Responses of medicinal plants to drought: implications for secondary metabolite production

Author: Saini N., Vijayvergia R., Kumar A.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 2 т.21, 2025.
Free access
Drought places immense stress on the environment, threatening biodiversity, plant growth and productivity worldwide. This burden falls especially hard on medicinal plants grown in arid regions and semi-arid regions. When water is scarce, these plants undergo significant changes in their biochemical and physiological processes. They close their stomata, reduce photosynthesis, and alter their water management processes. These shifts are often accompanied by biochemical adjustments, such as changes in osmotic balance, antioxidant enzyme activity, accumulation of compatible solutes, and modifications in phytohormone levels. Additionally, drought stress can influence the production of secondary metabolites, which are critical for the therapeutic properties of medicinal plants. This review aims to provide a comprehensive overview of the physiological, biochemical, and molecular responses of medicinal plants to drought stress, with a focus on how these responses affect the synthesis and accumulation of secondary metabolites. Understanding these mechanisms is crucial for improving the cultivation, conservation, and therapeutic quality of medicinal plants under changing environmental conditions.
Drought, signaling, toxicity, tolerance, secondary metabolites, medicinal plants
Short address: https://sciup.org/143184709
IDR: 143184709
Text of the review article Responses of medicinal plants to drought: implications for secondary metabolite production
Abbreviations: AAO - ABA Aldehyde oxidase; ABA- Abscisic acid; ABF - ABRE binding factors; ABRE - ABA– responsive element; APX - Ascorbate peroxidase; AREB -ABA responsive element binding protein; Brs – Brassinosteroids; DHAR - Dehyd roascorbate reductase glutathione; DRE/C - Dehydration – responsive element / C repeat; GA - Gibberellic acid; GABA - Gamma –amino butyric acid; JA - Jas monic acid; LEA - Late Embryogenesis Abundant; MAS - Marker assisted selection; MDHAR - Monodehydroascorbic acid reductase; NCED - 9-cis epoxy carotenoid dioxygenase; OAT - Ornithine- delta –aminotransferase; P5CS -Pyrrolline -5-carboxylate synthase; QTL- Quantitative Trait Loci; ROS - Reactive oxygen species; SA - Salicylic Acid; SLs - Strigolactones; SOD - Superoxide dismutase
Drought ranks as one of the worst natural disasters on Earth, with effects on ecosystems, agriculture, and the environment. Unlike sudden catastrophes, drought builds up gradually but can cause widespread and complex problems, making it harder to deal with (Wilhite, 2016). It occurs when there is low rainfall for an extended period, along with high temperatures and low relative humidity (Lipiec et al., 2013), which disrupts the water balance and leads to significant hydrological imbalances. The World Meteorological Organization (1992) defines it as a prolonged period of deficient precipitation, resulting in a severe water shortage. As a weather-related threat, drought can negatively impact the economy, society, and environment. It affects both wealthy and impoverished nations, with the consequences varying depending on the type of ecosystem.
In areas with dense flora and low soil moisture, prolonged dryness can lead to plant mortality, habitat degradation, and accelerated vulnerability to climate change (Zhou et al., 2019 ) . The lack of water affects all living organisms, which leads to biodiversity loss, increased ailment amongst wildlife, and habitat destruction. Forests, especially, are distinctly sensitive to drought and climate change (Xu et al ., 2024), with water deficiency inflicting decreased plant growth, soil erosion, and an increase in the frequency and severity of wildfires (Boer et al., 2020).
Plants serve as vital sources of numerous medicinal compounds. These phytochemicals are involved directly or indirectly in plant growth and development. Plant metabolites are responsible for reducing various diseases. Since ancient times, people have used plants and their byproducts as traditional medicine worldwide. Plants with healing properties play a key role in traditional healing practices but also in the global trade of primary goods. Many pharmaceutical drugs used today come from natural products or their chemical analogs, which shows the importance of plant-based compounds. Moreover, drought stress directly influences plant growth by inhibiting physiological processes such as photosynthesis, water absorption, and nutrient uptake, in the long run leading to reduced crop yields and changed plant development. In the case of medicinal vegetation, drought can significantly adjust the production of secondary metabolites, which can be accountable for their therapeutic uses. Understanding the effects of drought on medicinal plants is essential for improving their cultivation, ensuring sustainable production, and maintaining the efficacy of plant-based medicines.
This review provides a comprehensive examination of the physiological and biochemical responses of medicinal plants to drought stress. By exploring the mechanisms behind the production of secondary metabolites under water shortage conditions, this review aims to shed light on the complicated interactions between drought stress and plant metabolism. Understanding these processes is crucial for developing processes to enhance drought tolerance, optimize the cultivation of medicinal flora, and ensure the sustainable production of high-quality therapeutic compounds.
EFFECTS ON PLANT GROWTH AND
DEVELOPMENT
Impact of Water Stress on Plant Growth
Drought stress impacts plant growth at multiple degrees, from metabolic activity to biological functions (Blumwald et al., 2004 ) . The primary physiological modifications arise at the level, affecting processes together with cellular division, expansion, and differentiation (Lestari et al., 2023), all of which might be critical for overall plant growth. Hsiao (1973) concluded that water stress limits cell expansion more extensively than cellular compartments, which leads to stunted growth and reduced plant length. This restriction on cellular growth reduces the quality and amount of plant tissues, negatively impacting all over biomass production.
At the biochemical stage, drought stress disrupts several critical physiological processes, which include hormone production, photosynthesis, respiration, ion uptake, and nutrient metabolism (Qiao et al., 2024). These disruptions inhibit the plant's potential to perform fundamental life processes, such as energy production and nutrient distribution, which are very important for growth and development. The effects of these disruptions are especially mentioned in younger plants and seedlings, where growth inhibition can lead to significant reductions in survival rates.
Structural Changes under Drought
Drought also triggers complicated structural changes within plants, specifically in leaf and root tissues. Water stress has been observed to increase leaf thickness, as well as alter the structure of palisade and spongy tissues in the adaptation phases ( elissen et al ., 2018; Meng et al., 2021). These changes often serve to restrict water loss however at the expense of lesser leaf expansion and lower photosynthetic capacity.
One of the important processes plants use to live on water scarcity is modifying the root-shoot ratio, which tends to increase under drought conditions (Reinelt et al., 2023). This shift prioritizes root growth, allowing plants to get access to deeper water reserves within the soil. However, this adaptive response comes with tradeoffs, such as decreased leaf area and reduced photosynthetic ability (Fig. 1), in the long run mainly to decrease agricultural yields. In species like Populus deltoides , Morus alba , and Dalbergia sissoo (Thakur and Sood, 2005), drought has been shown to severely inhibit leaf growth, further contributing to decreased productiveness. The reduction of leaf growth, in turn, limits carbon assimilation, which leads to decreased biomass accumulation (Wei et al., 2024).
Impacts on Seed Production and Yield
Water shortage additionally has far reached effects on reproductive procedures in plants, which include seed production and yield. Drought conditions usually reduce the production of plants and seeds, and impair the development of vital yield components. Seed set, size, and quality are frequently compromised, leading to decreased yields (Albert et al., 2024), which is a big concern in both herbal and agricultural ecosystems. Water stress prevents the accumulation of fresh plant mass, further reducing both seed output and the overall health of drought-affected populations. As a result, drought-tolerant varieties of plants are often characterized by lower yields, despite their ability to survive harsh conditions.
EFFECT ON PLANT PHYSIOLOGY
Plants respond to drought through a variety of physiological changes, often involving alterations in their metabolic processes. Water stress disrupts key physiological structures, mainly modifications in growth rate, tissue osmotic capability, and antioxidant defenses. These changes permit the plant to deal with water deficits, but additionally, they come at the cost of decreased productivity and growth (Duan et al., 2007).
Stomatal Closure
One of the instant physiological responses to drought is the closure of stomata, which are small pores on the leaf surface that regulate gas exchange. Stomatal closure is an adaptive reaction to decrease water loss via transpiration. However, it also restricts the uptake of CO 2 , which is essential for photosynthesis, leading to decreased photosynthetic processes (Allen and Ort, 2001). As drought conditions persist, plants may revel in a decline in the efficiency of the photosynthetic process, which is responsible for carbon fixation and production of energy.
The mechanism of stomatal closure under drought is given in figure 2. The closure of stomata underneath drought is closely connected to leaf water status and stomatal conductance, both of which decline as moisture level of soil drops. Research has validated a positive correlation between leaf water capacity and stomatal conductance, with drought-induced soil drying playing an important role in regulating this process (Xia et al., 2024). The restriction of CO 2 absorption due to stomatal closure in the long run reduces the plant's capacity for carbon assimilation, proscribing energy production and growth.
Disruption of Photosynthesis
Drought-induced stress affects stomatal conductance and also impairs the core factors of the photosynthetic process. Key processes, consisting of the Calvin cycle and electron transport inside the thylakoid membranes, are disrupted, resulting in the accumulation of reactive oxygen species (ROS) and raised oxidative stress. The harm to photosynthetic membranes and the destruction of lipids via peroxidation in addition decreases the plant's ability to transform light energy into chemical energy.
Multiple studies have indicated that both stomatal and non-stomatal elements contribute to the decline in photosynthetic ability under drought stress (Beauclaire, 2024). Stomatal closure prevents the entry of CO 2 , while non-stomatal factors, such as electron transport and enzyme activity and loss of photosynthetic capacity exacerbate. Together, these disruptions result in significant decrease in plant growth and productivity, highlighting the profound effect of drought on plant physiology.
The physiological and biochemical responses of plants to drought are multifaceted and involve complex tradeoffs between survival and productivity. While stomatal closure and structural adaptation, such as altered root structure, allow flora to conserve water, these same mechanisms compromise photosynthetic ability and growth. Ultimately, the ability of plants to balance these tradeoffs determines their resilience to drought and their ability to recuperate as soon as water becomes available again.
Chlorophyll Content
One of the major signs of oxidative stress in flora during drought is a reduction in chlorophyll action. This decrease is primarily responsible for chlorophyll degradation and pigment photooxidation. Chlorophyll plays an essential role in photosynthesis, facilitating light absorption and the production of reducing agents important for energy production. As drought stress intensifies, chlorophyll a and b levels are significantly reduced which is vital for capturing light energy. However, the degree of chlorophyll reduction varies throughout species and depends on both the duration and severity of drought conditions (Mafakheri et al ., 2010). In a few instances, drought can also cause a marked decrease in chlorophyll content, while other species showcase minimum changes, reflecting their unique physiological processes for dealing with water stress.
The decline in chlorophyll disrupts light capture as well as impacts the complete photosynthetic mechanism, reducing the plant's capability to produce energy. The resulting oxidative stress can result in the formation of ROS, which similarly hastens chlorophyll breakdown and damages cellular structures involved in photosynthesis. Understanding the dynamics of chlorophyll degradation under drought is important for developing drought-tolerant plant varieties with sustained photosynthetic activity underneath stress.
Osmotic Adjustment
Osmotic adjustment is a vital physiological mechanism that is used by plants to maintain cell turgor throughout drought conditions. This system involves the accumulation of organic and inorganic solutes, like proline, glycine betaine, and soluble carbohydrates, in the cytoplasm of leaf cells (Bhutto et al., 2023). These osmolytes assist in lowering the osmotic ability of the cell, facilitating water retention and enabling the plant to extract moisture from increasingly dry soils.
The effectiveness of osmotic adjustment is especially dependent on the rate and severity of water stress. As soil moisture decreases, plants either increase water uptake by means of adjusting osmotic ability or lessen water loss via closing stomata. By gathering solutes, plants create a more negative osmotic capacity, which in turn induces cellular hydration and enables the structural integrity of metabolically active tissues, like leaf cells. This physiological adaptation enables plants to continue vital metabolic features under water deficit conditions, thereby improving drought tolerance and lowering harm to plant tissues. Furthermore, variations in leaf temperature under drought conditions can affect water status by affecting transpiration rate and stomatal conductance. Plants capable of efficient osmotic adjustment can optimize those processes, balancing water absorption with minimal water loss.
Reactive Oxygen Species (ROS) and Antioxidant Defense
Drought stress leads to the creation of ROS that may disrupt cellular homeostasis and damage physiological and biochemical pathways. The overproduction of ROS in plant cells under drought, such as superoxide radicals and hydrogen peroxide, results in oxidative stress that compromises membrane integrity and cell function. To counteract these harmful outcomes, plants set off their antioxidant defence systems, composed of both enzymatic and nonenzymatic components.
The enzymatic antioxidants involved in mitigating ROS harm encompass ascorbate peroxidase, catalase, glutathione reductase, and dehydroascorbate reductase (DHAR), which neutralize reactive oxygen molecules and prevent cell damage. onenzymatic antioxidants, such as ascorbic acid (vitamin C), carotenoids, and tocopherols (vitamin E), additionally play an important role in shielding plant cells from oxidative damage with the aid of scavenging ROS (Choudhury et al., 2017).
The level of antioxidant enzyme activity varies substantially among drought-tolerant and droughtsensitive cultivars. Plants with a better antioxidant reaction exhibit more advantageous drought resistance, as their capability to neutralize ROS minimizes the harm to cellular systems and guarantees persistent metabolic function. However, withstanding the activation of these defense mechanisms, extended or intense drought can weigh down the plant’s adaptive capacity, leading to irreversible harm.
ROLE OF PHYTOHORMONES INDROUGHT TOLERANCE
Phytohormones are principal regulators of nearly all organic processes in flora, from growth and development to their adaptive responses to environmental stressors like drought. During water scarcity, plants rely on these hormones to coordinate complicated physiological responses that mitigate drought’s adverse results. Drought triggers the discharge and modulation of phytohormones, which set off numerous mechanisms (Fahad et al., 2017), including osmotic regulation, stomatal closure, and root growth adaptation, permitting plants to deal with reduced water availability. These hormones either act as positive signals, enhancing drought tolerance by means of increasing osmoprotectant accumulation and decreasing water loss, or as negative signals, diminishing plant growth and productiveness under water stress (Yang et al., 2021). Phytohormone-mediated drought response in plants is given in figure 3.
Abscisic Acid (ABA)
Abscisic acid is the most considerably studied phytohormone on the subject of drought tolerance, playing an essential role in regulating plant responses to water deficit. ABA is pivotal for stomatal closure, supporting to prevent water loss through transpiration. During drought, ABA is synthesized in the roots and transported to the shoots, in which it alerts stomatal closure by regulating ion delivery in guard cells (Fig. 2), thereby reducing water loss. This procedure entails ABA-dependent activation of ion channels, which facilitate the elimination of K+ ions and anions from guard cells, resulting in turgor loss and stomatal closure (Kim et al., 2010; Sah et al., 2016).
ABA additionally regulates the expression of stress responsive genes and transcription elements involved in drought tolerance. For instance, the expression patterns of aquaporin genes, which control water transport and cellular turgidity, are modulated with the aid of ABA, helping plants maintain cellular hydration throughout water scarcity. Additionally, ABA is a key player in inter phytohormone crosstalk, interacting with different hormones like jasmonic acid (JA) and brassinosteroids (BR) to coordinate the plant’s stress response. Research has shown that those hormones, in combination with nitric oxide ( O), work synergistically with ABA to modulate stomatal dynamics and increase drought resistance (Daszkowska-Golec and Szarejko, 2013).
The mechanisms by which phytohormones adjust stomatal closure during drought is summarized in Fig. 2. ABA acts as the primary modulator, taking part in the exchange that enhances the stomatal closure reaction. Ion flux changes in guard cells, which results in the lack of turgor and the closing of stomata, thereby minimising water loss via transpiration.
Auxin
Auxin, particularly indole 3 acetic acid (IAA), has been increasingly known for its role in drought stress responses. Studies have proven that auxin regulates plant growth under drought conditions by modulating root structure and improving the plant's water absorption capacity (Shi et al ., 2014). Auxin facilitates lateral root formation, which will increase the surface location for water uptake and improves root elongation, allowing plants to get deeper soil water throughout drought.
Additionally, auxin has an exchange relationship with ABA, influencing the plant’s drought stress tolerance by affecting ABA production (Du et al., 2012). High auxin levels correlate with stronger drought tolerance, probably due to their function in retaining root growth and cell turgor during water stress.
Gibberellin (GA)
Gibberellic acid (GA) plays a twin role in plant growth regulation and drought stress tolerance. While GA is commonly related to stem elongation, seed germination, and reproductive development, its content decreases under drought stress, leading to a growth-repressing effect that aids in drought tolerance.
The accumulation of DELLA proteins, which act as repressors of GA signalling, is a key adaptive mechanism during drought. Increased DELLA protein level in response to drought restricts cell elongation and growth, allowing plants to preserve resources and increase survival under stress. This growth restraint, although decreasing biomass accumulation, enhances the plant’s overall stress tolerance by way of allocating energy towards survival as opposed to growth during drought conditions (Shohat et al. 2021).
Cytokinin
Cytokinin is concerned with regulating cell division, growth, and development, and it plays an enormous function in the plant’s drought response (Hai et al., 2020). Cytokinin levels generally decrease under drought stress, which is correlated with the inhibition of shoot increase, leaf senescence, and reduced photosynthetic capacity (Glanz-Idan et al., 2020). Cytokinins have a hostile relationship with ABA, as they promote growth processes that are suppressed using ABA at some stage in drought.
In drought-stressed plants, the reduction in cytokinin content results in the restriction of shoot growth and a shift in energy allocation in the direction of root development and osmotic adjustment, enabling plants to better deal with water scarcity. This cytokinin-induced adaptation process enables plants to enhance their drought tolerance even while conserving water and reducing the surface area of the leaf, which limits transpiration losses (Rivero et al., 2009).
Jasmonic Acid (JA)
Jasmonic acid, a lipid-derived phytohormone, plays a critical function in the growth, development, and defense mechanisms of plants, such as root formation, seed germination, and development of flowers. One of the earliest pieces of evidence of JA's function in drought stress came from studies on barley leaves, wherein drought stress was introduced by way of sorbitol treatment. This resulted in a major increase in jasmonate level, which resulted in activated downstream genes related to stress tolerance (Kramell et al., 2000). Further studies showed that jasmonate synthesis is associated with greater drought tolerance, supporting the concept of its involvement in drought stress (Lopes et al., 2024).
JA interacts synergistically with ABA, the primary hormone involved in stomatal closure, to increase drought responses. This exchange amplifies ABA signaling, resulting in elevated stomatal closure, thereby decreasing water loss throughout drought conditions.
Brassinosteroids (BRs)
Brassinosteroids are polyhydroxy steroid phytohormones found in numerous plant tissues which include pollen, immature seeds, flowers, and roots. They are significant for their roles in regulating plant growth processes such as seed germination, leaf bending, and ethylene production. In response to drought, BRs considerably make contributions to the enhancement of plant tolerance via the production of ROS (Salvi et al ., 2021). This expanded ROS generation, coupled with greater antioxidant defense mechanisms, enables flora to deal with drought stress. In drought conditions, BRs also promote cellular elongation and stimulate root development, which aids in water uptake. Furthermore, BRs positively affect ABA signalling, helping in the regulation of stomatal aperture and mitigating water loss during low water availability (Zhang et al., 2024).
Strigolactones (SLs)
Strigolactones, carotenoid-derived phytohormones, have emerged as a major player in plant adaptation to environmental stresses, which include drought. SL levels were observed to rise in plants under drought stress, indicating their involvement in stress tolerance (Ha et al., 2014). One of the key mechanisms through which SLs contribute to drought tolerance is by enhancing ABA sensitivity. The increased sensitivity permits quicker stomatal closure and reduces water loss, as a consequence aiding in drought survival.
In addition to regulating stomatal conductance, SLs also increase root structure by promoting lateral root development (Song et al., 2023), which helps vegetation discover deeper soil layers for water.
Salicylic Acid (SA)
Salicylic acid is generally known for its vital role in plant defense towards biotic stresses. However, its involvement in abiotic stresses, especially drought, has recently won interest. Studies suggest that SA regulates drought responses by modulating stomatal aperture and influencing gene expression associated with drought tolerance. The impact of SA on drought tolerance is dose-dependent, with lower concentrations frequently promoting tolerance, even as higher concentrations might also set off drought sensitivity (Catala et al ., 2007).
Interestingly, in some instances, high levels of SA treatment were shown to decrease drought tolerance in maize, highlighting the want for fine-tuned regulation of SA during drought stress ( emeth et al., 2002).
Ethylene
Ethylene is a gaseous phytohormone commonly recognized for its role in fruit ripening, flower senescence, and leaf abscission. In drought conditions, the production of ethylene will increase, which interacts with ABA to regulate stomatal closure. While ABA slows down stomatal closure, ethylene hastens this process by enhancing ROS production through ADPH oxidase activity in guard cells (Desikan et al., 2006).
Ethylene's twin function during drought not only includes promoting stomatal closure but also regulating root growth, ensuring that plants maintain water uptake under constricted water conditions. Additionally, ethylene's interplay with other hormones, like auxins and cytokinins, performs a crucial role in coordinating the plant's growth and stress responses during drought.
The coordinated moves of these hormones make sure that plants modulate their growth, stomatal function, and gene expression to survive during the course of limited water availability.
PERCEPTION, SIGNALING AND RESPONSES TO DROUGHT STRESS IN PLANTS
Plants have developed complex mechanisms to adapt to various environmental stressors, such as drought. These mechanisms include stress-specific and cross-protective responses that enable plants to tolerate multiple environmental challenges. The signal transduction pathways involved in drought and other abiotic stresses like salinity are highly complicated, with many aspects not understood fully.
When plants are exposed to water scarcity, root cells are the first to understand the environmental stress (Yang et al ., 2021). This preliminary signal triggers a cascade of secondary messengers, together with inositol phosphate, calcium ions, and ROS (Huang et al. 2012). These secondary messengers play an essential role in modulating cytosolic calcium levels, which might be detected with the aid of calcium-binding proteins. These proteins go through nonenzymatic conformational adjustments upon calcium binding, initiating a phosphorylation cascade that activates key stress-responsive genes and transcription elements. The activation of these stress-related genes brings about various physiological and biochemical adaptations, which in the end help the plants adapt to water stress conditions. Additionally, phytohormones play a pivotal function in regulating stress responses through gene expression changes.
Among the phytohormones, ABA is imperative to the drought stress response. Drought conditions enhanced the ABA synthase activity which subsequently increase the production of ABA in plants (Yang et al., 2021). The buildup of ABA initiates the activation of drought responsive genes through two unique pathways: ABA dependent and ABA independent (Fujita et al., 2005).
The ABA responsive element (ABRE) and the drought responsive element/C repeat (DRE/C) in the promoters of these genes are important in regulating drought tolerance. The major transcription factors, AREB (ABA responsive element binding protein) and ABF (ABRE binding factors), are involved in inducing the expression of ABA responsive genes (Uno et al., 2000).
These transcription factors are activated through ABA dependent phosphorylation. Other transcription factors including MYC, MYB, and AC additionally play a role in fine-tuning the plant's drought response, similarly highlighting the importance of ABA in controlling drought tolerance mechanisms. These interactions assist modulate water loss via stomatal regulation, increase root growth, and trigger other adaptive responses important for drought survival (Fig. 4).
Understanding the perception and signalling pathways involved in drought stress is crucial for increasing plant tolerance during environmental stresses. The role of phytohormones, especially ABA, in activating stress-responsive genes is highlighted in figure 4. Secondary metabolites further contribute to plant survival during drought by regulating defense mechanisms, signalling, and physiological responses.
SECONDARY METABOLITES AND THEIR SIGNIFICANCE
Plants produce a collection of chemical compounds called metabolites, which are categorized into primary and secondary metabolites. While primary metabolites are important for development and growth, secondary metabolites have specialized roles, often related to plant survival under stressful conditions. Secondary metabolites do not contribute directly to the basic physiological procedures of growth, but they play important roles in defense mechanisms, signalling, enzyme regulation, and interspecies communication. For example, some secondary metabolites act as repellents in opposition to herbivores, while others facilitate symbiotic relationships with pollinators or useful microbes.
Drought stress not only impacts primary metabolic processes but also considerably alters secondary metabolite production in medicinal plants. Secondary metabolites, which include alkaloids, phenolic compounds, and terpenoids, are critical for the plant's defense to resist environmental stressors and are at once related to their medicinal uses.
Under drought conditions, plants often increase the production of certain secondary metabolites as part of their defense mechanism. For instance, phenolic compounds, which function as antioxidants, are upregulated in many drought-tolerant species to mitigate the harm caused by ROS. Similarly, alkaloids, recognized for their role in plant defense, may also increase, enhancing the plant’s resistance to pathogens and herbivores during drought conditions.
However, the effects of drought on secondary metabolite production can range through species and stress severity. While a few compounds are more desirable, others may additionally lower due to the overall reduction in plant biomass and growth. Understanding the complicated interplay between drought stress and secondary metabolite production is vital for developing strategies to enhance the healing properties of medicinal plants in arid conditions.
Classification of Secondary Metabolites
Secondary metabolites are commonly grouped into three major classes:
-
1. Terpenoids
-
2. Phenolic compounds
-
3. Alkaloids
Terpenoids constitute one of the largest and most diverse classes of secondary metabolites. These compounds are composed of isoprene units and might range in size from five-carbon hemiterpenes to complex polycyclic molecules. Terpenoids play critical ecological roles, including serving as insecticides, like artemisinin, a sesquiterpenoid lactone from Artemisia annua . Terpenoids also have therapeutic properties, such as immunomodulatory, antibacterial, and antifungal activities.
Phenolic compounds are another commonly distributed group of secondary metabolites. They range from easy molecules like vanillin and gallic acid to more complex systems including polyphenols (stilbenes and flavonoids). Flavonoids, particularly, are water-soluble pigments saved in vacuoles of plant cells. These compounds play key roles in plant pigmentation, symbiotic nitrogen fixation, and plant defense.
Alkaloids are nitrogen-containing compounds derived from amino acids. Found predominantly in plants, alkaloids are recognized for their potent biological activities and are often used in pharmacology. Examples of alkaloids consist of morphine and codeine (analgesics), vinblastine and campthothecin (anticancer promoters), and scopolamine as a sedative (Crozier et al., 2006).
Functions of Secondary Metabolites
Plant secondary metabolites are organic compounds that are involved in various functions of plant defense and protection. Stress like drought, salinity, changes in temperatures, pollutants etc. affects plant growth and development. Secondary metabolites reduce the effect of both abiotic and biotic stress. Secondary metabolites made by plants can affect the biochemistry of humans and animals. The high concentration of these substances has a wide range of effects, from being immediately fatal to being beneficial or restorative. These molecules represent an important source of active pharmaceuticals and hence, have a good therapeutic potential.
It is commonly known that secondary plant metabolites play a vital role in the complex relationship between plants and their surroundings. Examples of these roles include protecting plants from infections of pathogens and herbivores and attracting pollinators (Wink, 2010). Regarding abiotic stress, there are several possible uses for secondary metabolites, such as defense against ultraviolet light, excessive light acting as suitable solutes for radical scavenging, or lowering transpiration. A natural product's actual production and accumulation are frequently impacted and governed by a multitude of circumstances. Under drought secondary metabolites reduce water content in leaf cells and stimulate stomatal closure for decline water loss.
Roles of secondary metabolites in medicinal plants
Plants are living chemical factories that synthesize several secondary metabolites. These secondary plant products are the building blocks of many pharmaceutical medications that are sold commercially and used in herbal therapies. The various biological components of plants have several activities that not only have substantial value in the agricultural, cosmetic, and perfume occupations but can also improve people's wellness through the sectors of food and medicine. Many secondary metabolites are being explored for use in medication research, including phenylpropanoids, terpenoids, and alkaloids.
Plants with specific properties that are employed in herbalism are referred to as medicinal plants. They are believed to provide a rich supply of botanical compounds that can be utilized as tools in the medication development process. That herbs can be taken as preventative, or synergistic medication. Alkaloids, glycosides, tannins, and other chemical entities with distinct activities are among the many different types of substances found in plants. The production of these valuable phytochemicals varies with the seasons, habitat, and other climate factors. umerous phytocompounds that are found in a variety of therapeutic plants have been identified, some phytochemicals and their medicinal properties are given in Table 1. Many of these substances are utilized as dietary supplements in our daily lives (such as curcumin from Curcuma longa ), due to their therapeutic characteristics, certain cosmetics contain ingredients used as mouth rinse.
DROUGHT-INDUCED CHANGES IN
PHYTOCHEMICAL PRODUCTION
Impact of drought on primary metabolites
Water deficiency leads to changes in plant primary and secondary metabolites. As a result of drought stomata close and fixation of CO 2 decreases. These changes occur because the Calvin cycle is also affected by drought. The activity of the enzyme ribulose-1, 5-bisphosphate carboxylase (Rubisco) decreased, which affected photosynthesis. The reduction in Rubisco activity leads to a drop in stromal fructose bisphosphatase (Zhou et al., 2007).
Drought also diminishes the functioning of various photosynthetic enzymes to varying degrees. Furthermore, the amount of CO 2 in the chloroplast regulates the electron transport chain involved in photosynthetic action, and dry circumstances frequently cause a reduction in PS II function (Qiao et al., 2024). Drought impairs electron transfer in both cyclic and non-cyclic forms during photosynthesis' light reaction in addition to its detrimental impact on the dark reactions of photosynthesis (Obidiegwu et al., 2015).
The pigments in the cells of leaves are reduced by water stress. Moderate water stress has a higher effect on the levels of carotenoids than severe drought conditions. Irrigation schedules had no discernible effect on the amount of chlorophyll b; however, they did increase under mild water stress and decrease in severe water deficiency conditions. During drought, a rise in the synthesis of photosynthetic pigments. The concentration of myo-inositol and sucrose in primary metabolites increases due to water deficiency and these osmolytes help plants to maintain water levels in cells. Due to drought, plants also increase the precursors of raffinose (Griesser et al ., 2015).
Alterations in secondary metabolites profiles under drought stress
In water deficiency conditions, primary metabolism is altered and secondary metabolite production commenced. Changes in secondary plant products concentrations in stressed plants are predicted by the theory that the source-sink carbon nutritional equilibrium and secondary substances based on carbon correlate ( iinemets, 2016). This concept states that any stress that inhibits photosynthesis to a greater extent causes secondary compounds to accumulate. It appears that the impacts of drought can differ for different compound classes and be correlated with the severity of the drought, however, the data is limited. Plants exposed to the stress showed the highest rise in alkaloid, terpenoid compounds, and tannin amounts, while total phenols, saponins and flavonoids dramatically decreased. The products of primary metabolism usually contain less carbon than secondary metabolites, including phenolics and isoprenoids, which are usually greatly reduced under drought stress.
Water deficit conditions also have a favourable impact on the content of compounds containing nitrogen, such as cyanogenic glucosides, glucosinolates, and alkaloids. Under water stress, essential oil yield dropped while essential oil content increased. Furthermore, plants may produce high terpene concentrations in water-stressed environments. Under drought also an increase in essential oil concentration. Some reports also support that the concentration of phenolics and terpenoids increases. In grapevine, an increment in polyphenols (85%) and volatile organic compounds (40%) was recorded under drought conditions (Griesser et al., 2015).
MECHANISMS REGULATING METABOLIC
PRODUCTION UNDER DROUGHT
CONDITIONS
Drought stress is one of the most significant abiotic elements that impact plant growth, productivity, and metabolism. When the availability of water is less, plants go through a series of physiological, biochemical, and molecular changes to adapt and survive. These adaptations are tightly regulated via gene expression, enzyme activation, and metabolic shifts. The complex mechanisms regulating plant metabolism under drought, specializing on gene expression adjustments, enzymatic regulation, and the interplay between drought stress and metabolite synthesis are summarized here.
Gene Expression Changes in Response to Drought Stress
Drought stress triggers changes within the expression of numerous genes, including those that are involved in the production of secondary metabolites and stress-related proteins. The response to drought is initiated by using sensors located within the plasma membrane, which detect deficiency of water and spark off downstream signalling cascades. These signalling pathways often contain the activation of transcription factors, secondary messengers (e.g., ROS, calcium ions), and phytohormones like ABA. ABA is a central participant in response to drought, regulating stomatal closure and modulating gene expression for drought tolerance (Fig. 2 and 4).
Key drought-responsive transcription factors
Plants have evolved complex molecular mechanisms to sense and respond to drought, including the regulation of specific transcription factors (TFs). These TFs act as mail regulators in coordinating gene expression and activating signalling pathways that help the plant tolerate drought.
MYB: Regulates secondary metabolite synthesis under drought.
AREB1, AREB2, ABF1: Upregulated by using ABA and play vital roles in activating drought-responsive genes (Aerts et al., 2024).
Furthermore, ABA biosynthesis is upregulated during drought via enzymes including ABA aldehyde oxidase (AAO) and 9 cis epoxy carotenoid dioxygenases ( CED), resulting in higher ABA concentrations and drought resistance (Yang et al. , 2021). ABA signalling additionally promotes the production of protecting proteins, like Late Embryogenesis Abundant (LEA) proteins, which help maintain cellular stability during water stress (Wang et al., 2024).
Proline, a major osmolyte, accumulates during drought due to the upregulation of pyrroline 5 Carboxylate synthase (P5CS), an enzyme that is involved in proline biosynthesis (Parida et al. 2008). Proline acts as an osmoprotectant, which helps in maintaining cell turgor and reducing water loss.
Enzymatic Pathways and Regulation under Drought
The accumulation of ABA during drought leads to stomatal closure, reducing transpiration and restricting water loss. However, stomatal closure also decreases CO₂ uptake, which impairs photosynthesis and the Calvin cycle.
Key enzymes affected by drought
Several key enzymes are involved in regulating drought responses by modulating metabolic pathways such as osmotic adjustment, oxidative stress management, and cellular repair mechanisms. These are
Rubisco : The number one enzyme in carbon fixation, whose activity declines during drought, decreasing photosynthetic efficiency.
Antioxidant enzymes: Superoxide dismutase (SOD) and ascorbate peroxidase (APX) enzymes that scavenge harmful ROS generated during drought. These enzymes play important roles in protecting plant cells from oxidative damage via neutralizing superoxide radicals and other ROS. Some scavenging enzymes are regulated by drought mechanisms, these are dehydroascorbate reductase, monodehydro-ascorbate reductase, catalase and peroxidase.
Proline biosynthesis enzymes : Under drought, proline biosynthesis will increase through the activation of P5CS and Ornithine delta aminotransferase (OAT), at the same time as the breakdown of proline by means of proline oxidase decreases, ensuing in proline accumulation, which enhances drought tolerance ( iinemets, 2016).
Interaction between Drought Stress and Metabolite Synthesis
Drought stress appreciably alters plant metabolism, affecting each primary and secondary metabolic pathway. The reduction in photosynthesis because of stomatal closure impacts the Calvin cycle and glycolysis, which results in the accumulation of certain metabolites and the reduction of others.
Metabolite changes under drought stress
Increased biosynthesis of osmoprotectants: Compounds like proline, trehalose, and sucrose accumulate to maintain balance and protect the cellular structure.
Phytohormones: Levels of stress-related hormones like ABA, JA, and SA increase, regulating numerous stress response pathways.
Secondary metabolites : The production of secondary metabolites, including terpenoids, alkaloids, and tannins, is often upregulated for the duration of drought, enhancing the plant’s capacity to tolerate stress. These compounds act as antioxidants, scavenging ROS and protecting the plant from oxidative damage.
However, drought can also negatively affect the production of other metabolites. For instance, phenolics and flavonoids, which are important for UV protection and defense towards herbivores, can also decrease under drought stress.
Drought stress
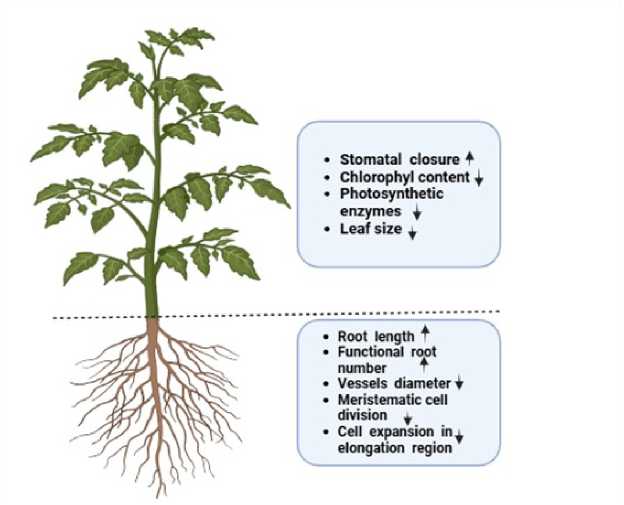
Figure 1. Physio-morphological responses of plants under drought.
increase
ABA
increases _ calcium “ concentration
Potassium release from guard cells
Concentration ofH* increase in

guard cells
H:O release from guard cell
Stomata close
Figure 2. Mechanism of stomatal closure under drought.
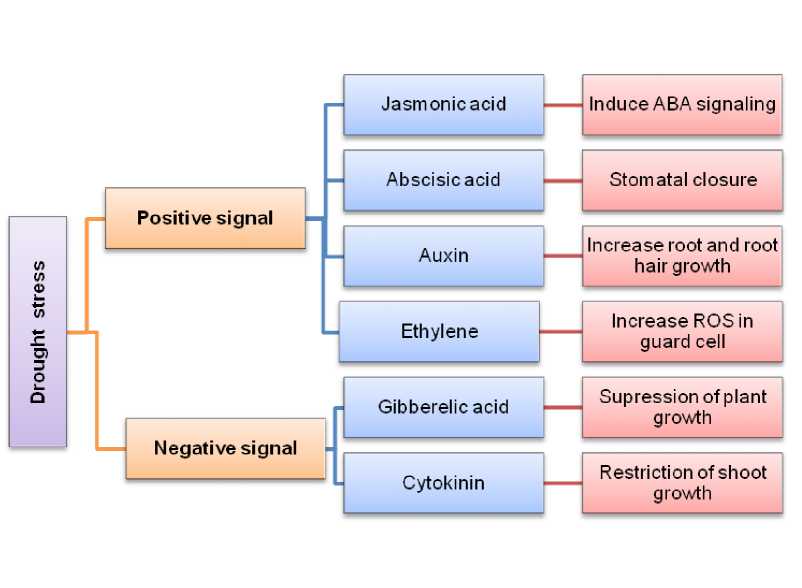
Figure 3. Phytohormone mediated drought response in plants.
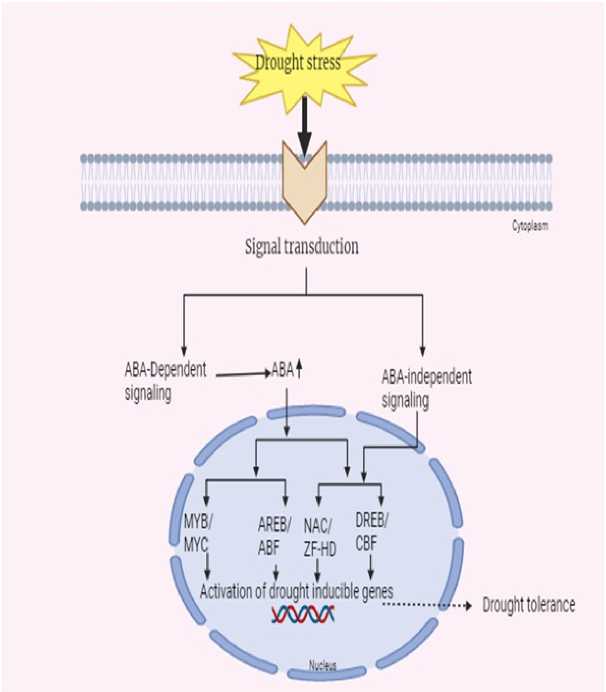
Figure 4. Molecular mechanism of drought tolerance in plants.
Table 1. Key secondary metabolites in medicinal plants.
|
S. o. |
Secondary metabolite |
Plant |
Medicinal properties |
Category |
References |
|
1. |
Atropine |
Atropa belladonna |
Anticholinergic, antiemetic |
Alkaloid |
Hoet et al., 2004 |
|
2. |
Morphine |
Papaver somniferum |
Pain relief, arcotic |
Alkaloid |
Meine et al ., 2005 |
|
3. |
icotine |
Nicotiana tabacum |
Performance enhancer |
Alkaloid |
Heishman et al ., 2010 |
|
4. |
Taxol |
Taxus brevifolia |
Anticancer treatment |
Alkaloid |
Kachooei et al ., 2014 |
|
5. |
Quercetin |
Many fruits, vegetables, grains |
Anticancer, antiinflammatory |
Flavonoids |
Carrillo-Martinez et al., 2024 |
|
6. |
Azadirachtin |
Azadirachta indica |
Insecticide |
Terpenoids |
Indira et al., 2024 |
|
7. |
Luteolin |
Salvia tomentosa |
Anti-inflammatory, antitumor |
Flavonoids |
Lopez-Lazaro, 2009 |
|
8. |
Vincristine and Vinblastine |
Catharanthus roseus |
Chemotherapeutic agent |
Alkaloid |
Paul et al., 2023 |
|
9. |
Convallatoxin |
Convallaria majalis |
Treat acute and chronic heart failure |
Glycosides |
Yang et al ., 2014 |
|
10. |
Digitoxin |
Digitalis purpurea |
Anticancer |
Cardiac glycoside |
Elbaz et al ., 2012 |
|
11. |
Wedelolactone |
Eclipta alba |
Anticancer |
Coumestan |
Mani et al., 2024 |
|
12. |
Andrographolide |
Andrographis paniculata |
Anticancer |
Diterpenoid lactones |
Mourya et al., 2023 |
Proposed Mechanisms for Drought-Induced Secondary Metabolite Production :
Under drought conditions, plants ensure survival by way of activating opportunity metabolic pathways to address stress. These encompass
-
1. ABA Signaling Pathway: Elevated ABA levels activate transcription factors that modify genes that are involved in secondary metabolite production.
-
2. ROS Signaling: Accumulation of ROS during drought acts as a signal for the synthesis of secondary metabolites, especially those with antioxidant activity (e.g., flavonoids, terpenoids).
-
3. Osmoprotectant Accumulation: Metabolites like proline and trehalose help to maintain osmotic balance and activate pathways leading to secondary metabolite synthesis.
To maintain metabolism plants, employ a whole lot of mechanisms and to survive drought stress. Gene expression modifications, enzyme activation, and the synthesis of protective metabolites are key to such adaptation. The role of secondary metabolites is especially essential, as they assist plants to cope with the physiological and biochemical changes associated with drought. For developing strategies to improve drought tolerance in crops understanding these mechanisms are crucial, ensuring agricultural productivity during drought.
ADAPTIVE PROCESSES OF MEDICINAL
PLANTS UNDER DROUGHT STRESS
Drought is a prime abiotic stressor that affects the growth, productivity, and survival of plants, which includes medicinal species too. Plants have developed a variety of adaptive mechanisms to address drought, consisting of metabolic modifications, physiological adjustments, and genetic variations. Specifically, medicinal plants should not only survive but also maintain their production of bioactive compounds that are crucial for their medicinal value. The metabolic and physiological strategies employed by plants to encounter drought stress and the ability for breeding and genetic improvements in medicinal plants to enhance their resilience and productivity are discussed here.
Metabolic Adjustment to Drought
Drought significantly affects every element of plant metabolism, leading to a decrease in productivity and alterations in chemical composition. To deal with drought, plants go through osmotic adjustment, a process in which plants accumulate osmolytes compounds that assist maintain cellular osmotic balance and guard plant tissues from dehydration.
Key metabolic responses
Proline accumulation: Proline is a major osmolyte that performs more than one role under drought situations. It protects cytoplasmic enzymes, stabilizes membranes, and serves as a nitrogen and carbon reservoir for post-stress restoration. Additionally, proline scavenges free radicals, preventing oxidative harm (Renzetti et al., 2024).
Glutathione: Another vital compound in the antioxidative defense mechanism which enables mitigate the harmful effects of ROS generated during drought stress.
Gamma-Aminobutyric acid (GABA): GABA is produced in response to various stress conditions such as drought. It performs a function in stabilizing cellular metabolism and helps in stress tolerance.
Dehydrins: These proteins are specifically produced in response to dehydration and helps in maintaining cellular integrity under drought. Dehydrins are considered markers of drought stress, aiding in the stabilization of membranes and for protecting proteins. Through these metabolic shifts, medicinal plants adapt to drought stress with the aid of minimizing water loss and protecting cellular systems from damage (Riyazuddin et al., 2022).
Physiological Adaptations to Drought
In addition to metabolic adjustments, plants exhibit numerous physiological responses to drought, which assist them to preserve water and maximize its uptake.
Key physiological responses
Stomatal closure: Plants lessen water loss by closing their stomata, which are responsible for the exchange of gas. However, this also limits CO₂ uptake and decreases photosynthesis. Despite this, medicinal plants can preserve productivity by shifting their metabolism to produce stress-related compounds, including secondary metabolites.
Root system modification: To get entry to deeper water reserves, plants often alter their root structure under drought, by extending root length (Fig. 1) and increasing root density in deeper soil layers.
Alternative photosynthetic pathways: Some plants, particularly those in arid areas, might also switch to more water-efficient photosynthetic pathways, including Crassulacean Acid Metabolism (CAM) or C₄ photosynthesis, enabling them to preserve water at the same time maintaining growth and metabolic activities.
These adaptations make sure that medicinal plants can resist drought while minimizing the effect of dehydration on their overall health and productivity.
Potential for Breeding and Genetic Improvement
Drought stress not only influences the productiveness of medicinal plants but additionally affects the biosynthesis of their bioactive compounds. Given the significance of these compounds for medicinal purposes, improving the drought tolerance of these plants through breeding and genetic processes is vital.
Breeding processes
Molecular markers: D A-based molecular markers are a very important tool for identifying genes associated with drought tolerance in medicinal plants. Marker Assisted choice (MAS) allows the breeders to select plant desirable traits like stress tolerance and superior bioactive compound production, with extra precision.
Quantitative Trait Loci (QTL) mapping: QTL mapping allows for identifying genomic regions related to drought tolerance and secondary metabolite production. This can facilitate the improvement of drought-resistant medicinal plants without compromising their medicinal value.
Transgenic approaches: Genetic engineering offers the potential to introduce drought-resistant genes into medicinal plants. For instance, drought-inducible promoters may be used to manipulate the expression of stress tolerance genes, ensuring that they are only activated under drought conditions.
Hybridization with wild relatives: Crossing medicinal plants with drought-resistant wild relatives can introduce new genetic traits that enhance drought tolerance. This strategy, combined with molecular breeding techniques, offers the ability to develop resilient medicinal plant varieties.
By integrating conventional breeding methods with contemporary genetic tools, it is possible to enhance the drought tolerance of medicinal plants, making sure that their survival and the continued production of bioactive compounds under stress conditions.
IMPLICATION FOR MEDICINAL PLANT
CULTIVATION UNDER DROUGHT
Plants are the source of many medicinally important components which are used in human welfare. There is a need to increase the production of these plant products. The plant can be transformed from wild to agricultural by increasing the products of its secondary metabolism. Harvesting a specific medicinal plant can cause a decrease in the genetic diversity of the medicinal plant and habitat loss. Cultivation of medicinal plants is an interesting and alternative method that has other benefits like easy identification, better conservation, and easily available raw materials.
Drought is typically viewed adversely as a farming issue due to significant reductions in yield. On the reverse, medicinal herbs cultivated in arid regions tend to produce more concentrations of vital compounds than the same species grown in temperate climates. Furthermore, intentional application of drought stress while herbal plant cultivating might lead to an increase in the formation of secondary metabolites; however, due to their simultaneous effects, this enhancement may frequently be paid for, if not overpaid. Agronomic research enhances the cultivation of medicinal plants. It increases the quality, resistance and productivity of the plants.
Herbal medicinal plants have a significant role in human healthcare and are a valuable source of active secondary metabolites. Though the area under herbal medicinal plant cultivation is slowly expanding due to growing demand and awareness for herbal drugs, medicinal plant species used for human health cure cannot take off in any significant way at this time due to the low content of specific molecules in plant tissue, overexploitation of naturally occurring plants, and practical excretion in the cultivation of these plants.
umerous agronomic and non-agronomic tactics as part of the management approach to tackle this worldwide issue. To create more resistant crop varieties, breeding with contemporary techniques like targeted gene editing, genomic selection, and marker-assisted selection has been suggested as a viable strategy. Additionally, some methods for enhancing crop resistance to drought through nutrient management including chemical fertilizer treatment were suggested. It was also mentioned that mycorrhizae and growth-promoting bacteria are useful tools for helping crops become drought-tolerant. Overall, it is concluded that creating crops resistant to drought needs the application of a variety of strategies, including advanced breeding instruments and both agronomic and non-agronomic techniques.
CONCLUSION
Drought is one of the most significant abiotic stressors, adversely affecting plant growth, productivity, and the synthesis of secondary metabolites compounds which can be crucial for the medicinal value of flora. To address such stress, medicinal plants adopt a number of metabolic and physiological processes, together with the accumulation of osmolytes, the production of stress-protective proteins, and modifications to root structure and stomatal function. These adaptive responses assist plants survive and maintain productivity during water shortages and at the same time maintaining their bioactive compound production.
The physiological and biochemical responses to drought, such as osmotic adjustments, stomatal regulation, and the activation of antioxidant defense mechanisms, are important not only for plant survival and sustaining secondary metabolite production. Secondary metabolites play a pivotal role in the healing properties of medicinal plants, making it essential to apprehend how drought affects their synthesis. Deeper information on those mechanisms can result in the development of superior breeding strategies that increase drought tolerance in medicinal plants, ensuring both sustainability and the preservation of their medicinal properties.
Future research ought to increase focus on the genetic and molecular mechanisms underlying drought tolerance, especially concerning secondary metabolite biosynthesis pathways. Advances in breeding techniques, such as molecular markers, QTL mapping, and transgenic technology, offer promising opportunities to enhance the resilience of medicinal plants to drought conditions. By incorporating these tools, it is possible to develop drought-resistant plant varieties that not only survive in drought environments but also hold their valuable bioactive compounds, making sure they endure use in both traditional and modern medicine. These insights will be useful for contributing to ecological conservation efforts and the pharmaceutical industry, making medicinal plant cultivation extra sustainable in the face of climate change.
CONFLICTS OF INTEREST
The authors declare that they have no potential conflicts of interest.
