Респираторные эффекты активации паратригеминальной области у крыс

Автор: Ведясова О.А., Ковалева Т.Е.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Физиология
Статья в выпуске: 4, 2017 года.
Бесплатный доступ
Актуальным аспектом проблемы регуляции дыхания у млекопитающих животных является изучение механизмов, опосредующих респираторную активность паратригеминальной области (ПТО) моста. Цель. Анализ изменений внешнего дыхания и реакций диафрагмальной мышцы при электростимуляции и активации L-глутаматом ПТО у крыс. Материалы и методы. Поставлены острые опыты на крысах, наркотизированных уретаном. Электростимуляцию ПТО осуществляли монополярно импульсным током посредством стального игольчатого микроэлектрода. Раствор L-глутамата (10-7 М; 0,2 мкл) инъецировали в ПТО через стеклянную микроканюлю при помощи микрошприца МШ-1. Внешнее дыхание регистрировали методом спирографии, реакции диафрагмы оценивали по электромиограмме (ЭМГ). Результаты. Активация ПТО путем электростимуляции и микроинъекций L-глутамата вызывала сходные эффекты в виде ослабления легочной вентиляции и уменьшения частоты дыхания за счет пролонгации экспираторной фазы. Изменениям паттерна дыхания соответствовало увеличение интервалов между инспираторными залпами на ЭМГ диафрагмы. Заключение. ПТО у млекопитающих животных участвует в регуляции ритма и паттерна дыхания. Реализация респираторных эффектов ПТО опосредуется глутаматергическими связями, которые у крыс преимущественно включены в механизмы, определяющие длительность фазы выдоха и, таким образом, частоту дыхания.
Паратригеминальная область, глутамат, паттерн внешнего дыхания, электромиограмма диафрагмы
Короткий адрес: https://sciup.org/14113313
IDR: 14113313 | УДК: 612.28 | DOI: 10.23648/UMBJ.2017.28.8756
Respiratory effects of the paratrigeminal area activation in rats
A very important aspect of breathing control problem in mammals is studying mechanisms, which mediate respiratory activity of the paratrigeminal area (PA) of the pons. Objective. The purpose of the study is to analyze the changes in external respiration and diaphragm reactions to electrical stimulation and L-glutamate activation into PA in rats. Materials and Methods. Acute experiments were performed on rats anesthetized with urethane (n=15). PA was irritated with a pulse current by means of unipolar steel needle-shaped microelectrode. L-glutamate solution (10-7 M, 0.2 μl) was injected into PA through a glass micro-cannula (microsyringe MSh-1). External respiration was registered by means of spirography; diaphragm response was estimated using electromyogram (EMG). Results. PA activation by electrical stimulation and L-glutamate microinjection caused similar effects in the form of pulmonary ventilation reduction and respiratory rate reduction due to prolongation of the expiratory phase. Changes in the breathing pattern corresponded to increased intervals between the inspiratory volleys on the EMG of the diaphragm. Conclusion. The data obtained indicate that PA plays an important part in the rhythm and breathing pattern regulation of mammals. PA involvement in respiratory control is mediated by glutamatergic bonds. In rats, such bonds are included in the mechanisms regulating the duration of the exhalation and, thus, respiratory rate.
Текст научной статьи Респираторные эффекты активации паратригеминальной области у крыс
Введение. Центральные механизмы регуляции дыхания составляют важную медико-биологическую проблему, при решении которой внимание исследователей долгие годы было сосредоточено на ритмогенерирующей функции бульбарного дыхательного центра [1–3]. Нейроны, участвующие в рит-могенезе дыхания, также обнаружены в варолиевом мосту, в понтинной дыхательной группе, куда входят регион «парафациальная респираторная группа/ретротрапециевидное ядро» [4, 5] и паратригеминальная область (ПТО), респираторная активность которой впервые была выявлена у миноги [6]. Нейроны ПТО, названные паратригеминальной респираторной группой (ПТРГ), исследователи рассматривают как генератор дыхательного ритма у низших позвоночных [6, 7]. У крыс вблизи ПТО (в интер-, супра- и сенсор- ном тригеминальных ядрах) выявлены дыхательные нейроны с фазовопереходным паттерном разрядов [8], что позволяет предполагать участие ПТО в регуляции респираторного ритма и у млекопитающих животных.
Актуальным является изучение нейрохимической организации понтинной дыхательной группы [9], при этом закономерный интерес вызывает глутамат как основной возбуждающий медиатор, участвующий в регуляции дыхания [10]. Роль глутамата на уровне ритмогенерирующих нейронов ПТО в настоящее время интенсивно изучается у низших позвоночных [7, 11]. Вопрос о значении глутаматергической нейропередачи в формировании респираторной активности ПТО и в целом об участии ПТО в регуляции дыхания у млекопитающих животных остается открытым.
Цель исследования. Анализ изменений паттерна внешнего дыхания и реакций диафрагмальной мышцы при электростимуляции и активации глутаматом ПТО у крыс.
Материалы и методы. Поставлены две серии острых опытов на беспородных половозрелых крысах обоего пола массой 250–300 г, наркотизированных уретаном (1,5 г/кг, внутрибрюшинно). В первой серии (n=6) изучали респираторные реакции на электростимуляцию ПТО, во второй (n=7) – на микроинъекции в ПТО L-глутамата. Все этапы исследования выполнены с соблюдением правил биоэтики и одобрены комиссией по биоэтике Самарского университета. В ходе операции проводили трахеостомию, открывали подход к диафрагме через кожный разрез на левом боку, обнажали крышу черепа и высверливали трепанационное отверстие в париетальной кости. Оперированных крыс закрепляли в стереотаксическом приборе СЭЖ-3 спиной вверх. ПТО раздражали импульсным током от универсального электростимулятора посредством монополярного игольчатого микроэлектрода с диаметром кончика 60 мкм. Индифферентный электрод закрепляли на мышцах шеи. Параметры тока: частота 5 Гц; напряжение 3 и 7 В; длительность импульса 1 мс, время раздражения 3 с. Для микроинъекций в ПТО использовали раствор L-глутамата (Sigma Aldrich) концентрацией 10-7 М, который готовили ex tempore на основе искусственной цереброспинальной жидкости. Раствор инъецировали в объеме 0,2 мкл через стеклянную канюлю (диаметр кончика 20–25 мкм), укрепленную на игле микрошприца МШ-1. Электростимуляцию и микроинъекции осуществляли по координатам стереотаксического атласа мозга крысы [12]: 12,72 мм каудальнее bregma; 2,8 мм ла-теральнее срединного шва; 7,6 мм вглубь от поверхности черепа.
Внешнее дыхание регистрировали через трахеостомическую трубку при помощи электронного спирографа. Электромиограмму (ЭМГ) диафрагмальной мышцы отводили стальными биполярными электродами (межэлектродное расстояние 3 мм), подсоединенными к миографическому усилителю. Выходные сигналы от спирографа и электро- миографа через аналогово-цифровой преобразователь подавались на компьютер и записывались в программе PowerGraph 3.2 Profes-soinal (ООО «Интероптика-С»). Респираторные эффекты регистрировали в исходном состоянии, при электростимуляции ПТО и в течение 60 мин после микроинъекции глутамата. По спирограммам определяли длительность вдоха (Ti, c) и выдоха (Te, с), частоту дыхания (ЧД, цикл/мин), дыхательный объем (ДО, мл), минутный объем дыхания (МОД, мл/мин). На ЭМГ оценивали длительность инспираторных залпов (ДЗ, с), межзалповые интервалы (МЗИ, с), максимальную амплитуду осцилляций (отн. ед.).
Для статистической обработки применяли парный t-test из пакета программ SigmaStat 4.0 (Jandel Scientific, USA). Все данные приведены как средние значения и ошибка изменения в % относительно исходного уровня. Достоверными считали различия при p<0,05.
Результаты и обсуждение. Характерным респираторным эффектом активации ПТО путем электростимуляции и микроинъекций L-глутамата было угнетение дыхания, обусловленное изменениями частотных показателей спирограммы и ЭМГ диафрагмы. Интегральный эффект раздражения ПТО проявлялся уменьшением МОД на 5,4–9,6 % (p<0,05) от исходного уровня в зависимости от силы раздражения. Ослабление внешнего дыхания обусловливалось снижением ЧД на 6,6–7,9 % (p<0,05), тогда как ДО практически не менялся. Изменения ЧД определяя-лись преобразованиями фазовой структуры дыхательного цикла за счет увеличения Te от 0,512±0,03 (исходный уровень) до 0,683±0,04 c (при стимуляции ПТО). По мере усиления раздражения эффект пролонгации выдоха возрастал и мог достигать 33,4 % (p<0,05) от исходного значения. Изменениям Te и ЧД соответствовали увеличение МЗИ на 17,9 % (p<0,05) и уменьшение частоты инспираторных залпов на ЭМГ диафрагмы (рис. 1). Значимых изменений Ti на спиро-граммах, как и ДЗ и амплитуды осцилляций на ЭМГ диафрагмы, при электростимуляции ПТО у крыс не наблюдалось.
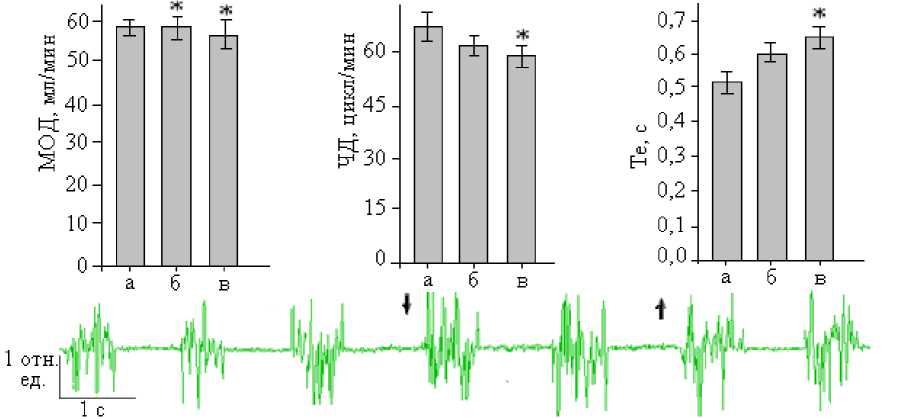
Рис. 1. Респираторные эффекты электростимуляции ПТО у крыс: вверху - изменение параметров внешнего дыхания
(а - исходное состояние; б, в - стимуляция током 5 Гц, 3 В и 5 Гц, 7 В соответственно), внизу - ЭМГ диафрагмы (начало и окончание раздражения отмечено стрелками).
* - достоверные различия с исходным уровнем (p<0,05)
Микроинъекции L-глутамата в ПТО крыс вызывали респираторные реакции, которые также проявлялись преимущественными изменениями временных параметров дыхания по тормозному типу. Результаты активации ПТО L-глутаматом подтвердили отмеченную в опытах с электростимуляцией тесную связь изучаемой области с экспираторными механизмами. Изменения спирограмм и ЭМГ после микроинъекции медиатора имели длительный латентный период, что можно объяснить участием глутаматцептивных элементов ПТО в регуляции ритмики дыхания посредством сложных межсинаптических взаимодействий.
Наиболее выраженные реакции дыхания в виде увеличения Те и синхронного снижения ЧД начинались с 30-й мин от момента введения медиатора и достигали максимальных проявлений после 40-й мин. Значения Те при действии L-глутамата на ПТО возрастали в диапазоне от 0,508±0,030 (исходное состояние) до 0,673±0,040 с на 60-й мин экспозиции (рис. 2а), что составляло 32,3 % (p<0,05). Эти изменения совпадали с динамикой ЧД, которая закономерно снижалась в течение всего периода регистрации. Наибольшее влияние L-глутамата отмечалось на 40-й мин экспози ции, когда ЧД уменьшалась на 22,9 % (p<0,05): от 66,88±3,82 до 51,56±3,08 цикл/мин (рис. 2б). Статистически значимых изменений ДО при этом не выявлено. Динамика временных параметров ЭМГ диафрагмы совпадала с отмеченными изменениями паттерна дыхания. Типичной реакцией диафрагмы на инъекции L-глутамата в ПТО было выраженное удлинение МЗИ на 27,9 и 38,9 % (p<0,001) на 50-й и 60-й мин экспозиции (рис. 2в) в сочетании с небольшим приростом ДЗ (10,7 %; p<0,05). То есть доминировали изменения МЗИ, которые являются электрофизиологическим коррелятом экспирации.
Величина ДО на фоне активации глутаматных рецепторов ПТО оставалась практически неизменной на протяжении всего времени наблюдений. Поэтому направленность динамики легочной вентиляции в сторону снижения была связана прежде всего с уменьшением ЧД. Как видно из гистограмм на рис. 2г, тенденция снижения МОД от исходного уровня (58,73±1,99 мл/мин) приобретала достоверный характер начиная с 40-й мин (уменьшение до 52,744±2,76 мл/мин, или на 10,2 %) и усиливалась к 60-й мин воздействия L-глутамата (уменьшение до 50,09± ±2,69 мл/мин, или на 14,7 %).
компонента внешнего дыхания и инспира-
мышцы торной активности диафрагмальной (рис. 3).
В качестве иллюстрации описанных реакций представлены спирограммы и ЭМГ, из которых видно, что активация ПТО L-глута-
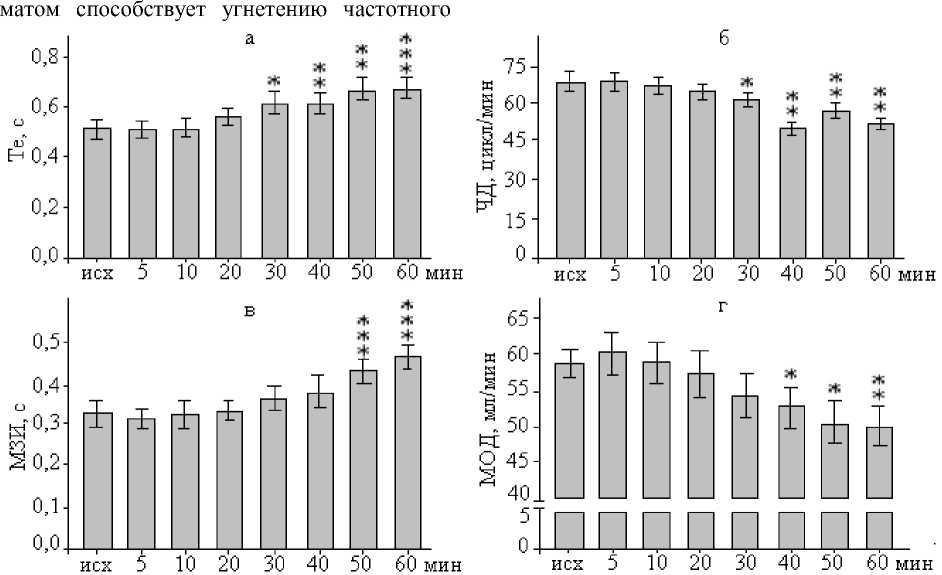
Рис. 2. Изменение параметров внешнего дыхания и ЭМГ диафрагмы после микроинъекции L-глутамата в ПТО у крыс.
Достоверные различия с исходным уровнем при: * - p<0,05; ** - p<0,01; *** - p<0,001
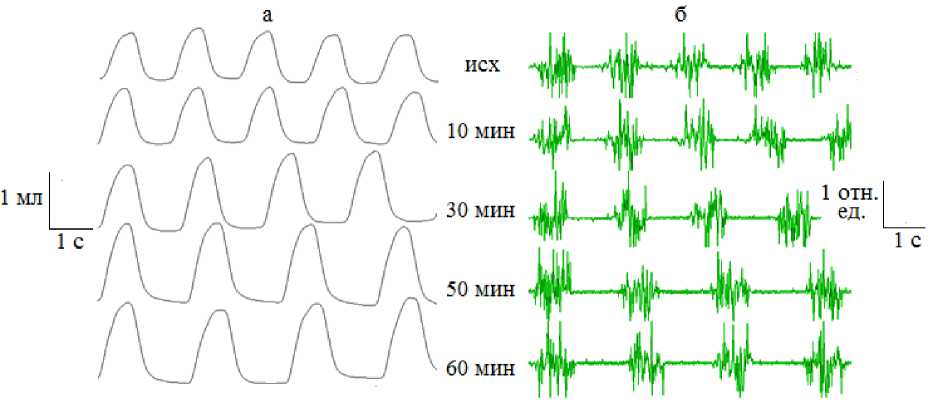
Рис. 3. Изменения абриса спирограмм (а) и ЭМГ диафрагмы (б) после микроинъекции 10-7 М раствора L-глутамата в ПТО у крыс
Результаты, полученные в настоящей работе с применением электрической и химической стимуляции ПТО у крыс, свидетельствуют об участии ПТО в регуляции дыхательной ритмики у млекопитающих живот- ных. Это в известной мере соответствует представлениям о ПТРГ как вероятном генераторе ритма дыхания [7], основанным на результатах изучения ПТО у низших позвоночных, в т.ч. на данных о прекращении дыхания после механического и химического разрушения указанной области у миноги [13].
Участие ПТО в респираторном контроле у крыс опосредуется глутаматергическими механизмами, активация которых снижает частоту инспираторных разрядов диафрагмы, а также уменьшает ЧД за счет значительной пролонгации экспираторной фазы. Существенное удлинение выдоха в ответ на введение L-глутамата в ПТО, на наш взгляд, может обеспечиваться двумя путями. Во-первых, при активации глутаматных рецепторов могут усиливаться залповые разряды премоторных экспираторных нейронов. Во-вторых, может происходить глутаматергическая активация тормозных нейромедиаторных механизмов, модулирующих разряды дыхательных нейронов, участвующих в респираторном ритмогенезе. В частности, у крыс это могут быть гистохимически и электрофизиологически идентифицированные фазовопереходные (I-E1, I-E2) и инспираторные нейроны, локализованные в пределах ПТРГ, взаимодействующие с дендритами нейронов пневмотаксического комплекса и направляющие свои аксоны в вентролатеральную часть моста [8]. Кроме того, в ПТРГ присутствуют нейроны, посылающие контралатеральные проекции в симметричную ПТРГ, а также премоторные интернейроны, образующие билатеральные связи со стволовыми респираторными мотонейронами [14]. В свою очередь нейроны понтомедуллярных районов мозгового ствола получают возбуждающие входы из каудальной вентральной респираторной группы [15]. Эти межнейронные связи позволяют говорить об участии ПТО как в генерации дыхательного ритма, так и в формировании паттерна дыхания.
В плане объяснения тормозного характера респираторных реакций на активацию глутаматных рецепторов ПТО у крыс представляют интерес исследования in vitro на мозге миноги, в которых установлено, что респираторные мотонейроны, направляющие аксоны в ПТРГ, обладают иммунореактивностью к глутамату и окружены глицинергическими клетками и ГАМК-иммунореактивными структурами. Блокада ГАМКА и глициновых рецепторов в моторных ядрах продолговато- го мозга увеличивает ЧД [7]. Однако исследователи полагают, что не ГАМК и не глицин, а только эндогенно высвобождаемые возбуждающие аминокислоты играют существенную роль в респираторном ритмогенезе [16]. В то же время ГАМК- и глицинергиче-ские входы к респираторным мотонейронам, например в пределах вагального комплекса, опосредуют изменения ЧД через модуляцию восходящих возбуждающих проекций к ПТРГ [17]. То есть восходящие глутаматер-гические пути к ПТРГ подвергаются мощным тормозным влияниям, а их растормаживание является одним из важнейших механизмов регуляции ЧД [7]. С учетом этих данных можно объяснить угнетение дыхания, наблюдаемое нами при микроинъекциях L-глутамата в ПТО крыс, как результат возбуждения имеющихся здесь ГАМКергических терминалей, выхода из них ГАМК и активации ГАМКА-рецепторов. Наличие последних в ПТО у грызунов подтверждено изменениями ларингеальных респираторных рефлексов и ЧД при микроинъекциях мусцимола в область ПТО у морских свинок [18].
Особенностью респираторных эффектов активации ПТО глутаматом оказалась их значительная продолжительность во времени, что, возможно, связано с метаботропной природой рецепторов и внесинаптическими влияниями экзогенного медиатора. В ряде работ установлено, что действие глутамата, как и ГАМК, не ограничивается только локальным постсинаптическим участком. Эти нейроактивные аминокислоты способны высвобождаться во внесинаптическое пространство за счет спилловера, т.е. растекания из синаптической щели [19]. Допустимо считать, что микроинъекции глутамата в ПТО приводят к широкому распространению данного медиатора по доступному пространству и активации определенных классов рецепторов, что способствует выделению эндогенной ГАМК. Имеются данные о том, что повышение содержания внеклеточной ГАМК может быть вызвано активацией каинатных рецепторов глутамата [20]. Высвобождающаяся ГАМК, в свою очередь, оказывает ингибирующее влияние на ритмогенерирующие и паттернформирующие нейроны, что ведет к снижению ЧД и легочной вентиляции у животных.
Заключение. Таким образом, ПТО у млекопитающих животных можно рассматривать как респираторно активную часть моста, участвующую в регуляции ритма и паттерна дыхания. Участие ПТО в респираторном контроле опосредуется глутаматерги-ческими связями, которые у крыс преимущественно включены в механизмы, регулирующие длительность экспираторной фазы и, следовательно, частоту дыхания.
Список литературы Респираторные эффекты активации паратригеминальной области у крыс
- Bianchi A.L., Denavit-Saubie M., Champagnat J. Central control of breathing in mammals: neuronal circuitry, membrane properties, and neurotransmitters. Physiol. Rev. 1995; 75 (1): 1-45.
- Сафонов В.А. Регуляция внешнего дыхания. Вестник СурГУ. Медицина. 2009; 1 (2): 13-22.
- Feldman J.L., Del Negro C.A. Looking for inspiration: new perspectives on respiratory rhythm. Nat. Rev. Neurosci. 2006; 7: 232-242.
- Huckstepp R.T.R., Cardoza K.P., Henderson L.E., Feldman J.L. Role of parafacial nuclei in control of breathing in adult rats. J. Neurosci. 2015; 35 (3): 1052-1067.
- Moraes D.J.A., Dias M.B., Cavalcanti-Kwiatkoski R., Machado B.H., Zoccal D.B. Contribution of the retrotrapezoid nucleus/parafacial respiratory region to the expiratory-sympathetic coupling in response to peripheral chemoreflex in rats. J. Neurophysiol. 2012; 108: 882-890.
- Cinelli E., Robertson B., Mutolo D., Grillner S., Pantaleo T. Bongianni F. Neuronal mechanisms of respiratory pattern generation are evolutionary conserved. J. Neurosci. 2013; 33 (21): 9104-9112.
- Cinelli E., Mutolo D., Contini M., Pantaleo T., Bongianni F. Inhibitory control of ascending glutamatergic projections to the lamprey respiratory rhythm generator. J. Neurosci. 2016; 326: 126-140.
- Song G., Yu Y., Poon C.-S. Cytoarchitecture of pneumotaxic integration of respiratory and nonrespiratory information in the rat. J. Neurosci. 2006; 26 (1): 300-310.
- Guyenet P.G., Bayliss D.A., Stornetta R.L., Ludwig M.-G., Kumar N.N., Shi Y., Burke P.G.R., Kanbar R., Basting T.M., Holloway B.B., Wenker I.C. Proton detection and breathing regulation by the retrotrapezoid nucleus. J. Physiol. 2016; 594 (6): 1529-1551.
- Alheid G.F., McCrimmon D.R. The chemical neuroanatomy of breathing. Respir. Physiol. Neurobiol. 2008; 164: 3-11.
- Villar-Cervino V., Barreiro-Iglesias A., Fernandez-Lopez B., Mazan S., Rodicio M.C., Anadon R. Glutamatergic neuronal populations in the brainstem of the sea lamprey, Petromyzon marinus: an in situ hybridization and immunocytochemical study. J. Comp. Neurol. 2013; 521: 522-557.
- Paxinos G., Watson C. The rat brain in stereotaxic coordinates. 6th ed. New York: Elsevier/Academic Press; 2007. 456.
- Mutolo D., Bongianni F., Cinelli E., Pantaleo T. Role of neurokinin receptors and ionic mechanisms within the respiratory network of the lamprey. Neuroscience. 2010; 169: 1136-1149.
- Gariepy J.-F., Missaghi K., Chartre S., Robert M., Auclair F., Dubuc R. Bilateral connectivity in the brainstem respiratory networks of lampreys. J. Comp. Neurol. 2012; 520: 1442-1456.
- Jones S.E., Saad M., Lewis D.I., Subramanian H.H., Dutschmann M. The nucleus retroambiguus as possible site for inspiratory rhythm generation caudal to obex. Respir. Physiol. Neurobiol. 2012; 180: 305-310.
- Bongianni F., Mutolo D., Cinelli E., Pantaleo T. Neural mechanisms underlying respiratory rhythm generation in the lamprey. Respir. Physiol. Neurobiol. 2016; 224: 17-26.
- Cinelli E., Mutolo D., Robertson B., Grillner S., Contini M., Pantaleo T., Bongianni F. Gabaergic and glycinergic inputs modulate rhythmogenic mechanisms in the lamprey respiratory network. J. Physiol. 2014; 592: 1823-1838.
- Driessen A.K., Farrell M.J., Mazzone S.B., McGovern A.E. The role of the paratrigeminal nucleus in vagal afferent evoked respiratory reflexes: A neuroanatomical and functional study in guinea pigs. Front. Physiol. 2015; 6: 378-391. URL: http://pubmedcentralcanada.ca/pmcc/articles/PMC4685097/(дата обращения: 15.02.2017) DOI: 10.3389/fphys.2015.00378
- Семьянов А.В. Диффузная внесинаптическая нейропередача посредством глутамата и ГАМК. Журнал высшей нервной деятельности им. И.П. Павлова. 2004; 54 (1): 68-84.
- Cossart R., Esclapez M., Hirsch J.C., Bernard C., Ben-Ari Y. GluR5 kainate receptor activation in interneurons increases tonic inhibition of pyramidal cells. Nat. Neurosci. 1998; 1 (6): 470-478.
