Результаты и перспективы биоиндикационных исследований водоемов и водотоков Волжского бассейна (на примере хирономид, Diptera: Chironomidae)
")
Автор: Зинченко Т.Д.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1 т.8, 2006 года.
Бесплатный доступ
Приводятся обобщенные результаты многолетних исследований хирономид на популяционном (факториальная экология), биоценотическом (структурно-функциональные характеристики сообществ) и экосистемном (многолетняя динамика) уровнях в биоиндикационных исследованиях оценки экологического состояния разнотипных водных объектов Волжского бассейна при разных типах природного и антропогенного воздействия. Впервые изучена хирономидофауна водоемов и водотоков бассейна Верхней (Учинский канал, бассейн р. Колокша), Средней и Нижней Волги (более 40 малых и средних рек). В лабораторных и естественных условиях установлено значение хирономид как биопомех на очистных сооруженх. Разработанный автором аутэкологический спектр ("экологические шкалы") хирономид и карты их распределения позволили создать информационную основу для формирования экологических прогнозов состояния рек, обоснования региональных критериев биологической оценки качества поверхностных вод при мониторинге экологического состояния водотоков. Полученные результаты позволили оценить динамику хирономидофауны и роль хирономид как индикаторов при проведении крупномасштабного мониторинга водоемов Волжского бассейна (Куйбышевское водохранилище и Каспийского моря).
Короткий адрес: https://sciup.org/148197805
IDR: 148197805 | УДК: 574.5+588+595.771
Results and prospects of bioindication researches of the reservoirs and channels in the Volga river basin (by the example of Chironomids, Diptera: Chironomidae)
The integrated results of long-term researches of chironomids at population (factorial ecology), biocoenosis (structurally functional characteristics of communities) and ecosystem (long-term dynamics) levels in bioindication researches of the ecological state assessment of multi-type fresh-waters in the Volga River Basin are given at different types of natural and anthropogenic impact. Chironomids fauna of the Upper Volga (Ucha channel, the Koloksha river basin), the Middle and Lower Volga (more than 40 stream and middle rivers) fresh waters is investigated for the first time. The importance of chironomids as biohindrances in the water supply plants is developed in experimental and natural conditions. The developed autecological spectrum of chironomids (ecological scales) and their distributional maps led to the creation of informative base for ecological prediction development of the rivers state, foundation of the regional criteria of the biological quality assessment in fresh waters during their monitoring. The received results allowed estimating chironomids fauna dynamics and chironomids role as indicators at conduction of large-scale monitoring reservoirs of the Volga river basin (Kuibyshev river basin, the Caspian Sea).
Текст научной статьи Результаты и перспективы биоиндикационных исследований водоемов и водотоков Волжского бассейна (на примере хирономид, Diptera: Chironomidae)
Биоиндикационные исследования в водоемах на разных уровнях организации жизни являются интегрирующим фактором изучения водных экосистем. Не вызывает сомнений, что биоиндикационные исследования в рамках гидроэкологического мониторинга должны носить системный характер и быть одной из приоритетных областей водной экологии.
Наблюдаемый подъем в области изучения экологического состояния водоемов и водотоков методами биоиндикации с использованием донных сообществ связан с необходимостью решения практических задач, касающихся защиты и сохранения экологических систем континентальных водоемов и в значительной степени обусловлен успехами развития действенной теории функционирования экологических систем [4]. Однако реализация теоретических предпосылок функционирования при изучении проточных гидроэкосистем до настоящего времени затруднена в связи с отсутствием надежных сведений по экологии отдельных видов и групп животных, которые могут оказывать значительное влияние на функциональные характеристи- ки надорганизменных систем [4, 5, 10, 29]. Среди амфибиотических насекомых хироно-мидам отводится решающая роль как индикаторам экологического состояния континентальных водоемов [10, 36, 40, 43, 47-49, 50, 51]. В связи с выше изложенным была проведена обобщенная оценка хирономид (Chironomidae) как объекта биоиндикации в водотоках и водоемах бассейна Волги при разных типах природного и антропогенного воздействия. Исследования проводились на модельных водотоках, характерных для экосистем разнотипных водных объектов (Учинский канал, р. Чапаевка). Оценивалась роль хирономид как индикаторов при проведении крупномасшабного мониторинга водоемов Волжского бассейна (процессы эвтрофирова-ния в Куйбышевском водохранилище) и в условиях гидрологических изменений в Каспийском море.
Материал и методы исследований
Материал включает многолетние сборы бентоса и хирономид из водоемов и водотоков Волжского бассейна: 43 рек и 10 озер Самарской области, 5 рек Саратовской области,
7 рек Владимирской области, Куйбышевского водохранилища, водотоков и водоемов из низовий дельты р. Волги (1981-1985, 1993 и 1995 гг.) и Каспийского моря (1981-1984 гг.), приведены оригинальные данные индикационных исследований хирономид из обрастаний Учинского водопроводного канала и очистных сооружений водопроводной станции в Московской области (1977-1980 гг.).
Гидрографические характеристики и индексы загрязнения воды (ИЗВ) водотоков представлены в табл. 1. Низкое качество воды отдельных рек (Чапаевка – в черте крупных промышленных центров г. Чапаевск) обусловлено процессами ускоренной эвтрофикации и токсификации. Большинство малых склоновых рек лесостепной зоны Волжского бассейна относятся к III и IV классам качества («умеренно загрязненные» и «загрязненные»). Воды II класса качества характерны для притоков р. Колокша (Владимирская область), р. Байтуган (Самарская область) и др. Учинский водопроводный канал и р. Чапаевка были выбраны в качестве модельных водотоков, испытывающих разную антропогенную нагрузку.
Разработанная информационная система [44] использовалась для анализа речных экосистем (более 40 рек разного типа и уровня антропогенной нагрузки, расположенных в лесостепной и степной зонах Среднего и Нижнего Поволжья) в их естественном состоянии и под воздействием загрязнения, а также структурно-функциональной оценки гидробионтов в экосистемах рек разного типа.
Для изучения влияния комплекса антропогенных факторов на развитие сообществ бентоса в рамках программы экологического мониторинга малых рек бассейна Средней и Нижней Волги были проведены специальные исследования р. Чапаевка в условиях критических уровней загрязнения (1990-1995 гг.). Методика отбора и обработки проб бентоса и обрастаний, изучение хирономид были стандартными [34, 38, 43], подробно изложены ранее [19, 20]. В отсутствии антропогенного загрязнения были исследованы очистные сооружения в системе: Учинское водохранилище ⇒ Учинский водопроводный канал (включает открытые водоводы и зак- рытые трубопроводы) ⇒ «ковш» (открытый отстойник с первичным хлорированием воды) ⇒ очистные сооружения водопроводной станции. Применялись разные методы: экспериментальные искусственные субстраты, отлов имаго, выведение хирономид из кладок и личинок, отбор бентосных проб и обрастаний, постановка ловушек для вылета имаго, изучение питания массовых видов [52].
Данные по хирономидофауне основываются на материале, полученном более чем из 500 количественных и 300 качественных образцов бентоса из рек, более 100 – из озер и более 300 количественных проб бентоса из Куйбышевского водохранилища [17, 53], более 150 образцов бентоса из водоемов дельты Волги и 105 образцов бентоса из Каспийского моря [21].
Были использованы различные гидрохимические и гидрологические параметры, а также показатели концентрации поллютантов в воде – скорость течения, глубина, t о С, pH, O 2 , P общ , БО, БПК 5 , содержание летучих фенолов, нефтепродуктов и др.
Химический анализ воды осуществлялся в аккредитованных гидрохимических лабораториях Института экологии Волжского бассейна РАН, «Центра мониторинга водной и геологической среды» (г. Самара), в производственной лаборатории СП «Водоканал» и в центре Госсанэпиднадзора (г. Тольятти).
Обработка исходной информации проводилась стандартными методами математического анализа.
При оценке состояния речных экосистем использовались следующие принципы:
-
- сезонная сопоставимость результатов (для рек Самарской области информационным периодом являются июнь-июль, характеризующиеся наибольшим видовым разнообразием);
-
- единовременность отбора проб (в реке от ее истока до устья);
-
- равномерность распределения и репрезентативность выборки для характеристики водотоков разных ландшафтных зон и рек разного типа (при обследовании рек Самарской области были по возможности охвачены все ландшафтно-географические зоны и
- реки разного размера и типа);
-
- сопоставимость полученных результатов при использовании оценок качества воды по химическим и биологическим показателям.
Результаты и обсуждение
Рассматриваются биоиндикационные особенности функционирования хирономид при разнотипном антропогенном воздействии на различные водные системы бассейна Верхней, Средней и Нижней Волги.
Водотоки Верхней Волги (Учинский водопроводный канал) . На примере детального изучения сообществ зоообрастаний Учинского водопроводного канала показано значение хирономид как биоиндикаторов с учетом анализа популяционных показателей и индивидуальной изменчивости доминирующих в обрастаниях хирономид [15, 16, 19, 22].
Хирономиды обрастаний водопроводного канала представляют ведущую группу беспозвоночных, которые, проходя с потоком воды на очистные сооружения, создают помехи в водоснабжении.
Разнообразие хирономидофауны и состав животных разных подсемейств свидетельствуют о широком экологическом диапазоне обитания животных, которые развиваются в разнообразных и специфических условиях водоводов очистных сооружений. Доминирующей группой по числу видов и количественной представленности являются хиро-номиды подсем. Orthocladiinae, из которых Cricotopus bicinctus (Mg.), Orthocladius oblidens (Walk.), Synortocladius semivirens (K.) составляют около 80% от среднесезонной численности хирономид. Установлено, что из 38 видов хирономид в потоке воды на очистных сооружениях, лишь личинки младших возрастов C. bicinctus и Tanytarsus sp. проходят до последнего этапа очистки воды. При изучении особенностей биологии хироно-мид-обрастателей (жизненные циклы, возрастная структура, условия обитания, питание и пищевое поведение, сезонная динамика), показано, что они демонстрируют различные стратегии приспособления к условиям обитания в канале и на очистных сооружениях [21].
Ценоз хирономид является чувствитель- ным индикатором общего состояния перифи-тонных сообществ канала и отдельных его компонентов, что выражается в их функциональной перестройке, реакции на специфичность условий обитания. Видовое разнообразие хирономид закономерно уменьшается от водоводов с ненарушенной структурой сообществ обрастаний (индекс Шеннона-Уивера Н = 3,52) начального участка магистрального канала к открытым парным водоводам середины канала, где имеет место эффект заселения незанятой экологической ниши (Н = 1,94) и трубопроводам, в которых высокое течение и отсутствие освещенности выступают как факторы отбора определенных видов (Н = 2,10).
Изменения структуры сообществ обрастаний и комплекса хирономид происходят при воздействии стресс-факторов (механическая чистка водоводов, закрытые трубопроводы, хлорирование воды в отстойнике) что выражается в сокращении числа видов, смене до-минантов, уменьшении видового разнообразия, упрощении системы трофических связей и др. Одним из примеров отклика воздействий является изменение характера и степени доминирования хирономид в разных водоводах канала [16,17, 52] .
Оригинальные исследования роли хиро-номид как биологических помех в водопроводном канале и на очистных сооружениях позволили установить закономерности количественного развития и распределения массовых видов хирономид в условно чистой среде и дать оценку биоиндикационных качеств хирономид как биологических помех на очистных сооружениях водопроводной станции при попадании их в водопроводную воду.
Равнинные реки лесостепной и степной зон Средней и Нижней Волги. При трансформации водного режима в условиях зарегулирования стока на всем протяжении р. Чапаевка и подпора ее уровня водами Саратовского водохранилища были изучены особенности структурно-функциональных характеристик реки и их изменений в условиях антропогенного воздействия на водоток.
Отмечается высокий токсический уровень загрязнения воды и донных отложений в
Таблица 1. Гидрографические характеристики и индексы загрязнения воды (ИЗВ) некоторых водотоков Волжского бассейна*
|
_D ci О CD О CO H о Ф У co ^ |
>s h К X с о Ф с d 2 & 2 О |
X X X |
О |
CD X X X |
S’ О X О о ZE CD e |
Q О o^ CD e |
- £ ° O v ч S T ZE О . Ф CL 3 "8" ^ ф ° ° 5? |
ri - csj CD ct 3-0 ^r ° °- ^ ^ Ф z -9 CD ZE |
IO co" О |
CD О |
o" о - о 5 О >,"8 s5 CD -8 CD X |
|
со о СО si го |
— > |
— |
= = |
> |
— > |
= > |
= |
> |
— |
> |
|
|
СО СО X |
CD СО СО |
со~ cd" |
co -^" cxj" |
LO co" |
|
co CXl" exT co" |
CD |
CO cxj" |
LO co~ co" |
co co" |
|
|
о £ 5 I i g a> О ф н |
CD СО со" СО СО со" |
CD СО cd" CD СО cd" |
CD cd" |
CD LO |
CD CO |
LO co co" |
CD CD CD^ co" |
CD co" |
CD °4 |
1— CD X |
|
|
си 2 £ = |
CD |
CO |
CO cxj" |
CO |
CD^ |
UD cxj" |
co~ co" |
co~ co" |
co co" |
||
|
2 го с; СЕ |
СО |
со CXI |
CO |
LO LO |
LO CO |
CD CXI |
О |
co |
CXI CXI |
||
|
-D ” 3 о 2 5-ю « 3 о о о 5 R Ч -П С он |
со |
cd" |
CD LO, CO |
co |
s |
CD LO" |
co co co" |
co" |
|||
|
н = Ф ч s ” ш |
со 2 ОС 5 d |
CL X СГ CD CD О О ZE S |
CD О S О E 3 cr (J D m X CO о >^ >x ЬС |
CL X cr CD CD О О CD О 1— CO d CO О |
CL X СГ CD CD О О CD О 1— CO CL CO О |
CL X СГ CD CD О О CD О 1— CO CL CO О |
co co 2 co О ci. |
о о ci. |
о о ci. |
d X СГ CD CD О О cr co d о о m |
|
|
СО ф О- |
СО 3 о с; о |
c; co ZE ZE s |
ro ZE CO d |
co d co 2 co о |
LO ^ О CD О |J . ° о |
Го - co oo m ^= ^ T CD । CD CXI CO T— t— t— j u (5 (3 |
CD ZE о 3 c; о LQ |
co CD о ZE d CD |
ZE co co LQ |
CO ZE CO CD d CD LQ |
|
органическое вещество (по БПК5,
нижнем течении реки (комбинированное воздействие эвтрофирования и загрязнения ниже г. Чапаевска) и обогащение питательными веществами сельскохозяйственных и бытовых стоков вод в верхнем течении реки. Донные отложения реки, аккумулирующие поллютанты из водной толщи, содержат тяжелые металлы, нефтепродукты, фенолы и пестициды в количествах, превышающих нормативы в десятки и сотни раз, и выступают в роли вторичного источника загрязнения воды р. Чапаевка [13]. Мощный антропогенный пресс привел к эвтрофированию реки, ухудшению экологических условий обитания флоры и фауны как на водосборе, так и в самом водоеме. Приоритетными загрязнителями реки с токсическим эффектом являются нитритный и аммонийный азот, нефтепродукты, фенолы, хлорорганические пестициды, тяжелые металлы, наиболее массовые из которых – медь, марганец, цинк, кадмий. Полученные данные о формировании химического состава отложений под влиянием поступающих в реку загрязняющих веществ позволили оценить техногенную составляющую зарегулированной реки. Состояние донных отложений и устойчивость их структуры определяют состав и функционирование донных сообществ [11, 13, 14].
Изменение структуры сообществ зообентоса в условиях антропогенного загрязнения р. Чапаевка. Проведение индикационных исследований в равнинной реке рассмотрено с позиций того, что водные сообщества и виды животных как в естественных, так и в антрпогенных условиях, являются компонентами экосистемы рек (природных или преобразованных техногенезом), входят в состав гидробиоценоза, изменяющегося в пространстве, развивающегося во времени в условиях критических антропогенных нагрузок.
В составе макрозообентоса в 1990-2004 гг. установлено 259 видов и таксономических групп гидробионтов, из которых ведущими являются хирономиды – 101 вид и личиночная форма. Особенностью макрозообентоса малопроточной р. Чапаевка является преобладание по численности и биомассе олигохет, представителей фитофильной зарослевой фауны и пелофильных хирономид.
Число таксонов верхнего и среднего течения р. Чапаевка превышает более чем в 3-7 раз видовой состав бентоса нижнего течения реки. Структура донных сообществ на сравниваемых участках реки (эвтрофирование – токсификация) различается по числу таксонов, видовому разнообразию, численности и биомассе бентоса. Выше г. Чапаевска, в отсутствии загрязнения промышленными сточными водами массовыми из хирономид в прибрежье и медиали реки являются Chironomus plumosus (L.), Polypedilum nubeculosum (Mg.), Procladius ferrugineus (K.), Tanytarsus spp., Cladotanytarsus mancus (Walk.), Cryptochironomus gr. defectus, Stictochironomus crassiforceps (K.). Наибольшая частота встречаемости (свыше 35%) характерна для эврибионтных Ch. plumosus , P. ferrugineus , P. nubeculosum . Из олигохет доминируют Limnodrilus hoffmeisteri Clap., L. udekemianus Clap., Potamothrix hammoniensis (Mich.).
Участок ниже г. Чапаевска при комбинированном воздействии загрязнения тяжелыми металлами (ТМ) и эвтрофирования характеризуется развитием олигохет и отсутствием почти всех представителей энтомофауны, за исключением хирономид. В медиали реки массовыми видами являются олигохеты Limnodrilus hoffmeisteri Clap, L. claparedeanus Ratz. (частота встречаемости – 90%), в прибрежье – Tubifex tubifex (O.F. Mu..ller). Личинки хирономид отсутствуют в медиали реки или локально, в прибрежье в различные сезоны, встречаются Chironomus plumosus (L.), Dicrotendipes nervosus (Staeg.), Procladius ferrugineus (K.), Cricotopus sylvestris (Fabr.), Parachironomus arcuatus (Goetgh.). Показатели таксономического различия довольно существенны и отражают характерные сдвиги в биоте района, подверженного эвтрофиро-ванию и токсификации. Индекс видового разнообразия для сообществ бентоса медиали «фоновых» и «грязных»участков реки изменяется от 3,81 до 2,04. Степень обеднения макрозообентоса на участках реки с экстремально высоким уровнем загрязнения (ст. 14-16, 18; см. рис. 1) определяется стремлением донных сообществ к монодоминантности и вы- ражается низкими значениями индекса видового разнообразия (Н = 0,1-2,1). Угнетение развития бентофауны свидетельствует о токсичном характере воздействия стоков.
Установлено, что хирономиды трибы Tanytarsini отсутствуют вблизи промышлен- ных сбросов с критическим уровнем загрязнения тяжелыми металлами (например, исчезают оксифильные Cladotanytarsus mancus (Walk.), Paratanytarsus gr. lauterborni, Paratanytarsus sp.).
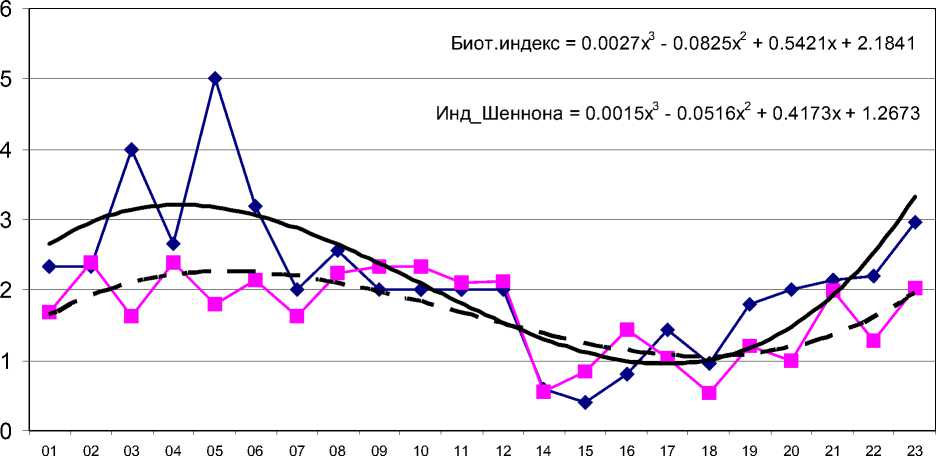
Биотический индекс
Индекс Шеннона
Рис. 1. Пространственная динамика индекса видового разнообразия и биотического индекса на русле р. Чапаевка (х - порядковый номер станции)
Выявлены специфические изменения в донных сообществах и хирономидном комплексе, которые характерны для эвтрофикации и токсификации. Для эвтрофных участков на фоне повышения общей численности и биомассы олигохет характерно доминирование эврибионтных Ch. plumosus , P. nubeculosum , D. nervosus, отсутствие оксиреофильных и стенобионтных видов , снижение числа видов и численности танитарзин. Токсифика-ция сопровождается резким сокращением числа видов и видового разнообразия бентоса и хирономид или наличием техногенных пустынь.
С учетом «биотической концепции» контроля среды [32] оценка состояния водных экосистем проводилась с приоритетом исследований биотической компоненты. При этом для нахождения адекватной связи индикаторов и индицируемых факторов в условиях антропогенной эвтрофикации и токсифика- ции была проведена интегральная оценка состояния экосистемы малых равнинных рек (на примере р. Чапаевка) с использованием комплексной системы биоиндикаторов с включением для оценки состояния водоемов изменчивость ценоза хирономид. Интегральная оценка качества экосистемы р. Чапаевка включала как водные, так и наземные индикаторы. Обобщенная схема интегральной оценки качества равнинной реки подробно обсуждалась в публикациях [23, 24] Индикационные методы показывают согласованные изменения, свидетельствующие о силе воздействия индицируемого фактора. Эффективность биоиндикации может быть повышена за счет сопряженного использования различных индикаторов и зависит от региональных условий, гидродинамических особенностей, типа рек, степени и характера антропогенной нагрузки, контрастности и физиономичнос-ти участков экосистем.
Аутэкологический спектр хирономид. Полученные в процессе многолетних исследований информационные показатели [17, 44] легли в основу создания аутэкологической характеристики ценоза хирономид в составе донных сообществ по отношению к основным абиотическим параметрам водоемов и водотоков степной и лесостепной зон Среднего и Нижнего Поволжья. Экологические шкалы составлены для хирономидофауны из обрастаний и бентоса рек. Установлено 219 видов и личиночных форм хирономид, представляющих 5 подсемейств: Tanypodinae, Diamesinae, Prodiamesinae, Orthocladiinae, Chironominae (трибы Chironomini и Tanytarsini) с преобладанием видов, распространение которых не выходит за пределы Палеарктики (табл. 2). Соотношение видов подсемейств Orthocladiinae и Chironominae составляет примерно 0,92, изменяясь от 0,4 в лимнических системах до 2,3 в холодноводных ручьях и родниках. Представители подсем. Orthocla-diinae занимают лидирующее место (68% ви- дов) в фауне бентоса и обрастаний холодноводных ручьев и родников. Они, являясь преимущественно фитодетритофагами-собирате-лями [17, 22], способны быстро колонизировать специфические микробиотопы твердых субстратов, освобожающиеся в результате периодических внешних воздействий. Представители подсем. Chironominae (57-66%) преобладают в устьевых участках равнинных рек лесостепной и степной зон, а также в заливах водохранилищ и в фауне небольших мелководных озер. Фауна подсем. Tanypodinae в общей численности видов составляет 13% (321,5%) в водотоках и водоемах разного типа (табл. 2).
Приведенные в аутэкологическом спектре данные объединяют комбинированную систему количественных и качественных составляющих генеральной совокупности полученных переменных (хирономиды – факторы среды) за многолетний периода исследований [17, 44].
Таблица 2. Количество видов и их процентное соотношение в основных таксономических группах хирономид из макрозообентоса водоемов и водотоков бассейна Средней и Нижней Волги (1987-2000 гг.)
|
Таксономическая группа |
Малые реки возвышенностей |
%% |
Средние равнинные реки |
%% |
Ручьи и родники |
%% |
Устья рек, заливы |
%% |
Озера, пруды |
%% |
Всего |
|
Tanypodinae |
16 |
10 |
23 |
15 |
1 |
3 |
12 |
13 |
12 |
21,5 |
28 |
|
Diamesinae |
9 |
6 |
6 |
4 |
- |
- |
1 |
1 |
- |
- |
9 |
|
Prodiamesinae |
3 |
2 |
2 |
1 |
- |
- |
- |
- |
- |
- |
3 |
|
Orthocladiinae |
70 |
45 |
42 |
27 |
19 |
68 |
18 |
20 |
12 |
21,5 |
86 |
|
Chironominae: Chironomini |
42 |
27 |
66 |
42 |
5 |
18 |
50 |
55 |
28 |
50 |
72 |
|
Tanytarsini |
15 |
10 |
17 |
11 |
3 |
11 |
10 |
11 |
4 |
7 |
21 |
|
Всего |
155 |
100 |
156 |
100 |
28 |
100 |
91 |
100 |
56 |
100 |
219 |
В состав доминирующего комплекса входят 13 видов хирономид (рис. 2). В многолетнем ряду наблюдений состав хирономи-дофауны равнинных рек обладает устойчивой стабильностью доминирующих форм, в то время как редкие, сопутствующие виды-«однодневки» обнаруживаются в специфических микробиотопах в определенные сезоны года. Биологическое разнообразие поверхностных вод, их специфичность, а также характерный «облик» рек определяется в первую очередь наличием или отсутствием в фауне редких таксонов [17]. Из них значительного развития достигают популяции только некоторых видов: Chironomus agilis Schobanov, Ch. piger Strenzke, Orthocladius clarki Soponis, Pseudodiamesa branickii (Now.), Paratendipes nudisquama Rdw., Tanytarsus usmaensis Pagast.
Проведенное ранжирование видов по отношению к обитанию в воде различного класса качества, дополненное картами распределения личинок хирономид (рис. 2) в реках разного типа и уровня антропогенной нагрузки, позволили выявить определенную толе- рантность хирономид к антропогенному воздействию.
К видам, условно названным «антропофилами» [20], относим те, которые встречаются (возможны единичные находки) только в грязных водах (V-VI классы качества) и (или) обитают в водах IV и V классов качества: Sergentia gr. longiventris, Parasmittia carinata Strenzke, Microtendipes gr. tarsalis, Apsectro-tanypus trifascipennis (Zett.), Labrundinia longipalpis (Goetgh.), Pentaneura sp., Campto-cladius stercorarius (de Geer), Cricotopus gr. algarum, Tanytarsus usmaensis Pagast. и др. Указанные виды можно считать устойчивыми к загрязнению и использовать (при их высокой численности) в качестве видов-ин- дикаторов антропогенной нагрузки в реках лесостепной и степной зон Средней и Нижней Волги.
Эврибионтные таксоны, примером которых служат Procladius spp., ( P. gr. ferrugineus ), Chironomus spp. ( Ch. gr. plumosus ) имеют достаточно широкий качественный диапазон возможностей обитания в воде разной степени загрязнения. Индикаторная значимость пластичных видов может быть выявлена с привлечением относительных количественных показателей, соотношения видов, ализа видовой и трофической структуры донных биоценозов вообще и хирономидных ценозов, в частности.
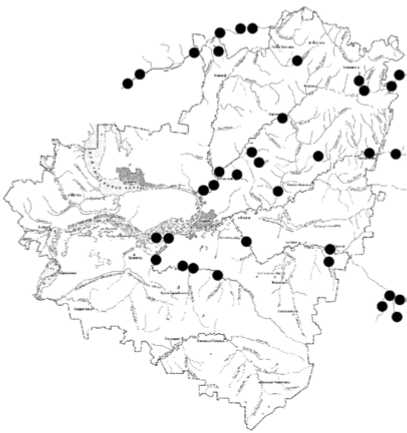
Polypedilum nubeculosum (Mg.)
Рис. 2. Пример распределения отдельных видов хирономид в реках бассейна Средней и Нижней Волги (в пределах Самарской области)
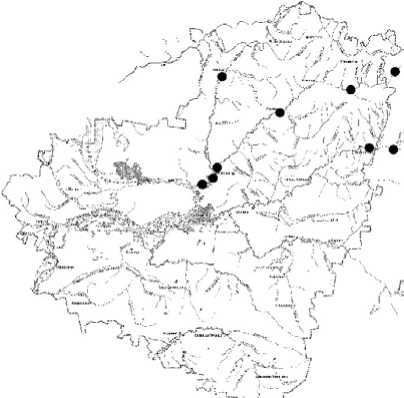
Stempellina bausei (K.)
Представленный нами портретный способ оценки индикаторных сообществ (видовая матрица) [17] выражает тенденции современного состояния хирономидофауны и может служить основой для построения фаунистических графов, математических вычислений, являться основой для конструирования разных индексов, направленных на выявление сходства, различия, разнообразия, а также оценки качества воды и классификации водоемов.
Длительные крупномасштабные биоин-дикационные исследования: экосистемный уровень. Обобщение многолетней информации по систематизации и обобщению архивных данных съемок бентоса на русле и затопленной пойме Куйбышевского водохранилища в 1958-1985 гг. и специальных экспедиционных исследований распределения донных сообществ в Приплотинном плесе в 1987-1990, 1991 и 1997 гг. позволило провести детальное исследование фауны хироно-мид за период существования водохранилища в связи с изменением его трофического статуса на фоне состояния общего бентоса. Широко известная специфическая особен-
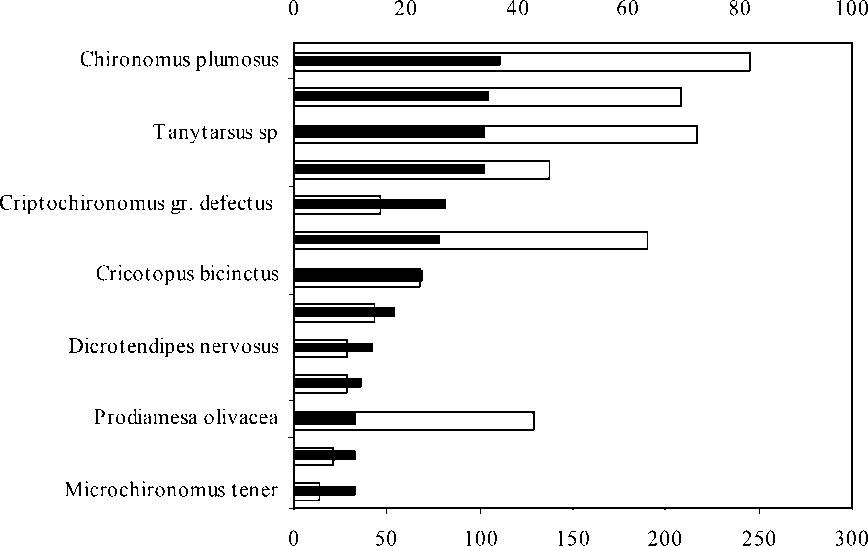
□ Средняя численность
■ Частота встречаемости
Рис. 3. Частота встречаемости и средняя численность доминирующего комплекса хирономид в реках бассейна Средней и Нижней Волги ( приводится средняя численность личинок хирономид за период исследований с 1987 по 2000 г., экз./м2 )
ность хирономид реагировать на интегральное «качество среды» позволяет использовать их для тестирования общего состояния водоемов [6, 17, 20, 28, 31, 40, 43, 49, 50].
По большинству используемых для оценки трофического статуса параметров (средняя глубина, прозрачность воды по диску Секки, величина рН, насыщенность гиполимниона кислородом, концентрация биогенных элементов, содержание хлорофилла «а», первичная продукция, содержание фитопланктона, степень развития высшей водной растительности и др.) Куйбышевское водохранилище относится к категории эвтрофных [12, 30, 37]. В 1983-1985 гг. была отмечена тенденция на увеличение биогенной нагрузки, по сравнению с периодом стабилизации в 70-е годы, что обусловлено усилением антропогенного воздействия на водоем. По величине биоген- ной нагрузки (Робщ – 7 г м-2/год, Nобщ. – 61 г м-2/год) водохранилище занимает ведущее место в каскаде волжских водохранилищ [12].
Рассматриваются три периода (1958-1966; 1967-1973 и 1975-1985 гг.), связанные с изменением трофического статуса водоема, когда наблюдаются резкие изменения уровня средневзвешенной биомассы бентоса во всех плесах водохранилища.
Начиная с 1975 г. биомасса бентоса в среднем по водохранилищу увеличилась в 4,8 раза, по сравнению с первым периодом, и составила 28,24±1,15 (n = 799), сохраняя тенденцию на увеличение, как на русле, так и на пойме, во всех плесах водохранилища. За весь период исследования соотношение биомассы основных донных организмов оставалось на уровне 54,4% – олигохет, 25,6% – личинок хирономид, 14% – моллюсков и 3,2% – ракообразных. В многолетнем ряду наблюдений соотношение основных групп бентоса, сохраняясь в целом для водохранилища, меняется в отдельных плесах. Наглядное представление о значении хирономид можно проследить при анализе динамики их видового состава и численности.
В 1958-1959 гг. при разрушении речных биоценозов хирономиды были представлены 34-39 таксонами при доминировании ре-офильных видов, характерных для фауны Волги до ее зарегулирования: Cryptochironomus rolli Kirp., С. macropodus Ljach. и Robackia demeijerei Krus. Псаммопелореофильный комплекс составляли Cryptochironomus gr. defectus , Polypedilum sp., Endochironomus sp., Dicrotendipes sp. (Волжский и Волго-Камский плесы) и другие, развивающиеся на фоне пе-лофильного сообщества, представленного уже в первые годы Chironomus sp., Procladius spр. (чаще P. gr. ferrugineus ). Начиная с 1967 г. состав хирономидофауны стабилизируется на уровне 18-25 видов, возрастая в 1975-1985 гг. до 28-31 таксона на фоне общего повышения биомассы бентоса. Для всего водохранилища по данным многолетних мониторинговых исследований установлено 53 вида хи-рономид.
Отдельные виды вышли в ранг доминан-тов исключительно после 1975 г. (высокие температуры, маловодность), который обеспечил вспышку развития всего бентоса и, в частности, хирономид: Cladopelma gr. laccophila , Cryptotendipes nigronitens (Edw.), Parachironomus vitiosus (Goetgh.), Paratendipus albimanus (Mg.), Polypedilum spр., Paralauterborniella nigrochalteralis (Mall.), Lauterborniella agrayloides (K.), Lipiniella araenicola Shilova, Cricotopus spр. и др.
Установлено, что на протяжении всех периодов и во всех районах водохранилища доминирующими по численности и биомассе были хирономиды Procladius gr. ferrugineus и Chironomus gr. plumosus (включает Сh. f. l. semireductus, Ch. f. l. reductus, Ch. plumosus). Большая плотность личинок Procladius spp. в сравнении с Chironomus spp. характерна для русловых участков и затопленной поймы всех исследованных районов. Увеличение их численности отмечается в 1975-1985 гг. Разли- чия в пищевой потребности, способах и характере питания обеспечивают преимущественное развитие личинок видов с большей пищевой пластичностью [47]. Личинки Procladius gr. ferrugineus (хищники, факультативные фитофаги), используя эпибентичес-кую пищевую нишу, «пасутся» на мейобентосе.
Трофические условия отдельных районов водохранилища, их степень эвтрофирования, климатические особенности отдельных лет, наряду с прочими факторами, являются регуляторами плотности популяций. Доля личинок Procladius sp. + Chironomus sp. в общей численности хирономид в многолетнем ряду наблюдений остается относительно неизменной. Это соотношение в бентосе русла и поймы Приплотинного плеса в 1958-1997 гг. колеблется в пределах 0,70-0,98 (табл. 3).
Для сравнительной оценки степени эвт-рофирования водохранилища или различных его участков в качестве индикаторных показателей применили соотношение численности преобладающих в бентосе олигохет и хи-рономид – NO / NC . Для Приплотинного плеса это отношение в бентосе русловой части возрастает от 23,5 в 1958-1966 гг. до 68,2 в 1975-1989 гг., а на залитой пойме в этот же период увеличивается относительная доля хирономид (отношение NO / NC снижается до 9,0).
Обращает на себя внимание резкое возрастание доли олигохет на пойме в 1991 г. (ст. 39) и отсутствие в бентосе личинок Chirono-mus sp. (табл. 3), что явилось результатом аварийного сброса неочищенных сточных вод г. Тольятти (в августе 1990 г.), повлекшим за собой резкое увеличение биогенных элементов в водной толще и, как следствие, массовое развитие синезеленых водорослей с последующим их поступлением в донные отложения [9].
На протяжении всех периодов четко прослеживается снижение численности личинок Chironomus (NCh) в отношении с хищными Procladius (NPr) как на русловых, так и на пойменных станциях. Определенное направленное изменение указанных соотношений говорит об изменении трофического статуса водоема, испытывающего влияние антропо- генного воздействия. Следствием повышения продуктивности водоема и увеличения уровня эвтрофирования является снижение среднего веса особи хирономид как на русле от 5,3 до 1,4 мг, так и на пойме – от 5,1 до 2,6
мг [18]. Процесс эвтрофирования сопровождается изменением соотношения хирономид на фоне динамики бентофауны даже в том случае, если отсутствуют адекватные данные по другим трофическим уровням.
Таблица 3. Многолетние изменения некоторых относительных показателей состояния бентоса на ст. 34 (бывшее русло Волги) и ст. 39 (затопленная пойма) Приплотинного плеса
|
Период |
NPr + NCh NС |
NCh NPr |
NO NC |
|||
|
ст. 34 |
ст. 39 |
ст. 34 |
ст. 39 |
ст. 34 |
ст. 39 |
|
|
1958-1966 гг. |
0,90 |
0,98 |
0,35 |
1,47 |
23,5 |
4,5 |
|
1967-1974 гг. |
0,91 |
0,94 |
0,40 |
0,54 |
36,5 |
16,8 |
|
1975-1989 гг. |
0,85 |
0,92 |
0,04 |
0,17 |
68,2 |
9,0 |
|
1991 и 1997 гг. |
0,70 |
0,88 |
0,02 |
0* |
6,34 |
29,4 |
Примечание: NPr + NCh NС
NCh NPr NO NC
- отношение суммарной численности хирономид ( Chironomus sp .+ Procladius sp.) к общей численности хирономид;
- отношение численности Chironomus sp . (мирные) к Procladius sp.
(хищные);
- отношение численности Oligochaeta к Chironomidae;
* - отсутствие личинок Chironomus plumosus (n = 23).
Низовья дельты р. Волги и Каспийское море. Произошедшие значительные изменения уровня воды Каспия оказали определенное влияние на состав, структуру и распределение донных биоценозов [1-3, 8, 21, 41, 42]. Наблюдаемое повышение уровня Каспийского моря привело к структурным изменениям в составе хирономидного комплекса водоемов авандельты. В 1983-1984 гг. в составе донных биоценозов авандельты наряду с типично пресноводными стали встречаться представители солоноватоводного комплекса, хирономиды Chironomus albidus Konst. [26, 27], отмеченные нами в августе 2002 г. и в Волгоградском водохранилище.
В результате подъема уровня моря в 80-х -начале 90-х годов глубины на дельтовых и авандельтовых участках возросли, что на фоне увеличения стока Волги привело к их проточности и уменьшению заиления [25, 41]. В бентофауне водоемов островной зоны авандельты произошло снижение доли пело-фильной фауны при возрастании пелорео-фильной и реофильной. Среди хирономид наряду с Chironomus plumosus (L.) , Fleuria lacustris K. , Polypedilum nubeculosum (Mg.), стали встречаться Cladotanytarsus mancus (Walk.), Polypedilum sordens (van der Wulp),
Psectrocladius sp., Clinotanypus nervosus (Mg.), Trissocladius sp., Cricotopus gr. algarum и др. Число видов хирономид увеличилось в аван-дельте от 55 (1967-1970 гг.) до 62 (1983-1984 гг.). Численность личинок осталась на уровне 1967-1970 гг., при увеличении биомассы от 3,7 до 4,76 г/м 2 [ 21].
Повышение уровня моря и его распресне-ние внесли изменения в качественный состав и количественные показатели гидробионтов. В 1991-1992 гг., при повышении водности р. Волги, подъеме уровня моря и увеличении его акватории, отмечена устойчивая тенденция на увеличение числа видов пресноводного комплекса бентоса [35]. Видовой состав хи-рономид дельты и авандельты низовьев р. Волги представлен устойчивым комплексом пелофильных и фитофильных эврибионтных видов, биомасса которых составила 4,81 г/м 2 [41, 42].
В фауне хирономид Северного Каспия (1981-1984 гг.) представители солоноватоводного комплекса личинки Chironomus albidus Konst. составляют около 90% от общей плотности хирономид. Встречаются до глубины 6 м. Биомасса личинок Ch. albidus в продуктивных районах Главного банка и Жесткого осередка на глубине 3,6 м может достигать
3,5-12,1 г/м2, при численности 700-1237 экз./м2. В 1982-1983 гг. (1,1 г/м2, 1700 экз./м2) нами установлено распространение личинок в «морской» акватории Волго-Каспийского канала. Многолетняя динамика биомассы хи-рономид в Северном Каспии на фоне общего изменения биомассы бентоса представлена в табл. 4.
Таблица 4. Изменение биомассы бентоса и хирономид (г/м 2 ) в Северном Каспии в различные периоды – 1939-1969 гг. [46]; 1981-1984 гг. (наши данные); 1990 г. [39].
|
Годы |
19391940 |
19411946 |
19471949 |
19501956 |
19571958 |
19591962 |
19631969 |
1981-1984 1990 |
|
Бентос |
9,9 |
23,3 |
26,4 |
23,7 |
68,8 |
73,3 |
49,5 |
64,2 |
|
Хирономиды |
0,3 |
0,3 |
0,4 |
0,2 |
0,5 |
0,3 |
0,2 |
1,3 |
|
Уровень моря (м абс.) |
(-27,69)(-27,91) |
(-27,95)(-27,97) |
(-27,84)(-27,89) |
(-28,07)(-28,39) |
(-28,31)(-28,17) |
(-28,13)(-28,44) |
(-28,37)(-28,40) |
(-28,15)-(-27,97); (-27,44) |
Наиболее значительное повышение биомассы бентоса наблюдалось в первые годы после зарегулирования р. Волги – в 1957-1962 гг. (табл. 4). Это было вызвано увеличением стока р. Волги, вымыванием органических веществ из ложа образованных вновь водохранилищ и их аккумуляцией в Северном Каспии. В дальнейшем, по мере снижения уровня моря (1963-1969 гг.), биомасса бентоса снизилась, что регистрировалось до 1977 г. В период повышения уровня моря (19781990 гг.) интенсивность развития бентофау-ны оказалась в 2 раза выше, чем в период понижения уровня [7]. По опубликованным данным [39], биомасса общего бентоса в Северном Каспии увеличилась до 64,2 (1990 г.) г/м 2 . Биомасса хирономид в течение маловодного периода не превышала 0,5 г/м 2 , тогда как в многоводные годы наблюдается тенденция к ее увеличению. Средняя масса хирономид в 1981-1984 гг. составила в Северном Каспии 1,3 г/м 2 (табл. 4).
По-видимому, формирующими факторами для хирономидофауны Каспия, наряду со специфическими абиотическими условиями региона, являются биотические – наличие обширных незанятых пространств и свободных экологических ниш, которые появляются как при снижении уровня моря, так и во время его повышения. Интенсивное развитие пресноводной фауны, наряду с расселением организмов солоноватоводного комплекса, приводит к быстрому повышению биомассы хи-рономид и общего бентоса. В настоящее время в Северном Каспии и в авандельте и дель- те Волги отмечается преобладание в донных сообществах морских и солоноватоводных видов [1]. Изменение гидрологических параметров в ходе колебания уровня Каспийского моря приводит к перестройке структуры сообществ зообентоса, что выражается в изменении видового состава и количественного развития хирономид, а это, в свою очередь, дает возможность использовать комплекс хирономид в качестве индикатора при мониторинге природных многолетних изменений уровня воды.
Перспективы мониторинговых исследований на основе биоиндикации природных и техногенных гидросистем. Проведенные исследования позволили предложить методику организации мониторинговых исследований, включающую несколько последовательных этапов: выделение однородных географические единиц с выявлением сходных физико-географических, гидрологических и гидрохимических факторов, а также типов антропогенного воздействия, выбор модельных водотоков; определение сети ключевых станций отбора проб и их характеристик; выбор индикаторов (организменный, популяционный, биоценотический, экосистемный уровни) и организация биоиндикационных исследований в сходные для сравниваемых водотоков периоды для оценки состояния индикаторов и динамики их изменений; проведение индикационных исследований в течение 3-5 лет в установленный период; разработка информационной системы для комплексной оценки структурных изменений в водных эко- системах; оценка экосистемы рек по комплексу гидрохимических, гидрологических и гидробиологических показателей с применением интегральных индексов их экологического состояния и построением карт распределения таксономических групп в водных экосистемах, типологическая классификация водотоков и водоемов и поиск информативных методов прогнозирования экологического состояния водных экосистем.
Список литературы Результаты и перспективы биоиндикационных исследований водоемов и водотоков Волжского бассейна (на примере хирономид, Diptera: Chironomidae)
- Аладин Н.В., Филипов А.А., Петухов В.А., Плотников И.С., Смуров А.О. Гидробиологические исследования Зоологического института РАН в дельте Волги и Северном Каспии в 1994-1997 гг. Ч. 2. Изучение зоопланктона и зообентоса//Касп. плавучий ун-т: Науч. бюл. Астрахань, 2000. № 1.
- Алексевнина М.С. К биологии и распространению Fleuria lacustris Kieff. (Diptera, Chironomidae)//Фауна и экология насекомых: Межвуз. сб. науч. тр. Пермь, 1981.
- Алимов А.Ф., Орлова М.И., Голубков С.М., Жакова Л.В., Умнова Л.П. Гидробиологические исследования Зоологического института РАН в дельте Волги и Северном Кас? пии в 1994-1997 гг. Ч. 1. Район исследований, изучение сообществ автотрофных организмов и продукционно-деструкционных процессов//Касп. плавучий ун-т: Науч. бюл. Астрахань, 2000. № 1.
- Алимов А.Ф. Сезонные и многолетние изменения биомассы зообентоса континентальных водоемов//Гидробиол. журн. 1991. Т. 27, № 4.
- Алимов А.Ф. Элементы теории функционирования экосистем. СПб.: ЗИН РАН, 2000.
- Балушкина Е.В. Функциональное значение личинок хирономид в континентальных водоемах. Л.: Наука, 1987.
- Беляева В.Н., Катунин Д.И., Осадчих В.Ф. и др. Влияние колебания уровня моря на формирование биологической и рыбной продуктивности Северного Каспия//Биологические ресурсы Каспийского моря (Тез. докл. первой междунар. конф.). Астрахань, 1992.
- Биологическая продуктивность Каспийского моря. М.: Наука, 1974.
- Варламова О.Е. Пространственное распределение, трансформация и выделение фосфора из донных отложений Куйбышевского водохранилища (на примере Приплотинного плеса): Автореф. дис.... канд. геогр. наук. М., 1998.
- Винберг Г.Г. Опыт применения разных систем биологической информации загрязнения вод в СССР//Влияние загрязняющих веществ на гидробионтов и экосистемы водоемов. Л.: Наука, 1979.
- Волга: Независимые исследования. Н. Новгород, 1994.
- Выхристюк Л.А. Биогенная нагрузка и гидрохимический режим//Экология фитопланктона Куйбышевского водохранилища. Л.: Наука, 1989.
- Выхристюк Л.А., Варламова О.Е. Химический состав воды и донных отложений//Экологическое состояние бассейна реки Чапаевка в условиях антропогенного воздействия (Биологическая индикация). Тольятти: ИЭВБ РАН, 1997.
- Выхристюк Л.А., Червякова Н.Г., Цыкало В.А. Антропогенная нагрузка на р.Чапаевка//Экологическое состояние бассейна реки Чапаевка в условиях антропогенного воздействия (Биологическая индикация). Тольятти: ИЭВБ РАН, 1997.
- Зинченко Т.Д. Хирономиды -основная группа фауны обрастаний Учинского водопроводного канала и некоторые вопросы биоиндикации качества вод//Научные основы контроля качества вод по гидробиологическим показателям: Тр. Всесоюз. конф. Л.: Гидрометеоиздат, 1981.
- Зинченко Т.Д. Хирономиды -биологические помехи водоснабжения//Перспективы развития исследований по естественным наукам на Западном Урале в свете решений XXVI съезда КПСС: Тез. докл. Пермь, 1981.
- Зинченко Т.Д. Хирономиды поверхностных вод бассейна Средней и Нижней Волги (Самарская область). Эколого-фаунистический обзор. Самара; Тольятти: ИЭВБ РАН, 2002.
- Зинченко Т.Д. Многолетнее формирование зообентоса Куйбышевского водохранилища: динамика хирономид (Diptera: Chironomidae) в связи с процессами эвтрофирования//Изв. Самар. НЦ РАН. 2003а. Спец. вып. № 2.
- Зинченко Т.Д. Особенности структурно-функциональной организации сообществ обрастаний водопроводного канала: хирономиды как биологические помехи в водоснабжении//Материалы междунар. симпоз. "Перифитон континентальных вод: современное состояние изученности и перспективы дальнейших исследований". Тюмень, 2003.
- Зинченко Т.Д. Биоиндикация как поиск информативных компонентов водных экосистем (на примере хирономид-Diptera,Chironomidae)//Чтения памяти В.Я. Леванидова. Вып. 3. Владивосток: Дальнаука, 2005.
- Зинченко Т.Д., Алексевнина М.С. Изменения хирономидофауны (Diptera, Chironomidae) в низовьях дельты Волги и в Каспийском море в связи с подъемом его уровня//Экология, эволюция и систематика хирономид: Материалы 10-го Рос. симпоз. по хирономидам. Тольятти; Борок, 1996.
- Зинченко Т.Д., Извекова Э.И., Семенов Ю.Б. Пищевое поведение личинок Cricotopus bicinctus Meig. и Orthocladius oblidens Walk. -хирономид-обрастателей водопроводного канала//Поведение водных беспозвоночных: Материалы IV Всесоюз. симпоз. Андропов, 1986.
- Зинченко Т.Д., Розенберг Г.С. (отв. ред.). Экологическое состояние бассейна реки Чапаевка в условиях антропогенного воздействия (Биологическая индикация). Тольятти: ИЭВБ РАН, 1997.
- Зинченко Т.Д., Шитиков В.К., Выхристюк Л.А., Варламова О.Е., Головатюк Л.В. Разнообразие и структурно-функциональная организация донных сообществ гидросистем бассейна Средней и Нижней Волги, их природная и антропогенная динамика//ИЭВБ РАН 20 лет: основные итоги и перспективы научных исследований. Тольятти: ИЭВБ РАН, 2003.
- Катунин Д.Н., Хрипунов И.А., Беспарточный Н.П., и др. Влияние волжского стока на гидролого-гидрохимический режим Каспийского моря//Каспийский плавучий университет: Научный бюллетень. 2000. № 1.
- Константинов А.С. К системе рода Chironomus Mg.//Тр. Сарат. отд-ния ВНИОРХ. 1956. Т. 4.
- Константинов А.С. К систематике личинок комаров рода Сhironomus//Зоол. журн. 1957. Т. 36, № 6.
- Константинов А.С. Биология хирономид и их разведение//Тр. Сарат. отд-ния ВНИОРХ. 1958. Т. 5.
- Криволуцкий Д.А., Шаланки Я., Гусев А.А. Введение. Международное сотрудничество в области биоиндикации антропогенных изменений среды//Биоиндикация и биомониторинг. М.: Наука, 1991.
- Куйбышевское водохранилище. Л.: Наука, 1983.
- Кухта А.Е. Проблемы экологического мониторинга и моделирования экосистем. Т. 16. СПб.: Гидрометеоиздат, 1996.
- Левич А.П. Биотическая концепция контроля природной среды//Докл. РАН. 1994. Т. 337, № 2.
- Макарченко Е.А. Хирономиды Дальнего Востока СССР. Подсемейства Podonominae, Diamesinae, Prodiamesinae (Diptera, Chironomidae). Владивосток, 1985.
- Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975.
- Научные основы устойчивого рыболовства и регионального распределения промысловых объектов Каспийского моря/Под ред. В.Н. Беляевой, В.П. Иванова, В.К. Зиланова. М.: ВНИРО, 1998.
- Панкратова В.Я. Личинки и куколки комаров подсемейства Orthocladiinae фауны СССР (Diptera, Chironomidae=Tendipedidae). Л.: Наука, 1970.
- Паутова В.Н., Номоконова В.И. Продуктивность фитопланктона Куйбышевского водохранилища. Тольятти, 1994.
- Руководство по методам гидробиологического анализа поверхностных вод и донных отложений/Под ред. В.А. Абакумова. Л.: Гидрометеоиздат, 1983.
- Смирнова Л.В. Динамика донной фауны в Северном Каспии//Биологические ресурсы Каспийского моря: Тез. докл. I Между-нар. Конф. Астрахань, 1992.
- Тодераш И.К. Функциональное значение хирономид в экосистеме водоемов Молдавии. Кишинев: Штиинца, 1984.
- Филъчаков В.А. Фауна, экология и распределение перакарид (бокоплавы, мизиды, кумовые и равноногие ракообразные) в водоемах дельты Волги: Автореф. дис.... канд. биол. наук. СПб., 1994.
- Чуйков Ю.С. Экологические проблемы Нижней Волги и Северного Каспия//Каспий -настоящее и будущее. Астрахань, 1995.
- Шилова А.И. Хирономиды Рыбинского водохранилища. Л.: Наука, 1976.
- Шитиков В.К., Зинченко Т.Д. Создание базы данных и алгоритмы обработки информации//Экологическое состояние бассейна реки Чапаевка в условиях антропогенного воздействия (Биологическая индикация). Тольятти: ИЭВБ РАН, 1997.
- Экологическое состояние бассейна реки Чапаевка в условиях антропогенного воздействия (Биологическая индикация)/Отв. ред. Т.Д. Зинченко и Г.С. Розенберг. Тольятти: ИЭВБ РАН, 1997.
- Яблонская Е.А., Осадчих В.Ф. Изменение кормовой базы бентосоядных рыб Северного Каспия//Тр. ВНИРО. 1973. Т. 80.
- Johnson R.K. The indicator concept in Freshwater biomonitoring//Cranston P.S. (ed): Chironomidae: from genes to ecosystems. Canberra: CSIROpublication, 1995.
- Moller Pillot H.K.M., Buskens R.F.M. De larven der Nederlandse Chironomidae (Diptera): autoekologie en verspreiding. 1C//Nederl. Faun. Meded. Nederland, 1990.
- Oliver D.R Life History ofthe Chironomidae//A. Rev. Entomol. 1971. V 16.
- Rosenberg D.M. Freshwater biomonitoring and Chironomidae//Neth. J. Aquat. Ecol. 1992. V. 26, № 2-4.
- Wiederholm T. (ed.) Chironomidae of Holarctic region: Keys and diagnoses. Part 1. Larvae//Entomol. Scand. 1983. Suppl. 19.
- Zinchenko T.D. Chironomidae (Diptera) as biological hindrances in the water-suplly//Acta Biol. Debr. Hung. 1989. V 3.
- Zinchenko T.D. Long-term (30 years) dynamics ofChironomidae (Diptera) fauna in the Kuibyshev water reservoir associated with eutrophication processes//Nether. J. of Aquat. Ecol. 1992.V. 26, № 2-4.
