Ретроспективный анализ филогенеза при формировании маточной субпопуляции голштинизированного молочного скота Красноярского края

Автор: Еремина Ирина Юрьевна, Герасимова Людмила Анатольевна, Лущенко Анатолий Егорович
Журнал: Вестник Омского государственного аграрного университета @vestnik-omgau
Рубрика: Биологические науки
Статья в выпуске: 2 (22), 2016 года.
Бесплатный доступ
РЕТРОСПЕКТИВНЫЙ АНАЛИЗ ФИЛОГЕНЕЗА В ФОРМИРОВАНИИ ПОДАВЛЕНИЙ СУБПОПУЛЯЦИЙ МОЛОЧНОГО СКОТА ХОЛШТЕЙНА В КРАСНОЯРСКОМ КРАЙ
Филогенез, генофонд, иммуногенетический мониторинг, популяция, селекция, дифференциация
Короткий адрес: https://sciup.org/142199409
IDR: 142199409 | УДК: 575.827.5.
The phylogenesis retrospective analysis in the formation of the breeding subpopulations of the holstein dairy cattle in Krasnoyarsk krai
THE PHYLOGENESIS RETROSPECTIVE ANALYSIS IN THE FORMATION OF THE BREEDING SUBPOPULATIONS OF THE HOLSTEIN DAIRY CATTLE IN KRASNOYARSK KRAI
Текст научной статьи Ретроспективный анализ филогенеза при формировании маточной субпопуляции голштинизированного молочного скота Красноярского края
Изучение филогенеза пород и составляющих их популяций (стад, линий, семейств) имеет как теоретическое, так и практическое значение. Разработка методов корректировки селекционного процесса с использованием достижений различных отраслей науки, в том числе иммуногенетики, в настоящее время очень актуальна. Процесс формирования генофонда стад и пород очень противоречивый и сложный, во многом не укладывается в рамки принятых понятий и концепций. На практике породообразовательный процесс основан на различных вариантах и методах скрещивания с последующим закреплением желательных генотипов животных, при этом установление сходства потомков с родоначальником базируется чаще всего на эмпирических расчетах. Однако традиционные критерии не всегда в достаточной мере позволяют выявить родство и происхождение пород и отдельных популяционных групп. Консолидация стад и пород в результате крупномасштабной селекции требует усиленного контроля. Наряду с традиционными методами сравнительного анализа популяций, основанного на данных генеалогии, истории и морфологии, иммуногенетический метод способен помочь в этом процессе.
Разработка методов корректировки селекционного процесса с использованием достижений различных отраслей науки, в том числе иммуногенетики, в настоящее время очень актуальна. Выявление генетических особенностей селекционного материала, степени консолидиро-ванности пород и популяций, их дифференциации в процессе селекции – вот возможные направления изучения генофонда иммуногенетическими методами. В последнее время вопрос изучения генетического сходства и его эволюционной истории решается с помощью более тонких механизмов генетической дифференциации, в частности используется метод вычисления генетической дистанции. В случае использования в качестве маркеров антигенов групп крови для сравнения популяции более правомерно анализировать антигенное сходстве популяций.
Селекция молочных животных невозможна без двух противоположных и взаимодополняющих методов: селекции в пределах чистопородных популяций как средства генетического совершенствования и межпородного скрещивания как средства увеличения производства животноводческой продукции, в частности молока. Вопрос об использовании гетерозиса при скрещивании в молочном скотоводстве для получения высокопродуктивных животных уже давно волнует селекционеров. Однако и до сего дня нет исчерпывающей информации о сочетаемости пород, отсутствуют оптимизированные схемы и условия скрещивания, в которых наиболее полно проявился потенциал продуктивности полученных помесей.
Формирование популяций сибирского скота происходило на основе великорусского, завезенного переселенцами из Центральной России. Сибирский скот был распространен на обширной территории Сибири, Бурятии и Дальнего Востока. Поголовье в 40-е годы составляло 7 млн голов. Сибирский скот отличался рядом особенностей: небольшая масса коров – 261–294 кг, низкий удой – 980 кг, высокая жирномолочность – 4–5 %, скороспелость, крепкое здоровье, устойчивость к заболеваниям. Проводимый в СССР с 30-х годов план породного районирования, главной задачей которого было преобразование местного малопродуктивного скота в высокопродуктивный, не учитывал генетический потенциал аборигенных пород, возможность сохранения небольших локальных популяций чистопородных животных.
К началу 50-х годов фактически сибирский скот прекратил существование в результате скрещивания с симментальской и черно-пестрой породами. Исчезновение популяций местного сибирского скота повлекло за собой потерю уникальных генных комплексов, в которых содержалась информация об адаптивных способностях животных к климатическим условиям Сибири, о высоком содержании белка и жира в молоке, резистентности ко многим заболеваниям.
Создание нового сибирского типа черного и красного корня осуществлялось с использованием голштинской породы, обладающей, как известно, самым высоким в мире генетическим потенциалом молочной продуктивности. В настоящее время в крае проводится большая работа по совершенствованию скота, более приспособленного к конкретным природным и технологическим условиям; разработана и осуществляется стратегическая программа работы с породой и линиями.
В генофондах многих пород отмечена тенденция к сокращению генетического разнообразия по группам крови [1]. Определяющим фактором, оказывающим влияние на эти процессы, является резкое уменьшение числа быков-производителей для искусственного осеменения.
Вследствие этого большинство наследственных факторов коров в новых поколениях элиминируются. Каждая популяция животных имеет свою генетическую структуру фенотипов, генотипов и частот полиморфных систем, на которую оказывает влияние направление отбора, состав производителей, различие в плодовитости и жизнеспособности отдельных генотипов, дрейф генов, естественный и презиготический отбор. Интегральное воздействие этих факторов ведет к поддержанию относительной стабильности частот основных аллелей групп крови при неуклонном сокращении генетического разнообразия за счет элиминации редких аллелей. Сохранение стабильного соотношения частот основных аллелей в течение длительного периода, несмотря на различные селекционные воздействия, дает основание полагать, что основные аллели со значительно большей частотой маркируют наиболее «удачные» блоки генов, сохранение которых для популяции предпочтительнее [2].
Считается, что потеря хотя бы одного аллеля группы крови в стаде должна рассматриваться как неблагоприятное явление, исходя из того, что снижение разнообразия полиморфных форм связано с нарушением адаптационных свойств, особенно при возникновении стрессовых ситуаций. В конкретных условиях племенной работы селективное преимущество приобретают животные только определенных генотипов, что обусловливает их лучшую выживаемость и, как следствие этого, увеличение концентрации данных антиген-комбинаций.
Таким образом, генетические маркеры, являясь составной частью генофонда популяций, дают ценную информацию об изменениях, происходящих в них в процессе селекции, которую можно использовать в практике племенной работы для повышения ее эффективности.
Целями исследования было изучение микроэволюционных процессов при формировании генофонда молочного скота в Красноярском крае и последующие рекомендации для корректировки селекционного процесса. В задачи входили: 1) характеристика уровня генетической дифференциации линий отдельных стад по иммуногенетическим показателям; 2) оценка генетической консолидации красно-пестрой и черно-пестрой популяций молочного скота; 3) сравнение методических подходов, применяемых в ходе анализа микроэволюционных процессов.
Объекты и методы
Исходным материалом для проведения исследования были результаты тестирования образцов крови животных, полученные в лаборатории иммуногенетики при кафедре генетики и биотехнологии Красноярского государственного аграрного университета в рамках научноисследовательской работы по теме: «Система эколого-генетического контроля в племенном молочном скотоводстве». Тестировали набором из 47 основных реагентов по девяти генетическим систем: А, B, C, F-V, M, L, J, S, Z. В дальнейшем результаты тестирования, зафиксированные в гемолитических тестах, подвергались генетико-математическому анализу, который традиционно используется для оценки состояния генетических систем в популяции, породе или селекционной группе животных.
Всего в анализе использованы данные о 15000 животных из девяти маточных популяций и быков-производителей племобъединения «Красноярское». Среди них популяции чернопестрого скота (племсовхоз «Сибирь», совхоз «Маяк», племзавод «Элита», учебное хозяйство «Миндерлинское») и популяции красно-пестрого скота (племсовхоз «Красный Маяк», птице-совхоз «Канский», племсовхоз им. Дзержинского, племсовхоз «Абанский», племзавод «Бородинский»). Изучали иммуногенетические характеристики групп крови животных линий чернопестрой и симментальской исходных пород и голштинизированных помесей F1 и F2 соответствующих линий (табл. 1).
Частоты антигенов выражали в процентах; сопоставление генетической структуры популяции по группам крови проводились по общепринятым показателям: индексу генетического (антигенного) сходства r ± m (Серебровский А.С., 1970; Животовский Д.С., 1979); мере генетического сходства (Ней М., 1981). Показатели иммуногенетических дистанций рассчитывали в соответствии с рекомендациями [3, 4].
Поскольку связь между мерой сходства и мерой различия можно записать как F = 1 – K, где F – мера различия; K – мера сходства, все свойства мер сходства можно экстраполировать на соответствующие им меры различия с помощью простого преобразования, и наоборот. При том, что расстояния используются намного чаще, чем меры сходства, их чаще реализуют в статистических программах [5].
Для изучения филогенеза использован метод кластерного анализа иммуногенетических дистанций. В дальнейшем обработка данных проводилась с использованием программы «Statistica». При кластеризации применяли иерархический подход, результаты группировки представлялись в виде дерева группировки (дендрограммы, которые строили методом невзве-шанной, попарной кластеризации).
Таблица 1
Схема анализа породной и линейной дифференциации молочного скота в Красноярском крае
|
Черно-пестрые |
Дифференциация на уровне: |
Красно-пестрые |
|
|
Породы: |
|||
|
Черно-пестрая, голштинская F 1 черно-пестрая |
Симментальская, голштинская F1 красно-пестрая |
||
|
Линии: |
|||
|
Гектора, АнасАдема (АА) КлайнеАдема (КА) |
Мергеля 2122 40-266 Клевера 68 КЕС-4 Синала 4863 ЧС-239 Фасадника 842 ЦС-9 |
||
|
ПС «Сибирь» |
Линии: ВисБекАйдиал 1013415 (ВБА) СилингТрайджунРокит 252803 (СТР) МонтвикЧифтейн 95679 (МЧ) РефлекшнСоверинг 198998 (PC) РозейфСитейшн 267150 (РзС) |
ПС «Красный Маяк» |
|
|
Сз «Маяк» |
ПтСз «Канский» |
||
|
Учхоз «Миндерлинское» |
ПС им. Дзержинского |
||
|
ПЗ «Элита» |
ПС «Абанский» |
||
|
ПО «Краснояское» |
ПЗ «Бородинский» |
||
|
ПО «Красноярское» |
|||
Проведено тестирование по типам крови крупного рогатого скота хозяйств, занимающихся разведением молочного скота. Предварительная фоновая паспортизация поголовья с анализом проведена в 1987–1989 гг. и в дальнейшем проводилась в 1992–1993 гг. по этапам до 2015 г. [6].
Результаты исследований
Генезис формируемых породных особенностей молочного скота Красноярского края представлен красным и черным корнями.
Голштинизированный красно-пестрый скот формировался на базе симментальской породы. Генофонд отдельных стад пополнялся также красно-пестрыми животными высокой кровности, импортируемыми из европейских стран (Германия, Дания).
Сравнительный анализ скота симментальской породы, сохранившегося в разных популяциях Красноярского края, выявляет как общие черты, присущие данной породе в регионе, так и специфические, определяющиеся различием экологических условий для каждой популяции.
Сравнительный анализ генофондов объединенных симментальских линий фиксирует наличие как общих, так и специфических черт, присущих линиям Фасадника, Сигнала, Клевера и Мергеля. Можно отметить некоторую обособленность от других линии Клевера. Наименьшее сходство определено между линиями Клевера и Мергеля (r = 0,930), а наибольшее – между линиями Сигнала и Фасадника (r = 0,990) (табл. 2). Тем не менее, индексы антигенного сходства довольно высоки и превышают значения, при которых регистрируются существенные отличия между группами по антигенному спектру.
Таблица 2
Коэффициенты генетического сходства (г ± mг) между интегральными линиями животных симментальской породы разных популяций Красноярского края
|
Линии |
Фасадника |
Сигнала |
Мергеля |
Клевера |
|
Фасадника |
||||
|
Сигнала |
0,990 ± 0,001 |
|||
|
Мергеля |
0,996 ± 0,006 |
0,973 ± 0,004 |
||
|
Клевера |
0,948 ± 0,008 |
0,941 ± 0,009 |
0,930 ± 0,013 |
Таким образом, зафиксирована точка начального этапа дивергенции линий, в которой прослеживается общность эколого-генетических корней. Аналогичная ситуация наблюдается и по объединенным голштинским линиям: МЧ, PC и СТР. Такое сходство объясняется использованием одной материнской основы при формировании разных линий. Индексы антигенного сходства фиксируют общность линии РзС с линией МЧ; PC с РзС. Наиболее удалены друг от друга линии PC и СТР (табл. 3).
Таблица 3
Коэффициенты генетического сходства (г ± mг) между интегральными линиями животных голштинской породы красно-пестрых популяций в Красноярском крае
|
Линии |
МонвикЧифтейн |
РозейфСитейшн |
СилингТрайджун Рокит |
Рефлекшн Соверинг |
|
МонвикЧифтейн |
– |
– |
– |
– |
|
РозейфСитейшн |
0,919 ± 0,007 |
– |
– |
– |
|
СилингТрайджунРокит |
0,866 ± 0,015 |
0,893 ± 0,013 |
– |
– |
|
РефлекшнСоверинг |
0,879 ± 0,015 |
0,914 ± 0,010 |
0,864 ± 0,019 |
– |
Было проведено параллельное исследование и сравнение результатов, полученных при вычислении индекса антигенного сходства между линиями каждой изученной популяции, и картины взаимосвязи этих линий, изображенной на дендрограмме.
Самыми консолидированными были закупленные в Германии телки – чистопородные голштины, а наиболее гетерогенными – симментал-голштинские помеси.
Анализ коэффициентов генетического сходства между линиями в популяции крупного рогатого скота племсовхоза «Красный Маяк» подтвердил генетическую близость симментальских линий между собой. Коэффициент колеблется от 0,960 до 0,985. Самыми близкими оказались линии Альберта Резвого, Фасадника и Сигнала (г = 0,985), а самыми удаленными друг от друга линии Сигнала и Радониса (г = 0,959) и линии Рафаэля и Сигнала (г = 0,959). Наиболее отдаленными и от симментальского окота и от других оказались голштинизированные помеси линий РзС и PC – между ними г = 0,915. Это объясняется использованием семени импортных быков при формировании данных линий, имеющих в своих генотипах специфические антигены, редкие для данной популяции [7].
Кластерный анализ линий популяций ПС им. Дзержинского наглядно показал генетическую консолидацию симментальских линий и монбельярдского окота и значительное удаление от них животных линии РзС. Следует отметить высокую степень консолидации всех селекционных групп данного хозяйства (г от 0,915 до 0,977), что говорит об общности генофонда популяции и ее существовании во времени как единого целого. Кроме того, высокие показатели индекса антигенного сходства между различными линиями говорят об отсутствии между ними четких различий, что может быть свидетельством недостаточно высокого уровня селекционного учета при беспорядочном кроссировании линий, или об отсутствии научно обоснованного подхода к подбору быков-производителей, участвующих в формировании стада в данных условиях генетического баланса.
В целом индексы антигенного сходства отражают генетическую близость линий, разводимых в данном хозяйстве, как симментальских, так и F1 помесных. Это объясняется, во-первых, беспорядочными кроссами линий, не позволяющими им оформиться в обособленную, достаточно выраженную структуру, и во-вторых, сильным генетическим потенциалом маточного стада. Его невозможно радикально изменить в F1 даже при введении в популяцию принципиально данных генотипов и тем более без участия генетического контроля процесса ввода этих генотипов, а также подбора наиболее подходящих для данной, конкретно взятой популяций скота.
Индексы антигенного сходства между популяциями подтверждают исключительность ПС им. Дзержинского, имеющего низкий показатель сходства при сравнении с популяциями ПС «Абанский», ПС «Красный Маяк» и ПЗ «Бородинский» (табл. 4). Между популяциями ПС им. Дзержинского и птицесовхоза «Канский» – наивысший из вычисленных индексов антигенного сходства. Примечательно, что эти два хозяйства позже других получили статус племенных и поэтому генофонд этих стад формировался несколько обособленно, что отразилось в процессе экогенеза и на генном уровне отмечается до сих пор.
Кластерный анализ всего комплекса данных по линиям в рассматриваемых хозяйствах подтверждает вывод о консолидированности животных в пределах одной популяции (рис. 1).
Дендрограмма представляет собой совокупность крупных кластеров, состоящих из последовательно соединяющихся ветвей – линий скота, составляющих определенную популяцию, которые затем объединяются в еще более крупные кластеры. Суммируя имеющиеся данные, с учетом накопленных генофондов объединенных линий можно с уверенностью говорить о наличии в настоящее время генетической разобщенности пород, как в пределах одной популяции, так и в их совокупности, о значительной роли материнского генофонда, обеспечивающего консолидацию селекционных групп животных одной популяции, и об уникальности экологоисторически сложившихся генофондов каждой популяции, заключающих в себе адаптационные механизмы к конкретным условиям данного ареала.
Таблица 4
Коэффициенты генетического сходства (г ± mг) между популяциями крупного рогатого окота племенных хозяйств Красноярского края
|
Популяция |
ПС «Канский» |
ПС им. Дзержинского |
ПС «Абанский» |
ПС «Красный Маяк» |
ГПЗ «Бородино» |
|
ПС «Канский» |
– |
– |
– |
– |
– |
|
ПС им. Дзержинского |
0,980 ± 0,002 |
– |
– |
– |
– |
|
ПС «Абанский» |
0,896 ± 0,009 |
0,823 ± 0,018 |
– |
– |
– |
|
ПС «Красный Маяк» |
0,948 ± 0,004 |
0,862 ± 0,012 |
0,931 ± 0,007 |
– |
– |
|
ГПЗ «Бородино» |
0,957 ± 0,003 |
0,890 ± 0,009 |
0,913 ± 0,008 |
0,962 ± 0,003 |
– |
Голштинизированный черно-пестрый скот формировался на базе черно-пестрой породы. Вкратце проанализируем особенности отдельных стад.
Племзавод «Элита» характеризуется сравнительно высокой консолидированностью стада и незначительной межлинейной дифференциацией, наибольшая разнородность между помесями линий черно-пестрой породы: КлайнеАдема и Гектора (r = 0,875), в отличие от межпородных помесей.
Стадо совхоза «Сибирь» значительно разнородное по своей генеалогической структуре, демонстрирует генетическую разобщенность единичных представителей базовой симментальской породы и межпородных помесей линий АннасАдема и ПабстГо-вернер. Среди помесей черно-пестрой и голштинской пород следует отметить минимальные значения показателей при наличии кровности по линии ПабстГовернер. Генетическое сходство отмечается между помесями голштинов F1 и F2 на основе чернопестрой линии АннасАдема.
ПЗ "Бор." симм.
ПЗ "Бор." Ғ1
ПЗ “Бор." Герм.
Сз "Кр. Маяк" Ғ1
Сз "Кр. Маяк" симм.
ПС "Абанский" Ғ1
ПС им "Дзерж." Ғ1+симм.
ПтСз "Канский"
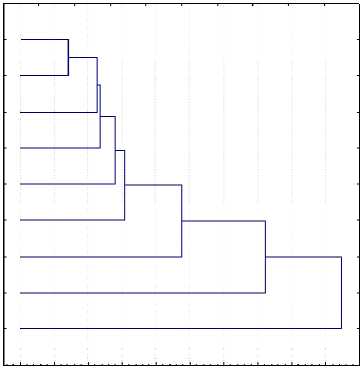
Расстояние
Рис. 1. Дендрограмма микрофилогенеза красно-пестрого голштинизированного корня молочного скота Красноярского края
О 10 20 30 40 50 60 70 80 90 100
Внутристадная дифференциация крупного рогатого скота стада совхоза «Маяк» весьма специфична. Наименьшее сходство отмечается между родственными группами РозейфСи-тейшн и РефлекшнСоверинг (r = 0,621). Невысок этот показатель и для представителей симментальской породы и черно-пестрой линии КлайнеАдема (r = 0,690). На основании проведенного анализа генетической дифференциации следует внести уточнение в генеалогическую структуру изучаемых групп помесей:
-
1. В период обследования стадо отделения «Седельниково» было представлено в большей степени помесями голштинской породы с симментальской, чем с черно-пестрой породой.
-
2. Субпопуляции отделений «Высотино» и «Кекур» имели в генеалогической основе линию АннасАдема, а стадо отделения «Абакшино» – КлайнеАдема.
Обновление черно-пестрого стада учебного хозяйства «Миндерлинское» происходило за счет проведения межпородного скрещивания с голштинскими быками (искусственное осеменение) и ввода в стадо помесей молочного скота различного экогенеза. Генетическая разобщенность наблюдается между группами сахалинских животных и местными коровами шилинской субпопуляции (r = 0,436), группами местного скота и шилинского отделения (r = 0,494), потомками голштинских линий и местным скотом (r = 0,579–0,599). Наибольшее сходство отмечается между черно-пестрой линией АннасАдема и линией ВисБекАйдиал (r = 0,990). Близки показатели генетического сходства между линиями ВисБекАйдиал, РефлекшнСоверинг и молочным скотом ГДР.
Обобщенное представление о состоянии переходного полиморфизма в период проведения скрещивания черно-пестрого и голштинофризского скота дает объединенный кластерный анализ исследуемых групп помесей в двух хозяйствах (рис. 2).
Многоступенчатость дендрограммы, выделение объединенных кластеров первого уровня преимущественно по признаку принадлежности к
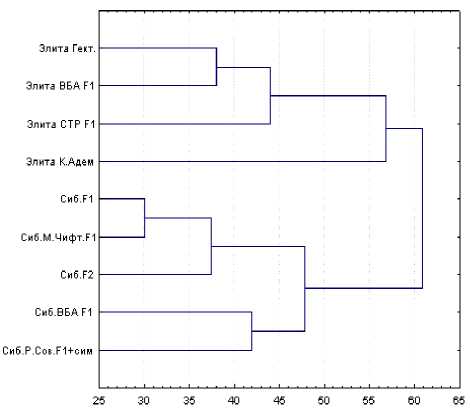
Рис. 2. Дендрограмма микрофилогении отдельных стад черно-пестрого скота Красноярского края
одному стаду и одновременно наметившаяся линейная дифференциация (наличие объединенных кластеров) указывают на фазу переходного полиморфизма. Метод разведения, в основе которого лежит регулярная сменяемость линий на массиве маточного поголовья, оказывает свое влияние на процессы, протекающие в популяции под влиянием межпородного скрещивания. Следствие этого влияния – низкая консолидиро-ванность генетического материала, выразившаяся в незначительных генетических дистанциях между помесями различных линий голштинского скота внутри каждого из стад.
В целом такое распределение стад вполне удовлетворительно объясняет их реальные взаимосвязи, известные из генеалогии и истории формирования, что свидетельствует о больших возможностях применения иммуногенетических маркеров в исследованиях по эволюционной истории.
Обобщенное представление о степени консолидированности объединенных групп красно-пестрого, черно-пестрого голштинизированного скота F1, F2 дает материал табл. 5. Межпородные коэффициенты генетического сходства высокие и сигнализируют о значительном генетическом сходстве. Так, объединенная группа симментальской породы близка к объединенной группе красно-пестрых голштинов F1, F2 (г = 0,954). А различия между объединенными группами красно-пестрого, черно-пестрого голштинизированного скота F1, F2 несколько выше (г = 0,897).
Наименьшее сходство со всеми группами отмечено у скота, завезенного из ФРГ, и у красно-пестрых помесей птицесовхоза «Канский» (r от 0,709 до 0,853). Все это указывают на слабую дифференцированность исходного селекционного материала [9] и фазу переходного полиморфизма.
В дальнейшем были предприняты попытки использовать разные программные продукты (PHYLIP, MEGA) для анализа имеющихся иммуногенетических баз данных. Однако данные программы предназначены, в первую очередь, для работы с последовательностями нуклеотидов ДНК или аминокислот белков. Кроме того, длительность предварительной подготовки первичных данных и не всегда понятного интерфейса программы усложнили поставленную задачу. Поэтому были использованы возможности программы «Statistica», алгоритмы которой предусматривают кластеризацию на основе расстояний и построение дендрограмм.
Таблица 5
Коэффициенты генетического сходства (г ± mг) между субпопуляциями и объединенными группами крупного рогатого скота Красноярского края
|
Порода |
Кр. п. голшт. ФРГ |
Кр. п. голшт. Канск |
Кр. п. голшт. F1, F2 |
Симм. |
Симм. быки |
Чр. п. голшт. Элита |
Чр. п. голшт. F1, F2 |
Чр. п. быки |
|
Кр. п. голшт. ФРГ |
– |
– |
– |
– |
– |
– |
– |
– |
|
Кр. п. голшт. Канск |
0,709 ± 0,002 |
– |
– |
– |
– |
– |
– |
– |
|
Кр. п. голшт. F1, F2 |
0,853 ± 0,002 |
0,798 ± 0,005 |
– |
– |
– |
– |
– |
– |
|
Симм. |
0,862 ± 0,001 |
0,784 ± 0,002 |
0,954 ± 0,007 |
– |
– |
– |
– |
– |
|
Симм. быки |
0,742 ± 0,004 |
0,728 ± 0,004 |
0,822 ± 0,005 |
0,806 ± 0,005 |
– |
– |
– |
– |
|
Чр. п. голшт. Элита |
0,788 ± 0,003 |
0,788 ± 0,006 |
0,882 ± 0,007 |
0,889 ± 0,004 |
0,727 ± 0,007 |
– |
– |
– |
|
Чр. пестр. голшт. F1, F2 |
0,835 ± 0,003 |
0,809 ± 0,005 |
0,897 ± 0,008 |
0,899 ± 0,001 |
0,807 ± 0,007 |
0,887 ± 0,005 |
– |
– |
|
Чр. п. быки |
0,793 ± 0,002 |
0,817 ± 0,004 |
0,861 ± 0,008 |
0,850 ± 0,005 |
0,832 ± 0,004 |
0,838 ± 0,006 |
0,856 ± 0,003 |
– |
Сопоставляя полученные результаты, следует отметить, что дендрограммы на рис. 3 адекватно отражают процессы, описанные при анализе показателей генетического сходства.
Заключение
Таким образом, микроэволюционные процессы на фоне голштинизации характеризуются определенной направленностью. В целом по исследуемым популяциям линейной консолидации не выявлено. Линейная дифференциация существует на внутристадном уровне в частности. Особенности филогенеза изученных популяций нашли свое отражение в дендрограммах, каждая из которых показывает ситуацию в конкретном стаде на момент исследования. Объединенный кластерный анализ всех исследуемых групп указывает на переходную стадию их генетической дивергенции. Кластерный анализ наглядно отразил уникальность каждой популяции, являющуюся результатом стратегии племенной работы.
Внутрипопуляционные межлинейные различия недопустимо малы и не оправдывают прогнозируемый селекционный ответ. Межпопуляционные генетические дистанции незначительны. Каждая популяция по результатам кластерного анализа обнаруживает внутреннюю консолидацию. На уровне породы и популяционном уровне выявлен эколого-генетический полиморфизм, обусловленный локализацией каждой отдельной популяции.
Результаты изучения особенностей спектра антигенов одноименных линейных групп в разных стадах указывают на начавшийся процесс линейной дифференциации внутри каждого конкретного хозяйства; выявлены маркерные антигены. Следует подчеркнуть, что межлинейные различия внутри этих двух стад, выявленные у представителей межпородных помесей F1, указывают на наличие определенного генетического полиморфизма. Внутрипопуляционные межлинейные различия недопустимо малы и не оправдывают прогнозируемый селекционный ответ.
Следует отметить, что проведенное исследование установило возможность использования ряда показателей, а именно коэффициента антигенного сходства, показателя иммуногене-тической дистанции ( d ) и меры генетического сходства ( I ) для оценки меры меж- и внутрипо-родной дифференциации. Приведенный метод является одним из методов молекулярной таксономии и представляет интерес с точки зрения совершенствования методов селекции животных.
Результаты исследований могут быть привлечены, в частности, для прогнозирования гетерозиса, дифференциации линий и др.
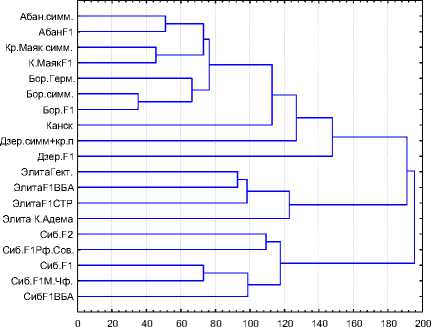
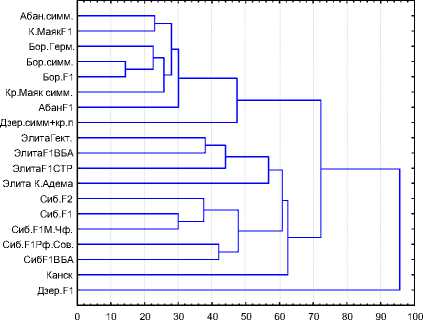
Рис. 3. Дендрограмма, отражающая эвклидову дистанцию (слева) и дистанцию по Чебышеву (справа)
Список литературы Ретроспективный анализ филогенеза при формировании маточной субпопуляции голштинизированного молочного скота Красноярского края
- Алтухов, Ю.П. Генетические процессы в популяциях/Ю.П. Алтухов. -М., 2003. -372 с.
- Сухова, Л.Г. Иммуногенетическая характеристика линий в племзаводе «Тагил» по В-и С-аллелям групп крови/Л.Г. Сухова//Труды Урал. НИИСХ. -Вып. 52. -1988. -С. 62-65.
- Животовский, Л.А. Популяционная биометрия/Л.А. Животовский. -М.: Наука, 1991 -272 с.
- Животовский, Л.А. Методические рекомендации по использованию статистического анализа иммуногенетических данных в селекции животных/Л.А. Животовский, А.М. Машуров; ВНИИ животноводства. -Дубровицы, 1987. -24 с.
- Шрейдер, Ю.А. Что такое расстояние?/Ю.А. Шрейдер. -М.: Физматлит, 1963. -76 с. l
- Еремина, И.Ю. Иммуногенетический анализ базового генофонда скота в ОАО «Племзавод Красный Маяк»/И.Ю. Еремина, Л.А. Герасимова//Вестник КрасГАУ. -2015. -№ 3. -С. 106-110.
- Жукова, Н.М. Возможности повышения эффективности использования производителей голштинской породы/Н.М. Жукова, А.И. Хрунова//Генетические методы в селекции сельскохозяйственных животных. -М.: ВНИИплем, 1990. -С. 64-66.
- Романова, Е.М. Междисциплинарная концепция экологогенетического мониторинга крупномасштабных селекционных программ: автореф. дис. … д-ра биол. наук/Е.М. Романова. -Красноярск, 1994. -32 с.
- Машуров, A.M. Учитывать генетические дистанции между породами при селекции/A.M. Maшуров, В.И. Черкащенко//Зоотехния. -1987. -№ 2. -С. 21-23.
