
Таким образом, сопоставление полученных и опубликованных данных о плодовитости живородящей ящерицы выявило ее уменьшение в направлении к северной периферии ареала европейской части России. Это факт, вероятно, может служить подтверждением литературных данных об уменьшении количества детенышей в северных частях ареала вследствие влияния климатических условий [13].
Сравнение еще одной репродуктивной характеристики - длины тела ( L .) самок живородящей ящерицы, участвующих в размножении - по нашим и литературным данным [6, 8, 11], не выявило каких-либо закономерностей. Она ненаправленно варьирует в пределах европейской части ареала.
Наши данные о длине туловища размножающихся самок Пермского края не отличаются от опубликованных сведений для областей, лежащих приблизительно на одной с ним широте (Новгородская, Пермская, Ивановская [8, 11]). Но, наряду с этим, длина тела самок живородящей ящерицы достоверно больше в регионах, расположенных севернее (Архангельская область t ф =3,83, р <0,001) [8] и южнее (Среднее Поволжье t ф =2,37, р <0,05) [6].
ВЫВОДЫ
-
1. Плодовитость живородящей ящерицы из Пермского края статистически значимо ниже таковой из популяций, расположенных вблизи южной границы ареала в Поволжье.
-
2. Новорожденные живородящей ящерицы с северной части ареала («северная» выборка из Пермского края) отличаются большей массой и размерами тела от детенышей из популяций, расположенных южнее («южная» выборка из Пермского края, Среднее Поволжье).
-
3. Плодовитость самок живородящей ящерицы в выборке из Пермского края положительно коррелирует с массой и размерами тела.
-
4. Отмечено, что у 10 из 25 самок потомство освобождается от яйцевых оболочек не в первые часы после откладки яиц, а позже, в интервале от 7 ч 30 мин до 47 ч.
Автор выражает признательность сотрудникам Пермского государственного педагогического университета к.б.н. Н.А. Литвинову, к.б.н. Н.А. Четанову и сотруднику Национального парка «Самарская Лука» А.А. Поклонцевой за помощь в сборе материала и сотруднику ИЭВБ РАН к.б.н. А.Г. Бакиеву за ценные замечания и помощь в подготовке статьи.
Список литературы Репродуктивная биология живородящей ящерицы Zootoca vivipara (Reptilia, Lacertidae) в Пермском крае
- Ананьева Н.Б., Орлов Н.Л., Халиков Р.Г. и др. Атлас пресмыкающихся Северной Евразии (таксономическое разнообразие, географическое распространение и природоохранный статус). СПб., 2004. 232 с.
- Ануфриев В.М., Бобрецов А.В. Амфибии и рептилии. СПб.: Наука, 1996. 130 с.
- Болотников А.М., Хазиева С.М., Литвинов Н.А., Чащин С.П. Распространение и сезонная активность амфибий и рептилий Пермской области//Вопросы герпетологии. Л.: Наука, 1973. С. 40-41.
- Булахова Н.А. Ящерицы (Reptilia, Squamata, Lacertidae) юго-востока Западной Сибири (география, экология, морфология): Автореф. дис. … канд. биол. наук. Томск, 2004. 26 с.
- Епланова Г.В. Особенности репродуктивной биологии живородящей ящерицы Zootoca vivipara в Среднем Поволжье.//Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Вып. 8. Тольятти, 2005. С. 25-29.
- Епланова Г.В. К репродуктивной биологии живородящей ящерицы Zootoca vivipara (Reptilia, Lacertidae) в Среднем Поволжье//Известия Самарского НЦ РАН, 2009. Т.11, № 1 (27). С. 83-88.
- Ивантер Э.В., Коросов А.В. Земноводные и пресмыкающиеся. Петрозаводск: Изд-во Петрозавод. ун-та, 2002. 168 с.
- Лазарева О.Г. Репродуктивная биология живородящей ящерицы Lacerta vivipara (Sauria, Lacertidae) в заповедниках лесной зоны России//Вторая конференция герпетологов Поволжья: Тез. докл. Тольятти, 1999. С. 27-29.
- Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990. 352 с.
- Литвинов Н.А., Шатненко Т.М. К экологии живородящей ящерицы в Камском Предуралье//Вопросы герпе-тологии. Л.: Наука, 1977. С. 134.
- Орлова В.Ф., Куранова В.Н., Булахова Н.А. Размножение живородящей ящерицы Zootoca vivipara (Jacquin, 1787) в восточной части ареала//Вестн. Томск. ун-та. Сер. «Биол. науки». Прил. № 8. Материалы науч. конф., симпоз., школ, проводимых в ТГУ. Томск, 2003. С. 150-158.
- Окулова Н.М., Колесова Т.М. Материалы Ю.Ф. Сапоженкова о живородящей ящерице Lacerta vivipara Jacquin, 1787 Костромской области.//Поволж. экол. журн. 2002. № 3. С. 236-245.
- Терентьев П.В., Чернов С.А. Определитель земноводных и пресмыкающихся. М.: Сов. наука, 1949. 340 с.
- Шамгунова Р.Р. Эколого-географическая характеристика рептилий средней и северной тайги Западной Сибири: Автореф. дис. … канд. биол. наук. Екатеринбург, 2010. 20 с.
- Orlova V.F., Kuranova V.N., Bulakhova N.A. Some aspects of reproductive biology of Zootoca vivipava (Jacquin, 1787) in the Asian part of its area//Herpetologia Petropolitana: Proc. of the 12th Ord. Gen. Meet. of SEH. SPb., 2005. P. 201-204.
- Tabachisin V., Zavialov E., Tabachisina I. Distribution and ecology of Zootoca vivipara in the north of the Volga region//Common Lizard: Abstr. and Progr. Intrern. Symp. Bonn, 2006.
